
Abstract
Epinephrine controls many important and sometimes opposite processes. This pleiotropic effect is achieved via coupling to different receptor/effector systems. In epididymal white adipose tissue (EWAT) of Wistar rats, we showed that epinephrine stimulated protein kinase B (PKB) phosphorylation on Ser473. Epinephrine further increased the glucose incorporation into glyceride-glycerol without decreasing glucose availability for other metabolic pathways (i.e. lactate production). Wortmannin (phosphatidylinositol 3-kinase inhibitor) treatment significantly decreased glucose incorporation into glyceride-glycerol and elevated the epinephrine-induced release of free fatty acids (FFA) from the adipose tissue without any change in the intensity of lipolysis measured as glycerol release. Using specific cyclic adenosine monophosphate (cAMP) analogs we demonstrated that cAMP–protein kinase A (PKA) signalling resulted in a strong PKB dephosphorylation and significantly lowered the glucose availability in EWAT. Specific activation of the Epac (exchange protein activated by cAMP)-dependent pathway had only a moderately negative effect on PKB phosphorylation and glucose metabolism. In contrast, α 1 agonist methoxamine increased PKB phosphorylation and lactate production. This effect of methoxamine was additive to the effect of insulin and it was abolished by wortmannin treatment. In EWAT of spontaneously dyslipidemic hereditary hypertriglyceridemic (HHTg) rats, we demonstrated significantly lower epinephrine-induced glucose utilization but higher sensitivity to its lipolytic effect. We conclude that in EWAT, epinephrine controls two opposite processes (FFA release and FFA retention) via two different effector systems. The impairment of α 1-dependent, epinephrine-stimulated, glycolysis-dependent FFA esterification may contribute to the establishment of dyslipidemia in insulin resistance.
Introduction
Epinephrine controls a wide range of important processes and its effects strongly depend on the type of receptor/effector pathway. The stimulation of β-adrenoreceptors leads to the increased formation of cyclic adenosine monophosphate (cAMP) via adenylate cyclase. For many years, major intracellular effects of cAMP were believed to be mediated by cAMP-dependent protein kinase A (PKA). Once activated, PKA can affect a range of diverse cellular events by phosphorylating an array of cytoplasmic and nuclear protein substrates, including enzymes and transcription factors. In white adipose tissue (WAT), the major role of PKA is the activation of lipolysis by phosphorylating hormone-sensitive lipase and perilipin. 1 PKA further inhibits insulin signalling by blocking phosphatidylinositol-3 (PI-3) kinase activity and consequently the activities of protein kinase B (PKB) and other downstream targets of the PI-3K pathway. 2,3 Recently, a novel intracellular target for cAMP (exchange protein activated by cAMP; Epac) was discovered. 4,5 Growing evidence indicates that PKA and Epac acts through a distinct set of signalling pathways and can exert opposing effects in regulating downstream targets such as PKB.
Glucose metabolism is a subject of stringent hormonal regulation and two major regulators, insulin and epinephrine, have been considered to exert opposing effects. Nevertheless, it has been shown that epinephrine may also positively stimulate insulin signalling cascade and glucose utilization in various tissues and cell types. The positive effect of epinephrine on glucose metabolism in adipose tissue was first reported 30 years ago. 6 Faintrenie and Geloen 7,8 and Cheng et al. 9 brought evidence indicating that glucose uptake and lactate production in rat white adipocytes is under α 1-adrenergic instead of β-adrenergic (i.e. cAMP-dependent) control. It has been established that activation of α 1-adrenoreceptors may stimulate the hydrolysis of phosphatidylinositol (4,5)-biphosphate by phospholipase C to produce inositol-1,4,5-phosphate. 10,11 This molecule is an important second messenger and serves as the substrate for PI-3 kinase, which has been implicated as a key player in the insulin signalling cascade. 12 However, the role of PI-3 kinase in the signal pathway of epinephrine is still unclear.
