
Abstract
The signals that mediate goblet cell expression of specific mucin chemotypes are poorly defined. Animal and in vitro studies show that acidomucin chemotypes may be altered by inflammation and changes in intestinal microbiota. To examine factors that may elicit this response, human adenocarcinoma-derived LS174T cells, which have a goblet cell-like phenotype and produce both sulfo- and sialomucins, were used to examine the effects of selected microbial and host factors on expression of goblet cell secretory product genes, sulfotransferases and sulfomucin production. Expression of genes encoding mucin 2 (MUC2), resistin-like molecule β (RETNLB), and trefoil factor 3 (TFF3) and Golgi sulfotransferases, carbohydrate (N-acetylglucosamine 6-O) sulfotransferase 5 (CHST5) and galactose-3-O-sulfotransferase 2 (GAL3ST2), was measured by quantitative reverse transcriptase-polymerase chain reaction following treatment with bacterial flagellin, tumor necrosis factor α (TNF-α) or the mucogenic cytokine interleukin-13 (IL-13). Expression of the toll-like receptor 5 (TLR5) gene was also analysed. Sulfomucin expression was examined via high-iron diamide/alcian blue (HID/AB) histochemistry and immunofluorescent staining for the Sulfo Lea antigen, which is synthesized in part by GAL3ST2. Flagellin, IL-13 and TNF-α all significantly increased GAL3ST2, MUC2, TFF3 and TLR5 expression, while only IL-13 increased RETNLB and CHST5 expression. Based on HID/AB histochemistry, mucin sulfation was significantly increased in response to both flagellin and IL-13 but not TNF-α. Only treatment with flagellin increased the expression of the Sulfo Lea antigen. Collectively, these results indicate that bacterial flagellin, IL-13 and TNF-α differentially modulate the expression of goblet cell secretory product genes, sulfotransferases and sulfomucin production.
Introduction
The integrity of the gastrointestinal mucosal surface is maintained by a thick mucus gel layer, which acts as a medium for protection, lubrication and transport between luminal contents and the epithelial lining. The viscoelastic and protective property of the mucus layer is attributed to mucins, which are large glycoproteins consisting of a protein core rich in serine and threonine residues that are glycosylated with O-linked oligosaccharides and cysteine-rich regions that contribute to mucus viscosity through disulfide linkages between mucin monomers. 1 Goblet cells found in columnar epithelium specialize in synthesis and secretion of secretory mucins, classified as neutral and acidic. Mucin subtypes and goblet cell distribution vary spatially throughout the gastrointestinal tract. While neutral mucins are predominant in gastric mucosa, acidomucins, such as sulfomucins and sialomucins, are expressed throughout intestinal epithelium, dominating in the large intestine. 2 In intestinal inflammatory diseases, alterations in the thickness of the mucus gel layer, goblet cell number, intracellular mucin content, and mucin glycosylation are associated with a chronic compromise in intestinal barrier function. 3–7 Although decreased sulfation of mucins typically has been associated with colorectal cancer and active inflammation in human inflammatory bowel disease (IBD), 8,9 the underlying causes and the relationship to disease pathology remain undefined. Both in vivo and in vitro studies have demonstrated an alteration in mucin chemotypes with changes in the normal microbiota and intestinal inflammation. 7,10–15 For example, in germ-free mice, crypt mucins remain predominantly sulfated, while the establishment of microbial communities parallels development of both sialo- and sulfomucin chemotypes in the colon. 11,16
A number of microbial and host factors induce mucin synthesis, secretion and degradation, but few studies have examined the role of these in determining mucin chemotypes. 11 The administration of lipopolysaccharide (LPS) from a commensal strain of Escherichia coli to germfree rats was found to increase colonic neutral mucin. 17 In addition, LPS from Helicobacter pylori inhibited mucin glycosylation and sulfation when applied to rat gastric mucosal segments. 18 Whether the effects of the microbiota and specific microbial components on mucin chemotypes occur via direct recognition by intestinal goblet cells or by a microbe-induced inflammatory response to which goblet cells respond remains to be determined. In addition, because of distinct interspecies differences in mucin chemotype patterns, 2 it is important to recognize that changes in sialo- and sulfomucin observed in animal models may not translate to humans. Likewise, among host factors, proinflammatory cytokines tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β) and IL-6 involved in the pathogenesis of Crohn's disease (CD) 3 induced the release of fewer glycosylated mucins with altered carbohydrate composition from the mucin-secreting intestinal LS180 cell line. 14 TNF-α also increased the expression of α-2,3-sialyltransferase expression in HT29 cells, 19 a finding consistent with the increased sialomucin seen in active IBD. 20,21 However, in a mouse model of salmonellosis, TNF-α was linked to an increase in sulfomucins in villus goblet cells. 10
Increased sialomucins and sulfomucins, altered terminal sugar chains, and increased expression of sulfotransferase genes were observed in rodent models of Nippostrongylus brasiliensis infection. 22–24 While the Th2 cytokines, IL-4 and IL-13, are considered to be mucogenic and important mediators of goblet cell responses to intestinal nematode infections, little is known regarding the extent to which these cytokines alter mucin composition in the human colon. 25,26 IL-4 treatment of the human goblet-cell like line, LS174T, enhanced gene expression of specific N-acetylgalactosaminyl-transferases and stimulated the attachment of N-acetylgalactosamine into a tandem repeat domain of the MUC2. 27
Mucin sulfation occurs within the trans-Golgi as a late step in glycoprotein synthesis when specific sulfotransferases transfer sulfate from 3′-phosphoadenosine 5′-phosphosulfate to mucin O-glycans at the C-3 position of galactose residues or the C-6 position of N-acetylglucosamine. 28,29 Of the known galactose 3-O-sulfotransferases and N-acetylglucosamine-6-O sulfotransferases, galactose-3-O-sulfotransferase 2 (GAL3ST2) appears to be the sulfotransferase responsible for sulfate addition to the C-3 position of Gal in human colonic mucins and carbohydrate (N-acetylglucosamine 6-O) sulfotransferase 5 (CHST5) the most likely candidate for sulfation of the C-6 position of N-acetylglucosamine. 30–32 Evidence for the involvement of these two sulfotransferases in colonic sulfomucin synthesis include the downregulation of GAL3ST2 expression in non-mucinous adenocarcinomas, corresponding to a decrease in sulfomucin expression, 30 and the abundance and specificity of CHST5 for human small intestine and colon. 31,32 In addition, CHST5 shows a preference for sulfating O-linked chains of the mucin-type, 31 suggesting that it is involved in the transfer of sulfate to the C-6 position of N-acetylglucosamine in sulfomucins.
