
Abstract
Inhalation of chemical pollutants has been associated with a reduced immune response in humans. Inhalation of dust is a major route of exposure for one endocrine-disrupting chemical and suspected xenoestrogen, polybrominated diphenyl ethers (PBDEs); however, the impact of PBDEs on immune function is unclear. The objective of this study was to investigate the action of PBDEs on cytokine and eicosanoid release by alveolar macrophages and determine whether the effects are mediated via the estrogen receptor. The production of tumor necrosis factor (TNF)-α, interleukin (IL)-6, IL-1β, IL-10 and prostaglandin E2 (PGE2) by porcine alveolar macrophages exposed to different concentrations of the pentabrominated diphenyl ether mixture, DE-71, were measured; cells were also exposed to varying concentrations of 17β-estradiol and the selective estrogen receptor-modulating agent, tamoxifen. Cells exposed to PBDEs released significantly less pro-inflammatory cytokines (TNF-α and IL-6) and PGE2 compared with controls; IL-1β and IL-10 were not detected in the culture medium. Cells exposed to 17β-estradiol released significantly less TNF-α compared with controls, an effect that was reversed by the addition of tamoxifen; tamoxifen had no effect on the inhibition of TNF-α release by PBDEs. Although the suppression of TNF-α with DE-71 was similar to that of estrogen, the inhibitory effects of DE-71 were not found to be dependent on the estrogen receptor. Findings of this study suggest that chronic exposure to PBDEs suppressed innate immunity in vitro. Whether the immunosuppressant effects of PBDEs occur in vivo, remains to be determined.
Introduction
A reduced immune response in humans has been observed in association with exposure to environmental chemicals. For example, concentrations of dichlorodiphenyltrichloroethane (DDT) metabolites in adipose tissue have been found to be directly associated with a reduction in T-cell cytokine response in Australian women. 1 In children, exposure to high levels of environmental chemicals was found to be correlated with a greater prevalence of upper respiratory infections and bronchitis compared with children of same age and lower exposure. 2
The endocrine-disrupting chemical class and suspected xenoestrogen, polybrominated diphenyl ethers (PBDEs), are a class of synthetic compounds similar in structure to DDT and polychlorinated biphenyls (PCBs), which are used extensively as flame retardants in the manufacturing of common household and commercial products including: plastics in electrical appliances, televisions and computers; and foams, carpets and upholstery found in furniture, automobiles and airplanes. 3 Unlike the long-banned DDT and PCBs, some formulations of PBDEs continue to be produced for use in domestic products. Consequently, either as a result of bioaccumulation and/or debromination of products, elevated levels of PBDEs continue to be detected in human tissue. 4,5
It is estimated that over 80% of total PBDE intake occurs by inhalation of house dust. 6 As a result of chronic PBDE exposure via inhalation, PBDEs may compromise the activation of innate immunity, an essential immune mechanism for maintaining lung homeostasis and the prevention of chronic respiratory conditions. However, despite the apparent significant contribution of PBDE exposure via inhalation, the immune response to PBDEs is not well characterized.
Alveolar macrophages account for approximately 93% of the macrophages in the lung 7 and because of their position in the lung and pleural cavities, interact continuously with the inhaled environmental antigens that are ubiquitous in ambient air. 8,9 Consequently, activated alveolar macrophages are considered the main regulators of lung homeostasis. 10 Macrophages have a wide repertoire of chemical signals as well as receptors to communicate with other cells. For example, tumor necrosis factor (TNF)-α, interleukin (IL)-6 and IL-1β are pro-inflammatory products, phenotypic of classically activated macrophages. Furthermore, macrophages have been shown to produce large quantities of the eicosanoid, prostaglandin E2 (PGE2), which can have both pro-inflammatory and anti-inflammatory effects. In addition, in certain conditions, macrophages can be alternatively activated and may produce the anti-inflammatory cytokine, IL-10. In addition, alveolar macrophages also express the estrogen receptor (ER), and have been shown to be regulated by estrogen. 11
In view of the limited studies examining the effects of PBDEs on immune response, 12–14 and since alveolar macrophages play a pivotal role in immune function, we were interested in examining the effects of PBDEs on macrophage function in vitro. Specifically, the primary objective of the following study was to determine the effects of PBDEs at different concentrations, on the release of pro-inflammatory cytokines and PGE2 by activated alveolar macrophages.