In the present study, we investigated the contribution of β-adrenoreceptor (i.e. cAMP-dependent) and α 1-adrenoreceptor-mediated pathways to the PKB phosphorylation and glucose utilization in epididymal WAT (EWAT). We report that the activation of both Epac- and PKA-dependent pathways downregulated PKB phosphorylation, the effect of the latter being much stronger. The decrease of PKB phosphorylation was accompanied by lower lactate production and glucose incorporation into glyceride-glycerol. In contrast, stimulation of α 1-adrenoreceptors by methoxamine was associated with the increased PKB phosphorylation and glucose utilization in the tissue. This process was wortmannin-sensitive, which indicated the involvement of PI-3 kinase.
Materials and methods
Animals and experimental protocol
Male rats (350 ± 15 g) were kept in a temperature-controlled room at 12:12 h light–dark cycle. Animals had free access to drinking water and standard laboratory chow diet. All experiments were performed in agreement with the Animal Protection Law of the Czech Republic 311/1997, which is in compliance with Principles of Laboratory Animal Care (NIH Guide to the Care and Use of Laboratory Animals, 8th edn, 2011 13 ) and were approved by the ethical committee of the Institute for Clinical and Experimental Medicine. Wistar rats were obtained from AnLab CR (Prague, Czech Republic). The strain of hereditary hypertriglyceridemic (HHTg) rats was originally selected from Wistar strain rats in our laboratory according to their susceptibility to short-term high sucrose feeding. 14 Parent pairs were assigned to low-triglyceridemic and high-triglyceridemic groups on the basis of triglyceridemia measurements. Their offspring were bred by cousin × cousin breeding up to 10 generations and brother × sister thereafter. This non-obese strain exhibits, apart from hypertriglyceridemia, a high serum free fatty acid (FFA) content, hyperinsulinemia and impaired glucose tolerance. 15,16
Chemicals
Adipose tissue incubation procedure
The animals were killed by decapitation. Approximately 150 ± 25 mg of EWAT was rapidly dissected. Only thin distal parts (relative to the testes) were used, which ensured sufficient oxygen and nutrient delivery to the tissue and product removal to the medium during the incubation in vitro. The tissues were incubated two hours in Krebs–Ringer buffer with 5 mmol/L glucose,
Measurement of lipogenesis and re-esterification in adipose tissue in vitro
The estimation of the 14C-glucose incorporation into total lipids was carried out as described before. 17 Briefly, adipose tissue was removed from the incubation medium, rinsed in physiological solution and immediately put into CH3Cl. The pieces of tissue were dissolved using a teflon pestle homogenizer, methanol was added (CH3Cl:methanol 2:1) and lipids were extracted at 4°C overnight according to Folch et al. 18 The next day the residual tissue was removed and the clear extract was taken for further analysis. An aliquot was evaporated, reconstituted in scintillation liquid and the radioactivity was measured by scintillation counting.
To determine the site (glycerol versus acyl moiety) of glucose incorporated into neutral lipids, an aliquot of clear extract was evaporated and saponified in ethanolic 15% potassium hydroxide at 70°C. Saponification was terminated by adding 5.4 mol/L H2SO4. After cooling, the released fatty acids were extracted repeatedly into petroleum ether. The pooled petroleum ether fractions were evaporated, reconstituted in scintillation liquid and the radioactivity was measured by scintillation counting. The amount of radioactivity incorporated into glycerol residues was calculated as the difference of total activity incorporated into neutral lipids and petroleum ether fraction of the same aliquot.