The aim of this study was to better understand both microbial and host factors that contribute to intestinal goblet cell mucin sulfation by examining effects of bacterial flagellin, IL-13 and TNF-α on goblet cell products, sulfotransferases, CHST5 and GAL3ST2, and sulfomucin production in the colonic adenocarcinoma LS174T cell line, which exhibits a goblet cell-like phenotype. 33 This cell line synthesizes and constitutively secretes mucins as well as other goblet cell products and is responsive to conventional mucin secretagogues. 34,35 Furthermore, unlike many other mucin-producing cell lines, the LS174T cell line produces both sialo- and sulfomucin under normal growth conditions. 11,34
Materials and methods
Reagents and cell culture
The human LS174T colorectal cancer cell line was obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). LS174T cells were maintained in minimum essential medium supplemented with 10% Fetalplex (Gemini Bio-Products, West Sacramento, CA, USA), 1.5 g/L of Na2CO3, 50 units/mL penicillin G and 50 mg/mL streptomycin sulfate at 37°C in 5% CO2. Recombinant human IL-13 and TNF-α were purchased from PeproTech (Rocky Hill, NJ, USA). Recombinant flagellin from Salmonella typhimurium was purchased from InvivoGen (San Diego, CA, USA).
RNA isolation and reverse transcription
Total RNA was isolated from cells using the RNeasyPlus Kit (Qiagen, Valencia, CA, USA) according to the manufacturer's instructions. The quality and quantity of RNA isolates were determined by Nanodrop (Thermo Fisher Scientific, Pittsburgh, PA, USA). RNA isolates were reverse transcribed using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA, USA) or Superscript III First Strand Synthesis system (Invitrogen, Carlsbad, CA, USA).
Gene expression
Primers used in this study for quantitative reverse transcriptase polymerase chain reaction
*Software available from Applied Biosystems
†Available at
IL-8 secretion
Secretion of IL-8 from LS174T cells was measured in conditioned media using a Human CXCL8/IL-8 Quantikine enzyme-linked immunosorbent assay kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
Sulfomucin analysis via high-iron diamine and alcian blue histochemistry
LS174T cells were seeded in 12-well μ-chamber slides (IBIDI, Verona, WI, USA) at a density of 4 × 104 cells/well and allowed to grow for 48 h. Cells were then washed twice with 1× Hanks balanced salt solution (HBSS), and fresh antibiotic-free media containing different concentrations of flagellin, IL-13 or TNF-α were added to the cells. After 48 h, cells were washed once with 1× phosphate-buffered saline (PBS) and fixed in Carnoy's solution for 10 min. Cells were subsequently washed and stained for sulfo- or sialomucins with high-iron diamine and alcian blue (HID/AB) staining for 16 h according to a previously described protocol. 36 Stained cells were dehydrated in increasing concentrations of ethanol and cleared in xylene and were scanned at 40× using the Nanozoomer Digital Pathology System (Hamamatsu, Bridgewater, NJ, USA). Images were imported into Axiovision 4.7 software (Zeiss, Thornwood, NY, USA) and the area of sulfomucin staining was selected and measured in μm2 in each image using the Automeasure module. The total area of cells in the image was measured in μm2 and used to normalize the area of sulfomucin staining.