Materials and methods
Materials
All materials were purchased from Sigma-Aldrich (St Louis, MO, USA), unless indicated otherwise. 3D4/31 Porcine alveolar macrophages were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). The DE-71 formulation (a mixture of brominated diphenyl ether-99, -47, -100 and -153) of pentabrominated diphenyl ether was purchased from Cambridge Isotope Laboratories, Inc (Andover, MA, USA). The 17β-estradiol was a gift from Dr Paul Tsang (Department of Molecular, Cellular, and Biomedical Sciences; University of New Hampshire). Enzyme-linked immunosorbent assays (ELISAs) were acquired from R&D Systems, Inc (Minneapolis, MN, USA) to measure the following: TNF-α, IL-6, IL-1β, PGE2 and IL-10. Cell viability was confirmed by lactate dehydrogenase (LDH; Cayman Chemical Company, Ann Arbor, MI, USA). The Roswell Park Memorial Institute (RPMI)-1640 and non-essential amino acids were purchased from ATCC; fetal bovine serum (FBS) and porcine serum were purchased from MP Biomedicals (Solon, OH, USA).
Cell culture protocol
For each experiment, a cryovial of passage #22, 3D4/31 porcine alveolar macrophages was removed from storage, seeded into T75 flask(s) in RPMI growth medium supplemented with 10% FBS and 1% non-essential amino acids, and incubated at physiological conditions (5% CO2, 37°C). At 60–70% confluency, cells were passaged and seeded into 24-well plate(s) (300,000 cells/well) in RPMI growth medium, supplemented with 10% porcine serum and 1% non-essential amino acids, and incubated (5% CO2, 37°C) for 24 h. After 24 h, cells were activated by inoculation with phorbol 12-myristate 13-acetate (PMA). The activating stimulus was prepared by solubilizing PMA (0.01 mg/mL) and ionomycin (0.05 mg/mL) in 1% dimethyl sulfoxide (DMSO). In addition to the activating stimulus, different concentrations of the experimental treatments and respective vehicle controls were introduced to the wells, and the plate(s) were incubated (5% CO2, 37°C). After a six-hour period, conditioned media was removed, centrifuged and stored at −80°C until further analysis. This protocol was adapted from that reported by others. 15
PBDE exposure and cytokine and eicosanoid release
Stock solutions were prepared by solubilizing the DE-71 mixture in DMSO at seven concentrations (0.1, 1, 10, 100, 1000, 150 and 2000 ng/mL). Replicate of cell samples (n = 5–9) were exposed to PMA and one of the seven different PBDE concentrations or a control treatment (0 ng/mL). The final DMSO concentration in the solutions was less than 1%. The following cytokines and prostaglandin (PGE2) were measured using ELISA: TNF-α, IL-6, IL-1β, IL-10 and PGE2, respectively. ELISA methods were conducted according to the manufacturer's instructions. In brief, 96-well plates were coated with capture antibody and incubated overnight. Wells were aspirated and washed three times with wash buffer; blocking buffer was added to each well and plates were incubated for one hour. After a series of wash steps, 50 μL of assay diluent and 50 μL of standard, control or sample was added to each well; plates were incubated for two hours. Wells were aspirated and washed five times with wash buffer. Porcine conjugate of the cytokine or eicosanoid was added to each well and plates were incubated for two hours. Wells were washed and substrate solution was added to each well; plates were incubated for 30 min removed from light. Stop solution was added to each well and absorbance was measured at 450 nm using a microplate reader. All incubations were conducted at room temperature.