PKB phosphorylation
Distal parts of adipose tissue (150 ± 25 mg) were incubated and pharmacologically treated for 120 min as described in the adipose tissue incubation procedure. Three samples from one animal were pooled, frozen in liquid nitrogen and stored in −80°C until further analysis. The homogenate was prepared with a micro Ultra-Turax homogenizer (IKA Werke, Staufen, Germany) in homogenization buffer (150 mmol/L NaCl, 2 mmol/L ethylenediaminetetraacetic acid [EDTA], 50 mmol/L TRIS, 20 mmol/L glycerolphosphate, 1 mmol/L Na3VO4, 2 mmol/L sodium pyrophosphate, 1 mmol/L phenylmethanesulfonyl fluoride [PMSF], leupeptin 10 μg/mL, aprotinin 10 μg/mL). Immunodetection was performed as described elsewhere. 19 The proteins were resolved by electrophoretic separation under denaturating conditions and electroblotted onto polyvinylidene fluoride membranes. The level of phosphorylation of PKB was assessed by immunodetection using specific phospho-PKB (Ser473) antibody. The total expression of PKB protein was determined on the same membrane after stripping and re-blotting using anti-PKB antibody. The antibodies were purchased from Cell Signalling Technology (Boston, MA, USA). The loading control was performed using rabbit polyclonal antibody to beta-actin (Abcam, Cambridge, UK). The bands were visualized using ECL and quantified using FUJI LAS-3000 imager (Fujifilm, Tokyo, Japan) and Quantity One software (Bio-Rad, Hercules, CA, USA).
Statistical analysis
Data are presented as mean ± SEM. Statistical analysis was performed using the Kruskal–Wallis test with multiple comparisons (n = 5–7). Differences were considered statistically significant at the level of P < 0.05.
Results
Epinephrine stimulates glucose utilization in EWAT and this effect is abolished by wortmannin
Effect of wortmannin on epinephrine-stimulated lipolysis and glucose utilization in EWAT
Distal parts of epididymal adipose tissue (relative to the testes) were incubated for 120 min in the presence of 14C-labelled glucose. When indicated, samples were preincubated for 15 min in the presence of wortmannin and then exposed to the epinephrine for another 120 min. FFA, glycerol and lactate content in the incubation medium was determined at the end of the incubation period. Glucose incorporation into glyceride-glycerol was determined after the extraction of neutral lipids, hydrolysis and separation of organic and water-soluble fractions. Data are expressed as mean ± SEM, n = 7–10
EWAT, epididymal white adipose tissue; FFA, free fatty acids
***P < 0.001 versus basal; # P < 0.05; ### P < 0.001 wortmannin + epinephrine versus epinephrine-treated
β-Adrenergic receptor-dependent signalling has a negative effect on glucose utilization in EWAT
Effect of β-adrenergic signalling on lipolysis and glucose utilization in EWAT
Experimental design is described in Table 1. Epac activator: 8-pCPT-2′-O-Me-cAMP; PKA activator: N 6-Benzoyl-cAMP. Data are expressed as mean ± SEM, n = 7–10
*P < 0.05, **P < 0.01, ***P < 0.001 versus basal
α 1-Adrenergic receptor agonist has a positive effect on glucose utilization in EWAT
Effect of α 1-adrenergic receptor agonist methoxamine on lipolysis and glucose utilization in EWAT
Experimental design is described in Table 1. When indicated, samples were preincubated for 15 min in the presence of wortmannin and then exposed to the methoxamine or insulin for another 120 min. Data are expressed as mean ± SEM, n = 7–10
*P < 0.05, **P < 0.01, ***P < 0.001 versus basal; †† P < 0.01, ††† P < 0.01 wortmannin-treated versus untreated
Effect of α 1-adrenergic receptor antagonist prazosin on adrenalin-stimulated lipolysis and glucose utilization in EWAT
Experimental design is described in Table 1. When indicated, samples were preincubated for 15 min in the presence of prazosin and then exposed to the epinephrine for another 120 min. Data are expressed as mean ± SEM, n = 7–10.