Analysis of Sulfo Lea antigen expression via immunofluorescence
LS174T cells were seeded in 8-well μ-slides (IBIDI) at a density of 6 × 104 cells/well and allowed to grow for 48 h. Cells were then washed twice with 1× HBSS (without phenol red), and fresh antibiotic-free media (without phenol red) containing flagellin, IL-13 or TNF-α were added to the cells. After 48 h, cells were washed 3× in 1× PBS and fixed with 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA, USA) for 20 min. Following fixation, cells were washed 3× in 1× PBS for five minutes each, incubated in 0.1% Triton X-100 for 15 min to permeabilize cells, and rinsed 3× in 1× PBS. Blocking was performed for 30 min in 5% normal goat serum (Vector Laboratories, Burlingame, CA, USA) and 5% IgG-free bovine serum albumin (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) in 1 × PBS. Cells were rinsed once in 1 × PBS, incubated for two hours at room temperature with a mouse monoclonal (F2) antibody to Sulfo Lea (Abcam, Cambridge, MA, USA) diluted 1:60 in 10% blocking solution, and washed 3× in 1 × PBS for five minutes each. Incubation with Alexa Fluor 647 goat anti-mouse IgM (H + L) (Invitrogen) diluted 1:200 in 10% blocking solution was performed for two hours at room temperature and followed by three washes with 1 × PBS for five minutes each. Cells were then counterstained with 10 μg/mL of 4′,6-diamidino-2-phenylindole (Invitrogen) for 15 min, rinsed 3× in 1 × PBS, and covered with Prolong Gold Antifade reagent (Invitrogen). After allowing Prolong Gold to cure for 24 h in the dark at room temperature, stained cells were stored in the dark at 4°C prior to imaging. Stained cells were imaged with a Zeiss LSM 710 confocal microscope using the 40× water objective and Zen 2008 software (Zeiss). Nine 2 × 2-tiled z-stacked images were collected from each well. Images were imported into Imaris (Bitplane, South Windsor, CT, USA) for reconstruction into three-dimensional (3-D) isosurfaces. Volume of Sulfo Lea staining was measured in each image in μm2 and normalized to volume of stained nuclei in each image in μm2.
Statistical analysis
Analysis of variance and Fisher's protected least significant difference test were used to compare differences. These analyses were carried out using SAS software (Statview, Version 5.0.1; SAS Institute, Cary, NC, USA). Differences were considered significant at P < 0.05.
Results
Effect of flagellin and human recombinant IL-13 and TNF-α on IL-8 secretion and toll-like receptor 5 expression
As a proinflammatory positive control, IL-8 secretion was measured in conditioned media following treatment of LS174T cells with various doses of flagellin, IL-13 and TNF-α for 24 h. As demonstrated in Figure 1a, flagellin at 500 and 1000 ng/mL significantly increased IL-8 secretion from LS174T cells. The two highest concentrations of IL-13, 10 and 100 ng/mL, however, slightly decreased IL-8 secretion (Figure 1b). Further, upon treatment of LS174T cells with TNF-α ranging from 1 to 1000 ng/mL, there was a significant increase in IL-8 secretion from LS174T cells incubated with 10, 100 and 1000 ng/mL. An approximately three-fold increase over untreated control was observed in response to 10 ng/mL TNF-α (Figure 1c). Taken together, these results indicate that LS174T cells respond to flagellin and TNF-α treatments by stimulation of proinflammatory IL-8 secretion.