Cell viability
To assess cell viability, cells were seeded in a 96-well plate at a density of 100,000 cells/well in 120 μL RPMI growth media, supplemented with 1% FBS and 1% non-essential amino acids. After a 24-h incubation period (5% CO2, 37°C), experimental treatments were added to the wells and the plate was incubated (5% CO2, 37°C) for six hours. Experimental treatments included: (1) culture medium; (2) DMSO (1%); (3) PMA (0.01 mg/mL) and ionomycin (0.05 mg/mL) in 1% DMSO; and (4–10) PMA + seven different concentrations of DE-71 (0.1, 1, 10, 100, 1000, 1500, 2000 ng/mL). After six hours, the plate was centrifuged at 400×g for five minutes. In the corresponding wells of a new 96-well plate, standards were added in triplicate (n = 3) and 100 μL of supernatant from each well of the centrifuged plate was added (n = 8). In accordance with the manufacturer's protocol, 100 μL of reaction solution (9.6 mL of assay buffer + 100 μL of the following: NAD+ , lactic acid, a tetrazolium salt [iodonitrotetrazolium chloride] and reconstituted diaphorase) was added to each well and the plate was placed on an orbital shaker and incubated at room temperature with gentle shaking for 30 min. The absorbance at 490 nm was then read using a microplate reader.
Estrogen exposure and TNF-α release
Five different physiological and supra-physiological concentrations of 17β-estradiol were prepared by diluting the 640 μg/mL of stock solution in ethanol (12.5, 25, 50, 250 and 1000 pg/mL). The final ethanol concentration in the solutions was less than 1%. Cells were exposed to PMA and each of the five different concentrations of 17β-estradiol (n = 5/treatment). The release of TNF-α was measured via ELISA as described above.
Tamoxifen exposure and TNF-α release
A stock solution was prepared by solubilizing the tamoxifen in DMSO, so that the final concentration was 10 μmol/L. Cells were exposed to PMA and 0.1 and 1 ng/mL of DE-71 and 12.5 pg/mL of 17β-estradiol with or without the addition of tamoxifen (n = 9/treatment). The release of TNF-α was measured via ELISA as described above.
Statistical analysis