*P < 0.05, ***P < 0.001 versus basal; †† P < 0.01 prazosin + epinephrine versus epinephrine-treated group
Epinephrine stimulates PKB phosphorylation in EWAT
PKB is an important downstream target of PI-3 kinase and its activation is associated with phosphorylation at Ser374. Figure 1 shows the immunoblot of Ser374-PKB phosphorylation in EWAT incubated in vitro with insulin (100 μU/mL), epinephrine 1 μmol/L, methoxamine 10 μmol/L, isoproterenol 1 μmol/L, 8-pCPT-2′-O-Me-cAMP (Epac activator; 0.5 mmol/L) or N
6-Benzoyl-cAMP (PKA activator; 1.66 mmol/L). Maximal stimulatory effect was reached after insulin stimulation that served as a positive control. Significant phosphorylation above basal state was further documented after epinephrine and methoxamine treatment. In contrast, Epac activator moderately decreased PKB phosphorylation and in the presence of isoproterenol or specific PKA activator, we found virtually no Ser374 phosphorylated PKB.
PKB phosphorylation. Distal parts of epidydimal adipose tissue (relative to the testes) were incubated in Krebs–Ringer bicarbonate buffer either alone or pharmacologically treated (insulin 100 μU/mL; epinephrine 1 μmol/L; methoxamine 10 μmol/L; isoproterenol 1 μmol/L; Epac activator 0.5 mmol/L; PKA activator 1.66 mmol/L). The results are expressed as fold change in PKB (Ser473) phosphorylation compared with basal condition. Representative Western blots are shown above the graph. The total PKB protein expression was determined after stripping the membrane and re-blotting with anti-PKB antibody. Values represent means ± SEM of seven animals. *P < 0.05; ***P < 0.001 versus basal. Epac, exchange protein activated by cAMP; PKA, protein kinase A; PKB, protein kinase B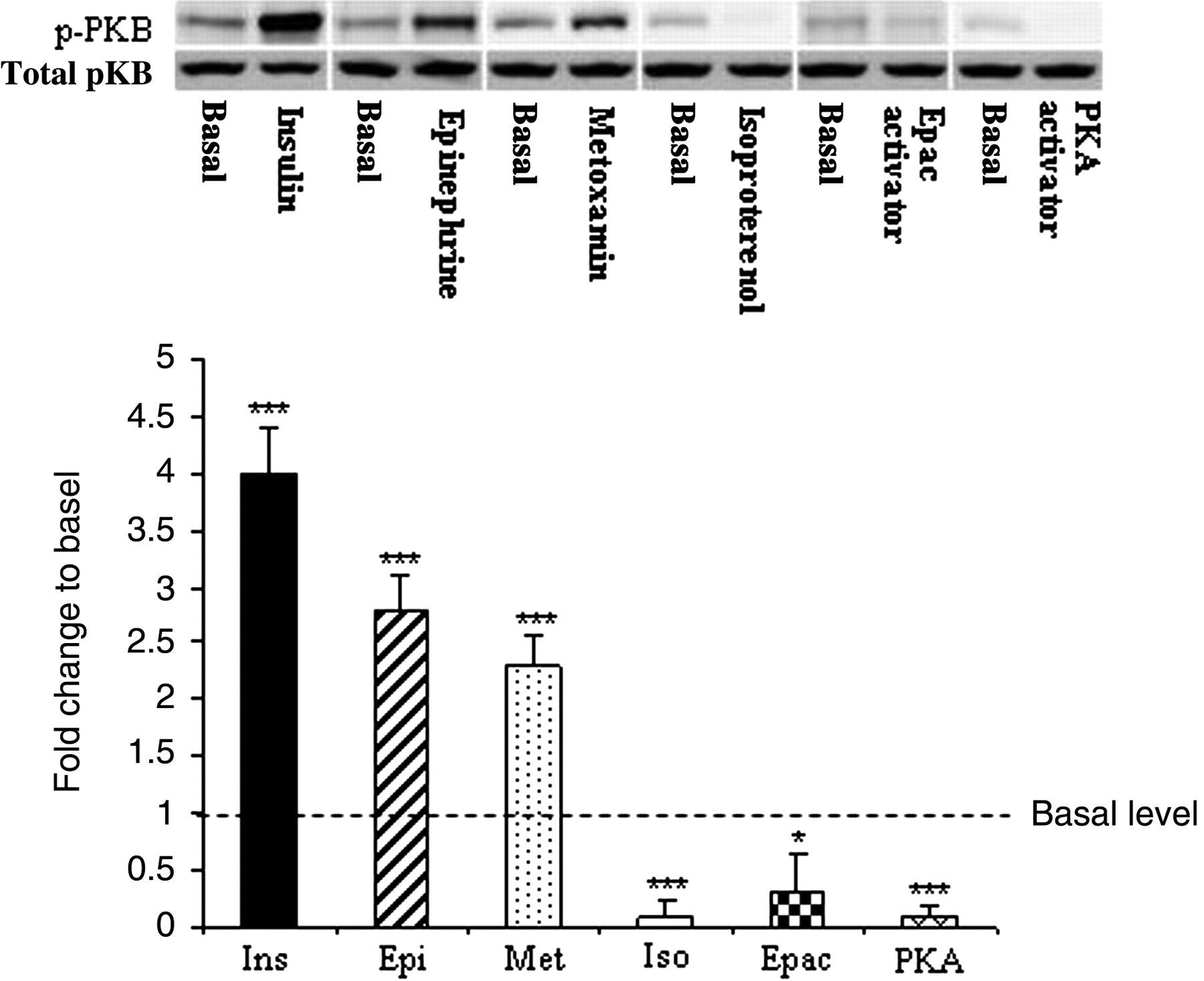