IL-8 secretion from LS174T cells in response to flagellin, IL-13 and TNF-α. LS174T cells were incubated for 24 h with various doses of flagellin, IL-13 and TNF-α, and supernatants were collected to measure IL-8 secretion (expressed as ng/mL) in response to (a) flagellin (b) IL-13 and (c) TNF-α by ELISA. The data represent means ± SEM (n = 4). Statistical differences are indicated by different letters. Bars that do not show a common letter are significantly different (P < 0.05). IL, interleukin; TNF-α, tumor necrosis factor α; ELISA, enzyme linked-immunosorbent assay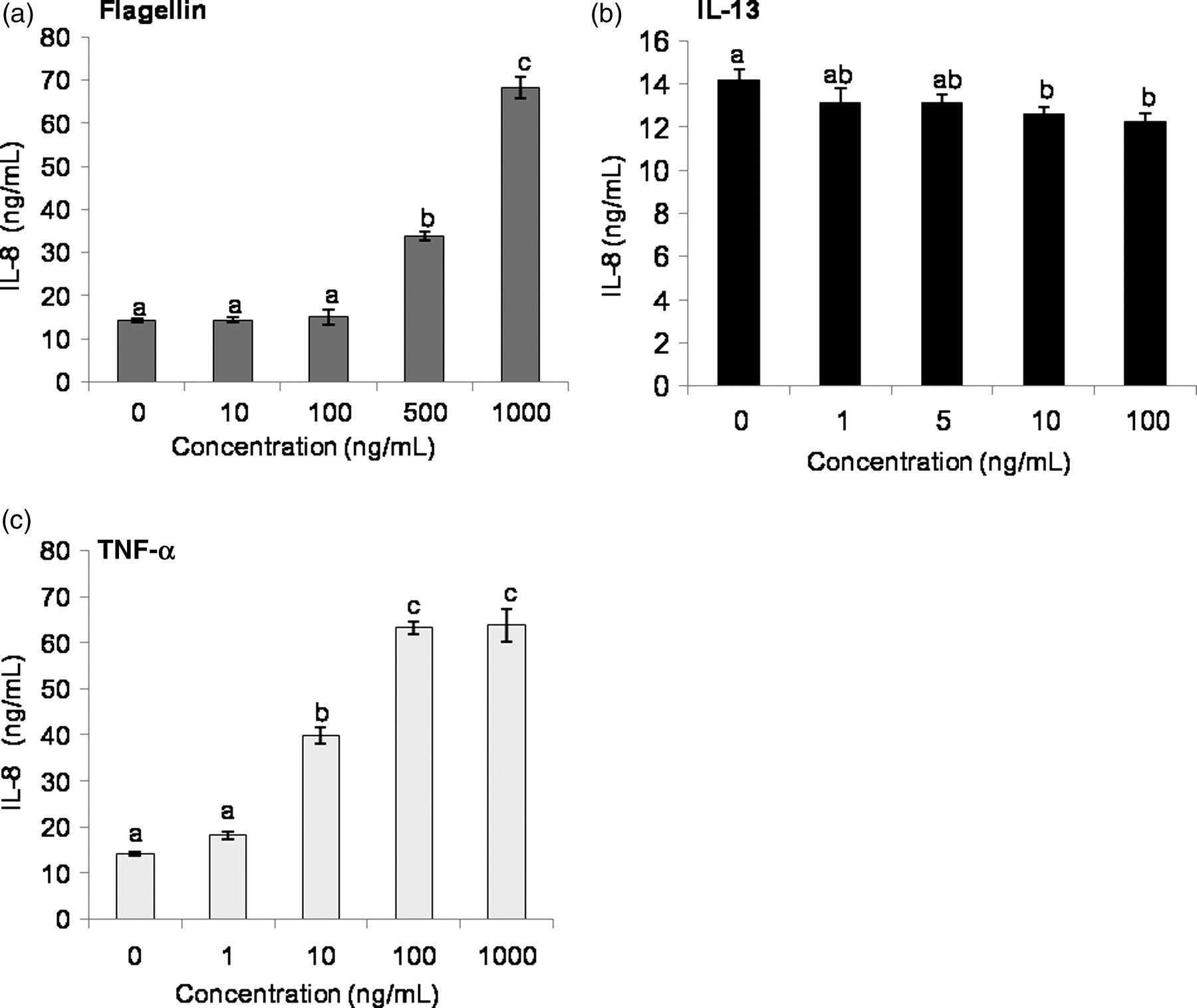
We further examined the expression of toll-like receptor 5 (TLR5) in response to its ligand flagellin and to TNF-α and IL-13. For this, LS174T cells were treated with various doses of flagellin, IL-13 and TNF-α for 24 h, and TLR5 expression was measured by realtime quantitative PCR. TLR5 expression was increased by approximately two-fold with 500 and 1000 ng/mL of flagellin (Figure 2a). Treatment with 5, 10 and 100 ng/mL of IL-13 resulted in a dose-dependent upregulation of TLR5 (Figure 2b). TNF-α also induced TLR5 expression to around 3–4-fold, again in a dose-dependent manner (Figure 2c). Collectively, flagellin, IL-13 and TNF-α induced TLR5 in a dose-dependent manner.
TLR5 expression in LS174T cells in response to flagellin, IL-13 and TNF-α. LS174T cells were treated with various doses of flagellin, IL-13 and TNF-α for 24 h and mRNA expression of TLR5 was measured by quantitative PCR. Fold change of TLR5 in response to (a) flagellin (b) IL-13 and (c) TNF-α was calculated over untreated controls. The data represent mean ± SEM (n = 4). Statistical differences are indicated by different letters. Bars that do not show a common letter are significantly different (P < 0.05). IL, interleukin; TNF-α, tumor necrosis factor α; PCR, polymerase chain reaction
Effects of flagellin and human recombinant IL-13 and TNF-α on goblet cell secretory product gene expression
With the indication that LS174T cells have high basal expression of MUC2, TFF3 and RETNLB, we examined the extent to which the expression of these genes is modulated by flagellin, IL-13 and TNF-α. For this, LS174T cells were treated for 24 h with various doses of these factors viz., 10–1000 ng/mL of flagellin, 1–100 ng/mL IL-13 and 1–1000 ng/mL of TNF-α (Figure 3). Overall, MUC2 expression increased in a dose-dependent manner with all the treatments. However, a strong induction of around 3–5-fold was observed in MUC2 expression with the highest concentration of each treatment (Figures 3a–c). The expression of RETNLB decreased significantly with flagellin and TNF-α in a dose-dependent manner; however, IL-13 at 100 ng/mL significantly upregulated the expression of RETNLB to around 2.3-fold (Figures 3a–c). TFF3 expression did not vary significantly with flagellin; however, a slight but significant increase in TFF3 expression was observed in response to IL-13 and TNF-α. Taken together, these findings indicate that MUC2 and RETNLB expression is modulated in LS174T goblet cells by various inflammatory stimuli.