Statistical analysis was performed using commercially available statistical software (SPSS 18; SPSS Inc, Chicago, IL, USA). Treatment effects were determined by analysis of variance using the general linear model with a Bonferroni adjustment. Results are presented as mean ± standard error (n = 5–9/treatment) from at least three independent experiments. Differences with a P value of less than 0.05 were considered statistically significant.
Results
For all experiments, neither cytokines nor eicosanoids were detected in the absence of PMA. Therefore, only conditions in which measurable levels of product were detected in response to PMA stimulation are reported.
PBDE exposure and cytokine and eicosanoid release
Alveolar macrophages treated with PMA alone released 340 ± 23 pg/mL of TNF-α. Cells exposed to each of the seven different concentrations of DE-71 (0.1, 1, 10, 100, 1000, 1500 and 2000 ng/mL) + PMA released significantly less TNF-α compared with cells exposed to PMA alone (Figure 1). In addition, cells exposed to 1 and 10 ng/mL concentrations of DE-71 + PMA released significantly less TNF-α compared with cells exposed to 1000 ng/mL of DE-71 + PMA. Furthermore, the cells treated with 1500 and 2000 ng/mL of DE-71 + PMA had significantly lower TNF-α concentrations than cells treated with 0.1 and 1000 ng/mL of DE-71 + PMA.
Effects of polybrominated diphenyl ethers on tumor necrosis factor (TNF)-α release by activated alveolar macrophages. All treatments included phorbol 12-myristate 13-acetate (PMA) (10 μL/mL). Values are mean ± SE: n = 6 (0.1–2000 ng/mL) and n = 9 (0 ng/mL). Means with different letters are significantly different (P < 0.05)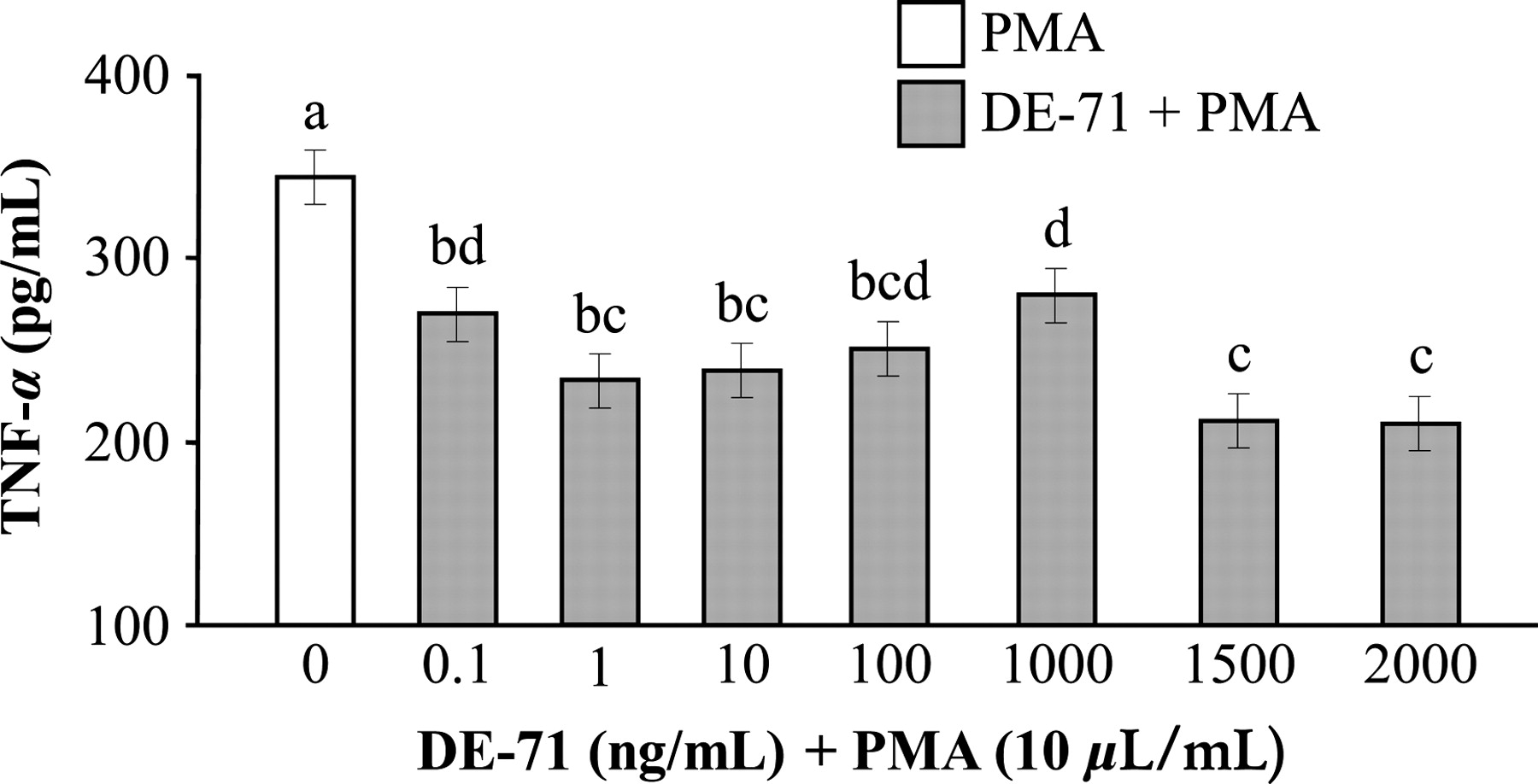
The concentration of IL-6 was also significantly lower in cells treated with 0.1, 1, 10, 100 and 1000 ng/mL of DE-71 + PMA compared with PMA alone (Figure 2). All treatment conditions of DE-71 (0.1, 1, 10, 100 ng/mL) + PMA produced lower concentrations of IL-6 than cells exposed to 1000 ng/mL of DE-71 + PMA.
Effects of polybrominated diphenyl ethers on interleukin (IL)-6 release by activated alveolar macrophages. All treatments included phorbol 12-myristate 13-acetate (PMA) (10 μL/mL). Values are mean ± SE: n = 5/treatment. Means with different letters are significantly different (P < 0.05)
Control cells treated with PMA alone released 1010 ± 101 pg/mL of PGE2. In comparison, significantly less PGE2 was released when cells were treated with either 0.1, 1, 10 or 1000 ng/mL of DE-71 (Figure 3).