Epinephrine-stimulated glucose utilization is impaired in insulin resistance
Characteristics of HHTg rats
Fasted animals were deprived of food for 16 h. Next morning, half of the animals in each group obtained an oral single dose of glucose (3 g/kg b.w.) and were killed by cervical dislocation 30 min later (‘postglucose load’). Data are means ± SEM, n = 5–7
*P < 0.05; **P < 0.01; ***P < 0.001 HHTg versus control; # P < 0.05; ## P < 0.01; ### P < 0.001 postglucose load versus fasting
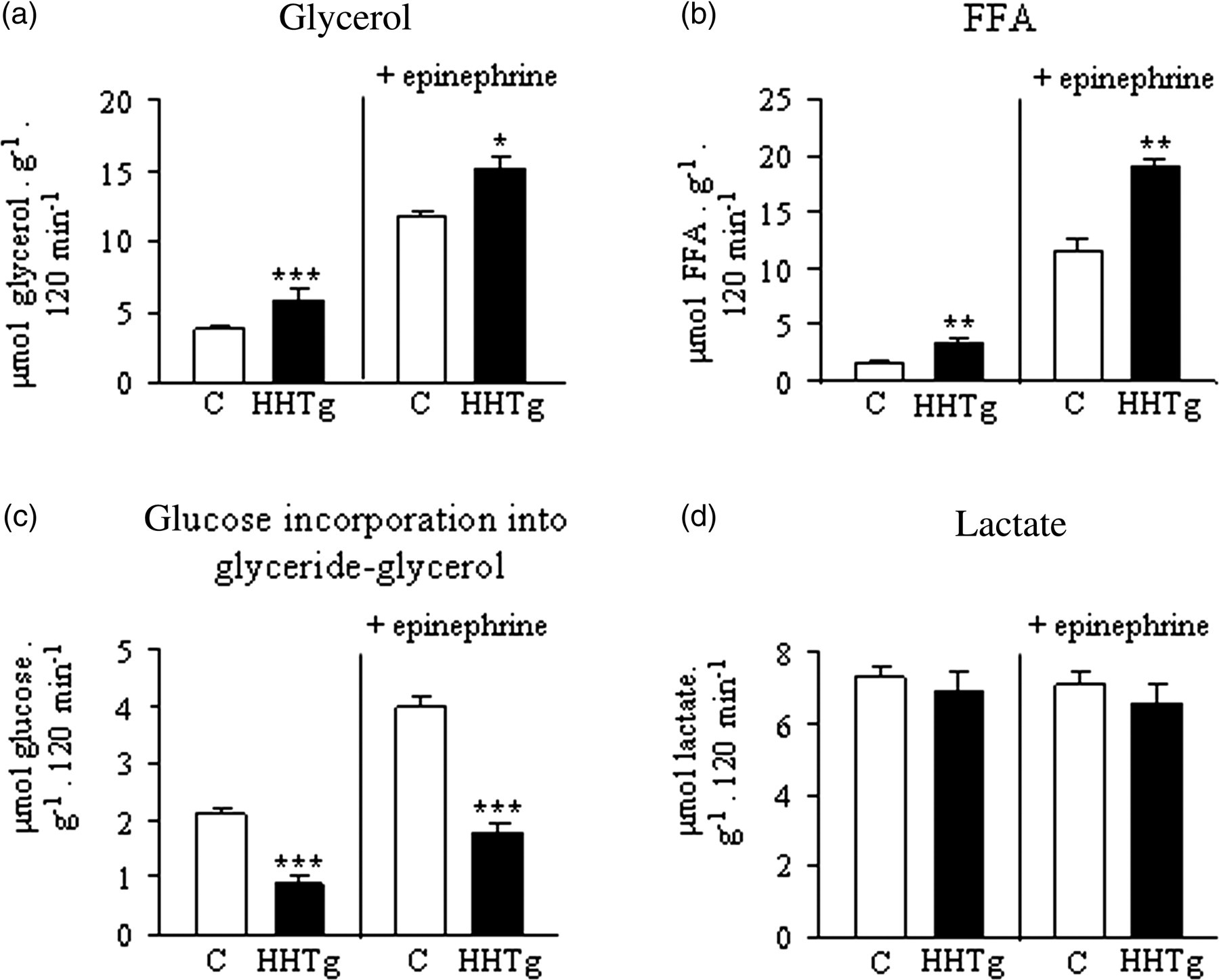
Effect of epinephrine on lipolysis and glucose utilization in insulin-resistant HHTg rats in vitro. (a) Glycerol content in the incubation medium; (b) FFA content in the incubation medium; (c) glucose incorporation into glyceride-glycerol moiety of TAG molecule; and (d) lactate content in the incubation medium. Distal parts of epidydimal adipose tissue were incubated in Krebs–Ringer bicarbonate buffer either alone or in the presence of 1 μmol/L epinephrine. Data are expressed as mean ± SEM, n = 6. Open bars, control rats; closed bars, HHTg rats. *P < 0.05, **P < 0.01, ***P < 0.001 HHTg versus control. FFA, free fatty acids

A hypothesis of the metabolic pathway by which epinephrine stimulates glucose uptake in EWAT. Activation of α 1-adrenoreceptors stimulates the phosphorylation of PKB and enhances the glucose uptake. This process is inhibited by wortmannin that implicates the involvement of PI3-K. Part of the G-3-P derived from the incorporated