Relative expression of goblet cell secretory product genes in response to flagellin, IL-13 and TNF-α. LS174T cells were treated with various doses of flagellin, IL-13 and TNF-α for 24 h, and mRNA expression of MUC2, RETNLB and TFF3 was measured by quantitative PCR. Fold change of MUC2, RETNLB and TFF3 in response to (a) flagellin (b) IL-13 and (c) TNF-α was calculated over untreated controls. The data represent mean ± SEM (n = 4). Statistical differences are indicated by different letters. Bars that do not show a common letter are significantly different (P < 0.05). IL, interleukin; TNF-α, tumor necrosis factor α; PCR, polymerase chain reaction
Effects of flagellin and human-recombinant IL-13 and TNF-α on expression of Golgi-sulfotransferase genes, sulfomucin production and Sulfo Lea antigen expression
LS174T cells were incubated with various doses of flagellin, IL-13 and TNF-α for 24 h, and RNA was isolated to analyze the expression of CHST5 and GAL3ST2 by realtime quantitative PCR. The expression of CHST5 was not modified when compared with untreated control, during incubation of LS174T cells with flagellin (Figure 4a). However, IL-13 increased CHST5 expression in a dose-dependent manner up to 7.5-fold for the highest concentration (Figure 4b). A modest increase of CHST5 expression to around 1.4-fold was observed with 10 ng/mL of TNF-α. The expression of GAL3ST2 was significantly increased approximately 2.5-fold with the two highest concentrations of flagellin (Figure 4a), while a three-fold increase was observed upon treatment with 10–1000 ng/mL of TNF-α. Also, there was significant increase in GAL3ST2 expression of approximately 2–3-fold in response to100 ng/mL of IL-13 (Figures 4b and c). Hence, both IL-13 and TNF-α as well as flagellin showed substantial induction of Golgi-sulfotransferase genes in LS174T cells.
Relative expression of Golgi sulfotransferase genes in LS174T cells. mRNA expression of CHST5 and GAL3ST2 was measured in LS174T cells by quantitative PCR in response to treatment with various doses of (a) flagellin (b) IL-13 or (c) TNF-α for 24 h. Fold induction was calculated over untreated controls. mRNA expression of (d) CHST5 and (e) GAL3ST2 was measured in LS174T cells in response to IL-13 (100 ng/mL) for various time periods, and fold induction was calculated over untreated controls taken at each time point. The data represent mean ± SEM (n = 3). Statistical differences are indicated by different letters. Bars that do not show a common letter are significantly different (P < 0.05). IL, interleukin; TNF-α, tumor necrosis factor α; PCR, polymerase chain reaction
As the expression of CHST5 and GAL3ST2 was modulated by treatment with various factors at 24 h, we further examined how the time period of incubation affects their expression. For time kinetics, LS174T cells were treated with IL-13 at a fixed dose of 100 ng/mL varying from 12 to 96 h. As demonstrated in Figure 4d, CHST5 expression was induced to approximately 6–7-fold as early as 12 h post-treatment and remained high at 48 h and thereafter, its expression decreased. On the other hand, GAL3ST2 expression peaked at 24 h to around approximately 2–3-fold and thereafter it decreased (Figure 4e). These data confirm that expression of Golgi-sulfotransferase genes in LS174T goblet cells is modulated in both a dose- and time-dependent manner in response to the cytokines IL-13 and TNF-α, and bacterial flagellin.
LS174T cells synthesize both sialo- and sulfomucin under normal growth conditions.
11,34
Taking this and the observations in Figure 4 into consideration, we examined the effects of flagellin, IL-13 and TNF-α on LS174T acidomucin chemotypes. LS174T cells were again treated with various doses of flagellin, IL-13 and TNF-α for 48 h, and the synthesis of sialo- and sulfomucin chemotypes was measured by HID/AB histochemistry. Comparison between treated and untreated control cells revealed a significant increase in sulfomucin expression in cells treated with 5 and 100 ng/mL of IL-13, and 1000 ng/mL of flagellin (Figures 5a and b). A numerical increase was observed in the abundance of sulfomucinsin response to TNF-α; however, these differences were not statistically significant with 48 h of treatment (Figure 5c).