Effects of polybrominated diphenyl ethers on prostaglandin E2 (PGE2) release by activated alveolar macrophages. All treatments included phorbol 12-myristate 13-acetate (PMA) (10 μL/mL). Values are mean ±SE: n = 6/treatment. Means with different letters are significantly different (P < 0.05)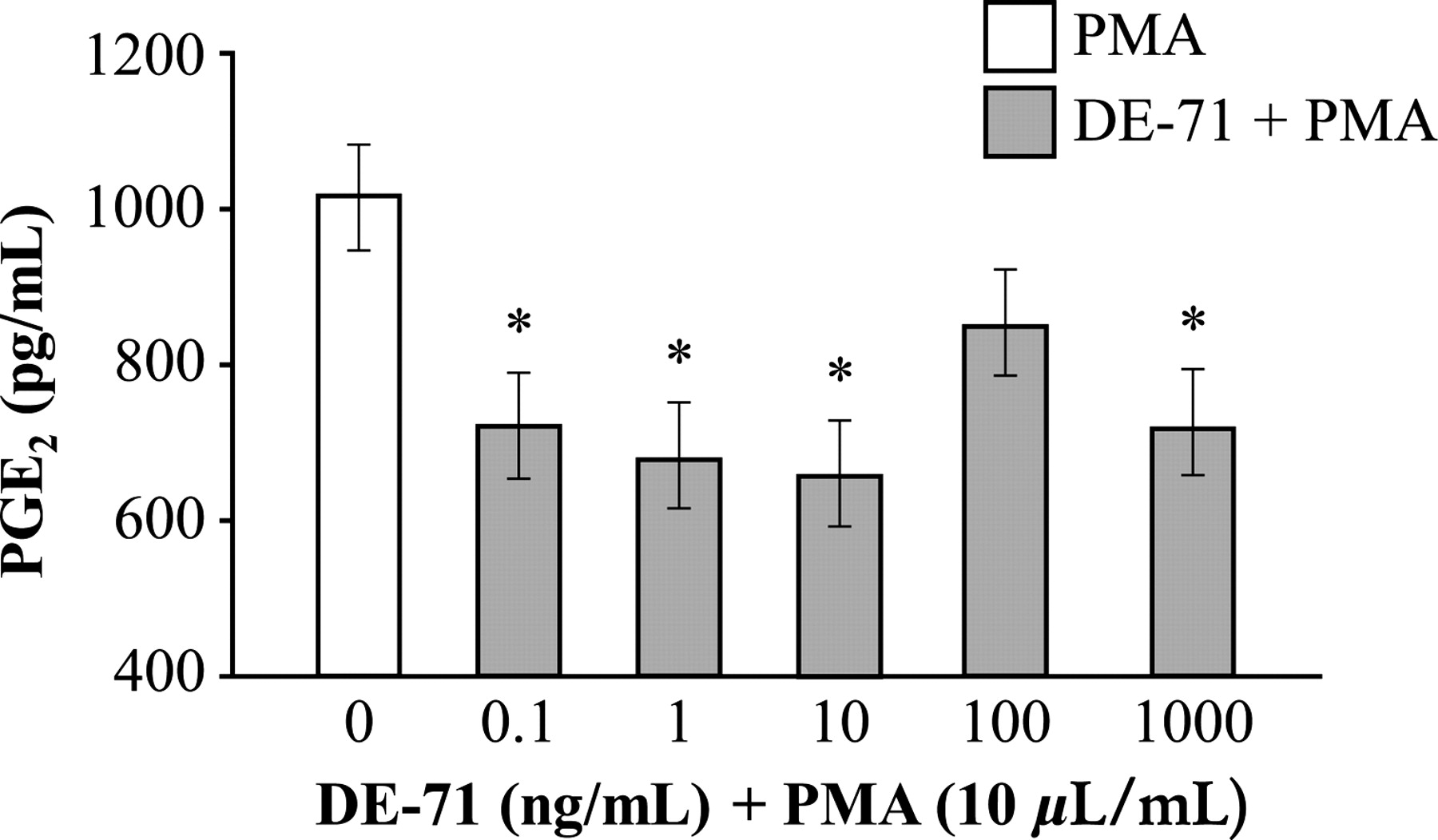
IL-1β and IL-10 were not detected in any treatment or control conditions (data not shown).