glucose is utilized for FFA esterification. β-Signalling (cAMP–PKA≫cAMP–Epac pathway) results in PKB dephosphorylation and lower glucose availability in the tissue. The final release of FFA from adipose tissue thus depends on the interaction of individual signalling pathways. AC, adenylyl cyclase; DGAT, diacylglycerol acyltransferase; EWAT, epididymal white adipose tissue; EPI, epinephrine; G-3-P, glycerol-3-phosphate; HSL, hormone-sensitive lipase; IR, insulin receptor; ISO, isoproterenol; MET, methoxamine; Rα, α-adrenoreceptor; Rβ, β-adrenoreceptor; PI-3K, phosphoinositide-3 kinase; PKA, protein kinase A; PKB, protein kinase B; N 6-Benzoyl-Camp, PKA activator; 8-pCPT-2′-O-Me-cAMP, Epac activator
Discussion
In the current study, we add to the existing knowledge on epinephrine control of glucose metabolism in EWAT in several ways. First, in EWAT, epinephrine stimulates glucose utilization specifically for re-esterification, thereby regulating the amount of FFA released from the tissue. Second, this effect of epinephrine involves PKB phosphorylation on Ser473. Third, β-adrenergic-dependent pathways have a negative effect on glucose utilization in EWAT. The activation of the cAMP–Epac effector system has only a moderately negative effect on glucose metabolism, while the cAMP–PKA-dependent pathway strongly inhibits PKB phosphorylation and epinephrine-mediated glucose availability. Finally, the positive effect of epinephrine on glucose metabolism is, at least partly, mediated by α 1-adrenoreceptor stimulation and this process is wortmannin-sensitive, which suggests the involvement of PI-3 kinase.