Sulfomucin expression from LS174T cells. LS174T cells were treated with various doses of flagellin, IL-13 and TNF-α for 48 h. Cells were fixed and stained for sulfomucins and sialomucins. (a) High-iron diamide/alcian blue blue staining for sulfo- and sialomucuins under indicated treatments. (b) Quantification of sulfomucins produced from LS174T cells after treatment with flagellin, IL-13 or TNF-α. Arrows indicate sulfomucin-positive areas. The data represent means ± SEM (n = 3–4). Statistical differences are indicated by different letters. Bars that do not show a common letter are significantly different (P < 0.05). (A color version of this figure is available in the online journal)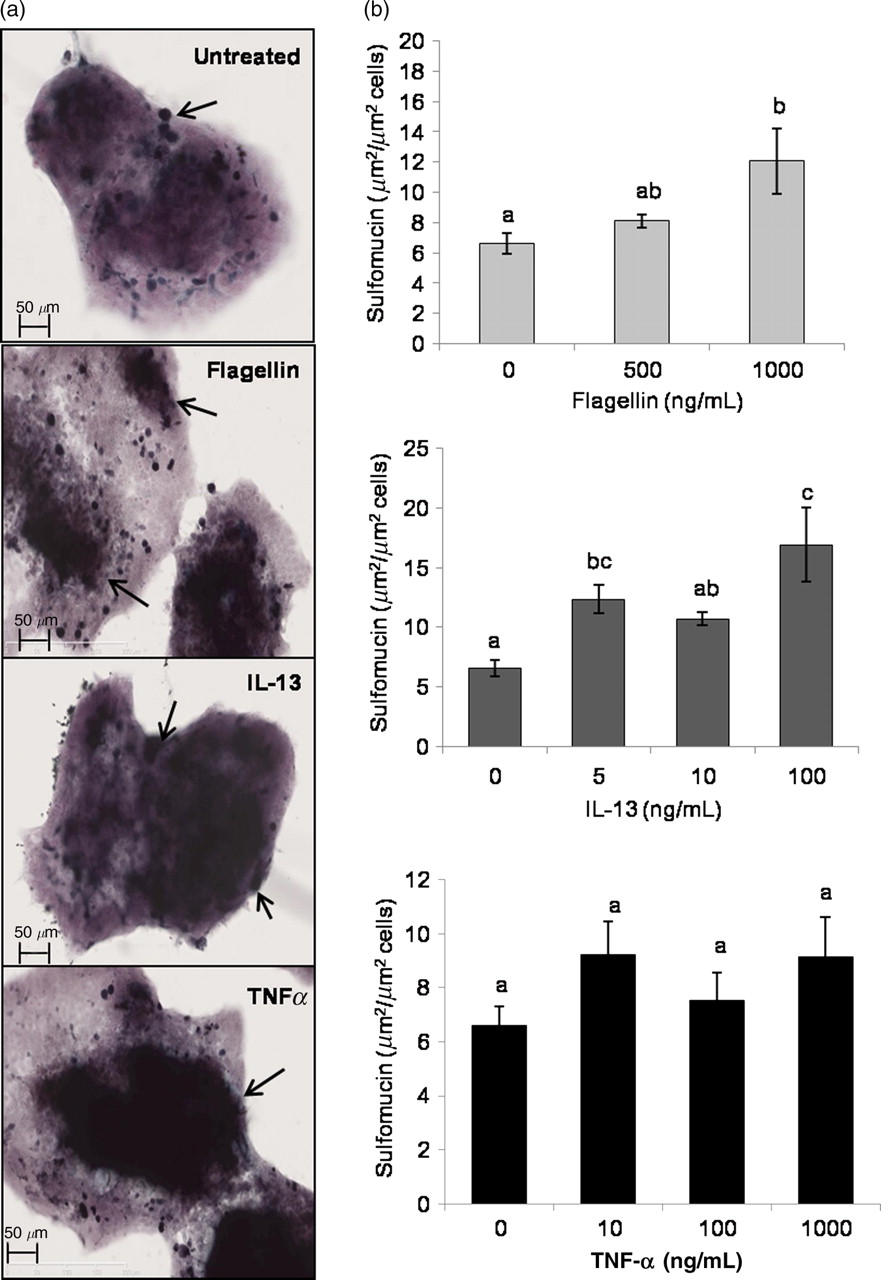
Sulfo Lea antigen is expressed on the major high-molecular-weight sulfated-glycoprotein produced by LS174T cells.
37
Thus, we examined the effects of flagellin, IL-13 and TNF-α on Sulfo Lea expression in these cells as these factors modulated both sulfotransferase expression and sulfomucin production in LS174T cells (Figures 4 and 5). LS174T cells were treated with 500 ng/mL of flagellin, 100 ng/mL of IL-13 or 100 ng/mL of TNF-α for 48h. These concentrations were selected based on the lowest concentrations of flagellin, IL-13 and TNF-α that produced the maximal increase in GAL3ST2 expression at 24 h (Figure 4). A representative z-stacked, tiled image of the immunofluorescent staining is shown in Figure 6a and a 3-D isosurface generated from that data is shown in Figure 6b. Comparison of the volume of Sulfo Lea staining per volume of nuclear staining between treatments revealed a significant increase in Sulfo Lea expression in LS174T cells treated with 500 ng/mL of flagellin for 48 h, while treatment with IL-13 or TNF-α did not significantly alter the level of Sulfo Lea expression (Figure 6c) during this time period.