PBDEs and cell viability
LDH is a cytosolic enzyme released into the culture medium when the cell is damaged due to either apoptosis or necrosis, and can therefore be used as an indicator of cytotoxicity. Figure 4 shows the LDH cytotoxicity results. Alveolar macrophages in culture medium released 5.94 ± 0.3 mU/mL of LDH. Cells exposed to 1, 100 and 1000 ng/mL of DE-71 + PMA released significantly less LDH (3.96 ± 0.1, 3.24 ± 0.2 and 3.38 ± 0.4 mU/mL, respectively) compared with cells in culture medium alone, but the same as cells exposed to PMA alone. When exposed to the highest concentration of DE-71 (2000 ng/mL) + PMA, cells released significantly more LDH (8.83 ± 0.7 mU/mL) compared with all other experimental and control treatments. Also in the presence of 1500 ng/mL of DE-71 + PMA, cells released significantly more LDH (5.95 ± 0.6 mU/mL) compared with cells exposed to either 1, 100 or 1000 ng/mL of DE-71 + PMA, but significantly less LDH compared with cells exposed to 2000 ng/mL of DE-71 + PMA.
Lactate dehydrogenase (LDH) cytotoxicity. Cells were exposed to culture medium (CM); 1% dimethyl sulfoxide (DMSO); phorbol 12-myristate 13-acetate (PMA) (0.01 mg/mL) and ionomycin (0.05 mg/mL) in 1% DMSO; and PMA + seven different concentrations of DE-71. Values are mean ± SE: n = 8/treatment. Means with different letters are significantly different (P < 0.05)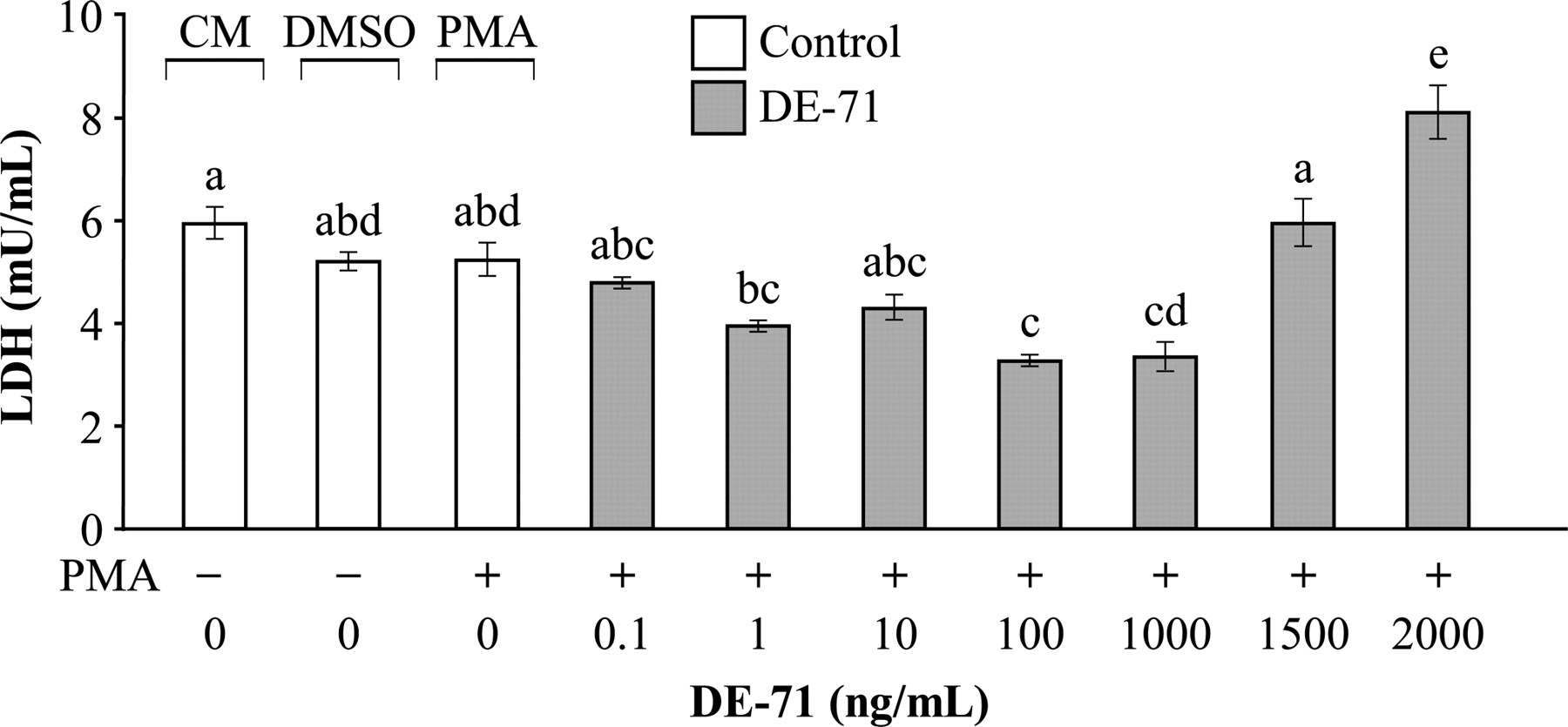
Estrogen exposure and TNF-α release
To establish the effects of estrogen on TNF-α release, alveolar macrophages treated with varying concentrations of estrogen and TNF-α were measured. Control cells treated with PMA secreted 204 ± 3.6 pg/mL of TNF-α. Cells treated with one of five different concentrations of 17β-estradiol (12.5, 25, 50, 250 and 1000 pg/mL) + PMA released significantly less TNF-α compared with control cells (169 ± 3.5, 162 ± 2.5, 159 ± 1.4, 164 ± 3.7 and 169 ± 3.1 pg/mL, respectively) (Figure 5).