The metabolism of glucose is subject to a tight multilevel control. Epinephrine is generally considered as a counter regulator of the main glucose-regulating hormone insulin, 20 but recent evidence indicates a complex interaction between epinephrine and insulin signalling pathways in regulation of activities of several key enzymes like PI-3 kinase, PKB and GSK-3. 21–23 Nevertheless, the data concerning the effects of epinephrine on glucose uptake are contradictory and depends to a great degree on particular experimental design.
In the current study, we presented data showing that epinephrine alone stimulates PKB phosphorylation on Ser473 and that this effect is independent of insulin. Epinephrine further increased the glucose incorporation into glyceride-glycerol without decreasing the lactate production. We did not measure the glucose oxidation, but as the amount of glucose metabolized through this pathway in EWAT is approximately 1% that of lipid synthesis or lactate production 24 and the glycogen stores in EWAT are negligible (MC, unpublished results), it is reasonable to presume that the sum of lactate production and glucose incorporation into lipids approximates the glucose uptake.
Epinephrine activates several effector systems in the cell via two different classes of receptors (α- and β-adrenoreceptors) that are further divided into several functionally different subclasses (Figure 3). The diversity of published data concerning the epinephrine effect on glucose metabolism in various models stresses the cell type and organ specificity of these processes. 25–27 β-Adrenergic signalling is coupled with cAMP production via adenylate cyclase. It was recently established that two cAMP-dependent systems (cAMP–Epac and cAMP–PKA pathways) exist within most of the cell types. The development of cAMP analogs highly specific for either Epac 28 or PKA 29 provides an effective tool to study these two signalling pathways separately. cAMP signalling has been implicated in modulating PKB activity, but depending on the cell type, cAMP can either stimulate or inhibit PKB phosphorylation.
In skeletal muscle, cAMP–Epac-dependent signalling potentiated insulin-stimulated PKB phosphorylation but it had no insulin-independent effects, 19 while in adipocytes, Epac suppressed PKB activation. 30 Similarly divergent results are reported for the role of PKA in the control of PKB activity and glucose metabolism. 19,30–32 In our experiment, the direct stimulation of EWAT by Epac activator in vitro had a moderately negative effect on Ser473-PKB phosphorylation and it decreased total glucose utilization by 16%. Stimulation by β-adrenoreceptor specific agonist isoproterenol and by a PKA-specific activator produced a strong dephosphorylation of PKB and lowered glucose availability in adipose tissue. Collectively, our experiments suggest that in rat adipose tissue, β-adrenergic signalling is associated with down-regulation of PKB activity and glucose metabolism and that these effects are predominantly mediated by the cAMP–PKA pathway.
Epinephrine is a potent activator of lipolysis in vitro. Its prolipolytic effect is mediated via β-adrenoreceptor-coupled pathways. Data presented in this study showed that the cAMP–Epac pathway is not involved in the control of lipolysis. As expected, treatment with isoproterenol, PKA activator or epinephrine resulted in a significant stimulation of lipolysis. When the lipolysis was determined according to the glycerol release, the effects of PKA activator, epinephrine and isoproterenol were comparable, the latter being most effective. Nevertheless, the amount of FFA released from adipose tissue was significantly higher after specific β-adrenergic stimulation either by isoproterenol or by PKA activator compared with the effect of epinephrine. We suppose that this discrepancy indicates that epinephrine activates two opposite processes (FFA release and FFA re-esterification) at the same time while the β-adrenergic pathway stimulates only lipolysis.