Sulfo Lea antigen expression in LS174T cells. (a) Immunofluorescent staining of the Sulfo Lea antigen (red) in LS174T cells and nuclear staining with DAPI (green). (b) 3-D isosurface generated from z-stacked image. (c) Sulfo Lea antigen expression in LS174T cells in response to incubation for 48 h with the indicated treatments. The data represent means ± SEM (n = 3–4). Statistical differences are indicated by different letters. Bars that do not show a common letter are significantly different (P < 0.05). DAPI, 4′,6-diamidino-2-phenylindole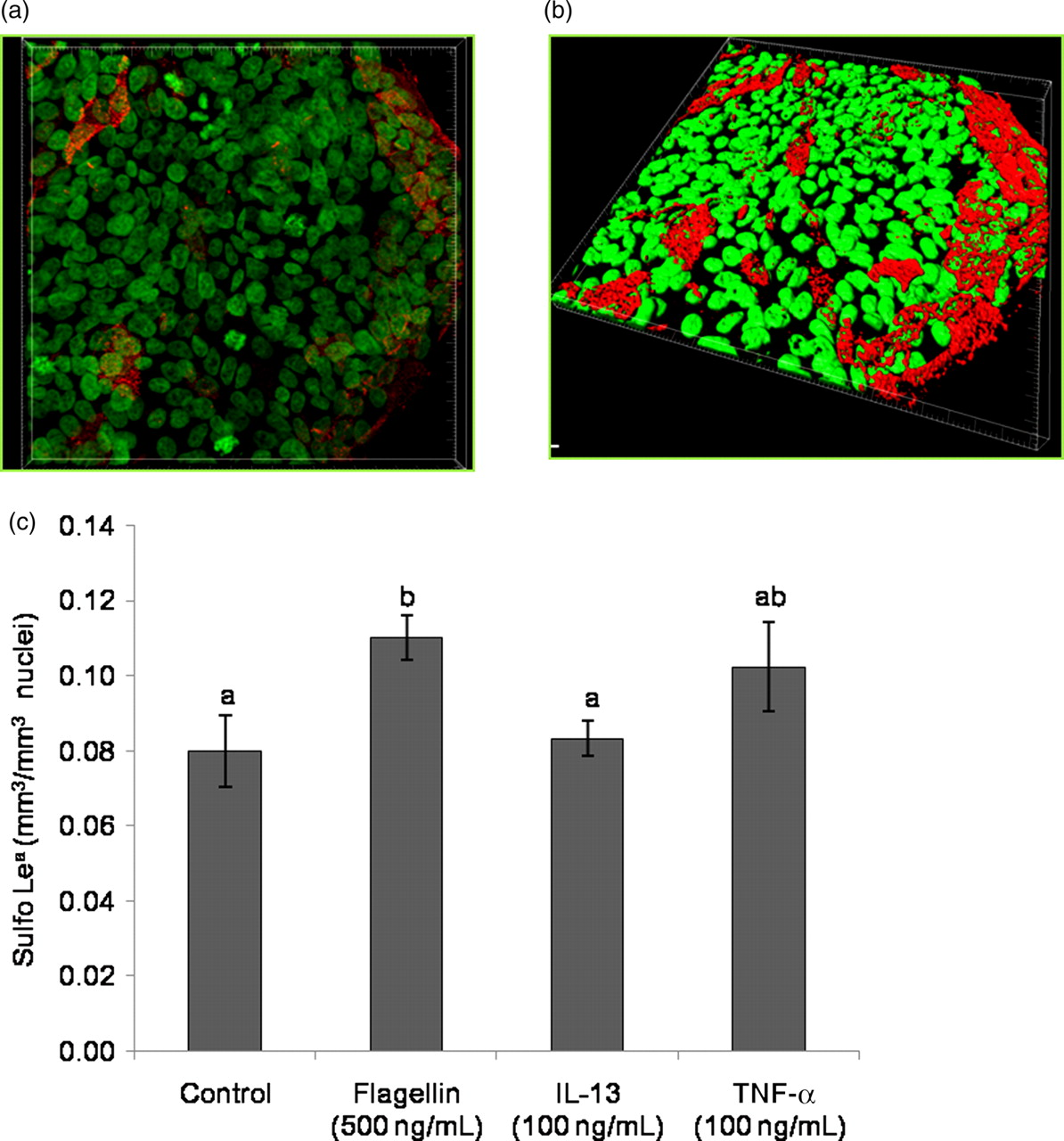
Discussion
Few studies have examined the extent to which individual microbial and host factors modify expression of specific mucin chemotypes and particularly mucin sulfation. In the present study, we focused on the role of bacterial flagellin and inflammatory cytokines IL-13 and TNF-α on the expression of goblet cell secretory product genes, mucin sulfotransferases and sulfomucin production. We found that expression of MUC2 was significantly induced by flagellin, IL-13 and TNF-α in a dose-dependent manner. The expression of RETNLB decreased with increasing doses of IL-13 and TNF-α. Further, we observed high induction of CHST5 as early as 12 h after treatment with IL-13. Flagellin, IL-13 and TNF-α all, on the other hand, induced GAL3ST2 expression. The observed induction of sulfotransferases was consistent with the increased sulfomucin synthesis from LS174T cells observed in response to IL-13 and flagellin, indicating that sulfotransferases and sulfomucin synthesis can be differentially modulated by particular inflammatory signals.