Effects of 17β-estradiol (17β-E2) on tumor necrosis factor (TNF)-α release by activated alveolar macrophages. All treatments included phorbol 12-myristate 13-acetate (PMA) (10 μL/mL). Values are mean ± SE: n = 5/treatment. Means with different letters are significantly different (P < 0.05)
Tamoxifen exposure and TNF-α release
To determine whether the effects of PBDEs on cytokine release were mediated via the ER, we used the selective ER modulator, tamoxifen. Figure 6 shows the effects of tamoxifen on the release of TNF-α by alveolar macrophages exposed to different treatment conditions. Cells treated with tamoxifen + PMA released significantly less TNF-α compared with PMA alone (102 ± 3.8 versus 120 ± 2.5 pg/mL, respectively). PMA + 0.1 or 1 ng/mL of DE-71 resulted in a significant reduction in TNF-α concentration compared with PMA alone (98 ± 3.5 and 97 ± 2.0 pg/mL, respectively); the suppression remained and tended to decrease further with the addition of tamoxifen (not significant); there were no differences in TNF-α between treatment conditions, DE-71 + PMA and tamoxifen + PMA.
Effects of tamoxifen (TAM) on tumor necrosis factor (TNF)-α release by activated alveolar macrophages. All treatments included phorbol 12-myristate 13-acetate (PMA) (10 μL/mL). Values are mean ± SE: n = 9/treatment. Means with different letters are significantly different (P < 0.05). 17β-E2, 17β-estradiol
When PMA + 17β-estradiol were added to the culture medium, there was a significant reduction in TNF-α concentration compared with PMA alone (97 ± 2.9 pg/mL). The suppressing effect of 17β-estradiol on TNF-α was reversed when tamoxifen was added in combination with 17β-estradiol + PMA. In contrast, the release of TNF-α in response to 17β-estradiol + PMA was similar to TNF-α concentrations of cells treated with either concentration of DE-71 + tamoxifen + PMA or tamoxifen + PMA (112 ± 3.6 pg/mL). TNF-α concentration following 17β-estradiol + PMA + tamoxifen, however, was higher than following either treatment of DE-71 + PMA + tamoxifen.
Discussion
PBDE and macrophage function
In this study, we report for the first time, in vitro evidence that exposure of alveolar macrophages to the PBDE mixture DE-71, at concentrations not associated with cell toxicity, suppresses the release of pro-inflammatory products involved in normal innate immune function. As alveolar macrophages play a pivotal role in both innate immunity and adaptive immunity, any disruption in normal pro-inflammatory signaling by macrophages related to PBDE exposure could compromise maintenance of respiratory health.
PBDEs are not fixed in the polymer product by chemical bonding and therefore, products containing PBDEs spontaneously emit bromine atoms into the ambient air with continued use and disuse over time. Owing to their position in the interstitial spaces of the bronchi and alveoli in the vascular compartment and in the alveolus, alveolar macrophages are interacting continuously with the inhaled environmental antigens that are ubiquitous in ambient air. 8,9 Commensurate with their role, macrophages have a wide repertoire of chemical signals (i.e. cytokines and eicosanoids) as well as receptors (e.g. ERα and ERβ) to communicate with other immune cells to effectively clear any potentially infectious antigens.