Re-esterification of FFA requires adequate glucose availability. The role of α 1-adrenoreceptors in epinephrine-stimulated glucose uptake in WAT has been reported previously. 33 Using α 1 agonist methoxamine, we confirmed the positive effect of α 1 receptors on lactate production. To our knowledge, we were the first to show that that both epinephrine and specific α 1-adrenoreceptor activation leads to the phosphorylation of PKB. We further demonstrated that the α 1-dependent stimulation of glucose utilization in EWAT is wortmannin-sensitive, which implicates PI3 kinase in this (alpha-1) adrenoreceptor activation.
In accordance with previous observations, α 1 stimulation alone had no effect on lipolysis and only very moderate effect on glucose incorporation into glyceride-glycerol. The main observed effect of methoxamine was the increase of the lactate production. Epinephrine stimulates both lipolysis and glucose incorporation into glyceride-glycerol but it has no effect on lactate production. In our opinion, the key factor determining the pattern of glucose utilization is the presence or absence of lipolysis. Epinephrine stimulates concomitantly both α and β receptors in WAT and our data suggest that under these conditions, the surplus glucose mobilized by α 1 stimulation is used preferentially for partial re-esterification of the released fatty acids. The net FFA production thus depends on the interaction between α 1 and β signalling pathways. This hypothesis is supported by two observations. First, wortmannin significantly elevated the amount of released FFA after epinephrine stimulation in spite of the fact that the intensity of TAG breakdown was unchanged. Second, in HHTg rats that exhibit many aspects of metabolic syndrome including dyslipidemia and elevated serum FFA concentrations, 13,14 the positive effect of epinephrine on glucose uptake in EWAT in vitro was greatly diminished. It is important to stress that there is great variability in metabolic activity and (patho)physiological importance of various fat depots. 34,35 In mammals, adipose tissue is distributed in numerous depots throughout the body and an increasing number of reports emphasize site-specific physiological properties, including lipolysis and energy metabolism. 36–39 We studied EWAT because it represents the most commonly used fat depot in rat or mouse studies (including those dealing with the contribution of β- and α-adrenoreceptor-mediated pathways to the PKB phosphorylation and glucose utilization in WAT cited in this paper), is easily accessible and is relatively homogeneous. On the other hand, it is important to mention that EWAT exhibits some specific metabolic features. Brito et al. 40 reported that norepinephrine turnover in response to food deprivation or low temperature is lower in EWAT compared with retroperitoneal or inguinal fat depots. Recently, Caesar et al. 41 published interesting data showing that EWAT exhibits physiological zonation. A proximal part of EWAT closely adjoining to the testes was characterized by low de novo lipogenesis and a high content of polyunsaturated fatty acids, indicating that it may serve as a storage pool of unsaturated fatty acids for spermiogenesis. In contrast, in the distal parts of EWAT, both lipogenesis de novo and fatty acid composition were comparable with other fat pads tested in this study. Metabolic differences between the distal and proximal part of EWAT emphasize the need for accuracy in tissue sampling. In our study, we strictly used only the thin distal part of EWAT. Nevertheless, the applicability of our conclusions remains to be tested on other fat depots as well.
Taken together, we show that in EWAT, epinephrine controls opposite processes that could be ascribed to the different effectors' systems. β-Signalling (cAMP–PKA≫cAMP–Epac pathway) results in the PKB dephosphorylation and lower glucose availability in the tissue, whereas α 1 stimulation increases the glucose availability and utilization via mechanism involving PI-3 kinase and its downstream effector, PKB. The glucose is specifically used for FFA esterification. The final release of FFA from adipose tissue thus depends on the interaction of individual signalling pathways. The defect in α 1-dependent glucose transport may contribute to the elevation of serum FFA in pathological states like metabolic syndrome.
Footnotes
Acknowledgements
This study was supported by grant NS 9696-3 from the Ministry of Health of the Czech Republic.