LS174T cells expressed all the human TLRs; however, because TLR5 was among the most abundantly expressed TLR, flagellin, a known agonist of TLR5, 38 was selected as a microbial factor for further examination. Besides induction of TLR5 by flagellin, a dose-dependent increase in TLR5 expression was observed with IL-13 and TNF-α. While the response of IL-13 and TNF-α on TLR5 has not been studied previously, flagellin was shown to activate basolaterally expressed TLR5 to induce epithelial-driven proinflammatory gene expression. 39 Chronic inflammatory conditions can lead to aberrant IL-8 production, which has proinflammatory activity. 40 The enhanced IL-8 secretion observed in our study following treatment with flagellin and TNF-α is in accordance with results from previous studies in which TNF-α induced IL-8 secretion from LS174T and HM7 cells, 38 while flagellin induced IL-8 secretion from unstimulated epithelia when conditioned media from S. typhimurium-infected T84 and HT29-cl.19A cell-derived model intestinal epithelia was used. 41 Further, the decreased IL-8 secretion observed with IL-13 is consistent with a previous report where Th2 cytokines IL-4 and IL-13 significantly reduced IL-8 secretion by human intestinal epithelial cells. 42
MUC2 is the major gel-forming mucin in the small and large intestine, and its expression can be modulated by several factors including toxins, microbial products, hormones, cytokines, bile salts and other growth factors. 43 We found induction of MUC2 as a common response to flagellin, IL-13 and TNF-α. Similar observations have been reported for human colon adenocarcinoma cells (HT29, HCT8 and HM3M2 cell lines), where TNF-α upregulated MUC2 expression via the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway and IL-4 and IL-13 via the mitogen-activated protein (MAP) kinase pathway. 44,45 Flagellin from Pseudomonas aeruginosa also activated MUC2 transcription following binding to the surface glycolipid receptor, Asialo-GM1; 1,46 however, MUC2 induction in response to flagellin from S. typhimurium has, to our knowledge, not been reported previously.
RETNLB and TFF3 represent other goblet cell secretory products with distinct functions. TFF3 contributes to epithelial restitution on mucosal surfaces, and RETNLB is highly induced with enteric bacterial colonization and helminth infection. 26,43,47 In this study, TFF3 was slightly but significantly increased in response to IL-13 and TNF-α. The enhanced expression of TFF3 in response to IL-13 has been reported for HT29 CL.16E cells, which develop a goblet-cell-like phenotype after reaching confluence. 25 The increased expression of RETNLB observed with IL-13 is in accordance with the earlier studies demonstrating upregulation of RETNLB in LS174T cells treated with IL-13; 26,47,48 however, the dose-dependent decrease in the expression of RETNLB observed with flagellin or TNF-α has not been reported previously. Further, disruption of RETNLB has been associated with reduced severity of colitis in a murine model of dextran sodium sulfate-induced colonic injury 49 and, more recently, has been shown to protect against N. brasiliensis infection in mice by inducing intestinal epithelial cells to differentiate into goblet cells that produce RELM-β. 50 RETNLB, therefore, plays an important role in maintaining homeostatic gastrointestinal function and as a mediator of mucosal inflammatory responses. 49,51 The upregulation of both MUC2 and TFF3 holds relevance, as interactions between these goblet cell products serve to stabilize the mucus gel layer, especially under inflammatory conditions. 52,53
Sulfomucins have been implicated in protection of the intestinal mucosa. 54,55 While CHST5 shows a preference for sulfation of mucin O-glycans, GAL3ST2 has been linked with sulfomucin synthesis, and its expression was found to be downregulated in non-mucinous adenocarcinoma. 30–32 GAL3ST2 expression was enhanced in LS174T cells following treatment with flagellin, IL-13 and TNF-α for 24 h, indicating that increased mucin sulfation at the C-3 position of Gal might be a common response to inflammatory stimuli. The induction of all four GAL3STs has been reported previously in a N. brasiliensis infection model, which exhibits a Th2 phenotype. 24 Analysis of CHST5 expression revealed IL-13 to be a strong inducer of its expression, thereby indicating that mucin sulfation at the C-6 position of N-acetylglucosamine may be augmented in response to IL-13 as early as 12 h after treatment. Further, increased expression of these sulfotransferases corresponded to an increase in sulfomucin production by LS174T cells in response to flagellin and IL-13. Only exposure to flagellin increased the expression of the Sulfo Lea antigen, which is synthesized in part by GAL3ST2. 56 Taken together, these results indicate that IL-13 may increase sulfomucins primarily via increased expression of CHST5, while flagellin may increase sulfomucins, including those with the Sulfo Lea antigen, by enhancing the expression of GAL3ST2.
Considering that the protective role of sulfomucins may be due to their ability to increase mucus viscosity and resistance to both bacterial degradation and microbe adhesion, 57 the results with TNF-α are of particular interest because of its association with IBD and the alterations in mucin sulfation seen with active IBD. 7,8 As IBD is characterized by dysregulated immune responsiveness, 58 it may be that individuals with IBD fail to respond appropriately to TNF-α and, hence, fail to increase the production of sulfomucins. However, additional studies will be needed to better understand the mechanisms by which flagellin, IL-13, and TNF-α enhance expression of specific sulfotransferases and sulfomucin production. It is possible that pathways governing goblet cell secretory product gene expression in response to TNF-α and IL-13 44,45,59,60 may be similarly involved in the regulation of CHST5 and GAL3ST2. Further, CDX2 has been shown to transactivate the promoters of MUC2, RETNLB and TFF3, 61–63 and we found induction of CDX2 in response to IL-13 (data not shown). Therefore, it is possible that CDX2 may also transactivate CHST5 or even GAL3ST2, which, however, remains to be explored. Collectively, our results indicate that flagellin, IL-13 and TNF-α exhibit differential effects on the expression of goblet cell secretory product genes, mucin sulfotransferases and sulfomucin production, providing further evidence that both microbial and host factors influence mucin synthesis and sulfation.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by NIH grant DK061568 (HRG). The authors thank Ann Benefiel for editorial assistance.