The suppression of TNF-α, IL-6 and PGE2 in this study may be explained, in part, by disrupted signaling of nuclear factor-kappa B (NF-κB). Pro-inflammatory cytokines TNF-α, IL-6 and IL-1β are regulated through the activation of the transcription factor, NF-κB.
Many stimuli have been shown to activate NF-κB, including TNF-α and protein kinase C activators, such as PMA and lipopolysaccharide. 16 The eicosanoid, PGE2, is a potent inflammatory mediator, derived from arachidonic acid through the cyclooxygenase 2 enzyme, which is also regulated by NF-κB. 17 Our data suggest that the inhibitory effects of PBDEs observed was not the result of cell toxicity, as LDH released by macrophage cells in response to PBDE + PMA treatments (with the exception of 2000 ng/mL) was either similar to, or lower than, the PMA treatment alone.
Estrogen-like effects of PBDEs
The endocrine-disrupting potential of PBDEs has largely been studied with regard to their ability to disrupt thyroid hormone homeostasis. In addition, many environmental pollutants have proven to be xenoestrogens. 18–21 Owing to the structural similarity between PBDEs and other environmental pollutants, such as DDT and PCBs, it is highly likely that PBDEs are estrogenic as well. 22–24 Indeed, Meerts et al. 22 report evidence that several PBDE congeners are agonists of the ERα and ERβ receptors; and additional evidence has since followed of the weak estrogen-like activity of PBDEs. 25,26
To determine whether the effect of PBDEs on TNF-α release was similar to that of estrogen, we compared the effects of PBDEs and 17β-estradiol on TNF-α release. Cells exposed to PBDEs showed a dose-related U-shaped curve, where at low concentrations (0.1 and 1 ng/mL of DE-71), there was a steady decline in TNF-α, and as the concentration of DE-71 increased (10, 100 and 1000 ng/mL of DE-71), the production of TNF-α increased, although it remained lower than PMA alone, throughout. These findings are consistent with those reported by others. 27,11
Possible receptor pathways of PBDE action
In this study, release of TNF-α by alveolar macrophage was suppressed in response to treatment with the PBDE mixture DE-71; this effect was observed in response to treatment with 17β-estradiol, and also tamoxifen alone. When tamoxifen and 17β-estradiol treatment were given together, the inhibitory effect of 17β-estradiol on TNF-α release disappeared. Tamoxifen, however, failed to disinhibit the suppressing effects of PBDEs on release of TNF-α. These findings suggest that the inhibitory effect of PBDEs on TNF-α release is not dependent on ERα or ERβ receptors 26 and may act through alternative pathways, such as the recently discovered steroid and xenobiotic receptor (SXR). 28
SXR is a nuclear receptor that has been described as a xenobiotic sensor that regulates the clearance of xenobiotics in the liver and intestine. 29 Interestingly, SXR has been shown to be highly expressed in alveolar macrophages. 30 Many endogenous compounds, including estrogen and exogenous compounds, such as tamoxifen, as well as PBDEs, are known activators of SXR. 29 Tamoxifen has been shown to be a potent activator of the SXR, 31 which may explain the apparent decrease in TNF-α by tamoxifen alone, as well as by PBDE + tamoxifen that were observed in this study.
In summary, the observed suppression in pro-inflammatory cytokine and eicosanoid release by PMA-activated porcine alveolar macrophages exposed to DE-71 is a novel finding.
We show that the suppression seen with DE-71 is similar to that of estrogen; however, our results demonstrate that the inhibitory effects of DE-71 are not dependent on the ER. It is important to note, however, that these findings are limited to that of a clonal cell line and in vitro conditions. Whether the immunosuppressant effects of PBDEs occur in vivo, remains to be determined. In addition, there are 209 different congeners of brominated diphenyl ethers (BDEs), five of which account for 90% of the PBDEs found in human tissues (BDE-47, -99, -100, -153, -154). 32 In this study, the effect on immune cell function was produced using the DE-71 formulation (a mixture of BDE-99, -47, -100 and -153). It will be important to determine in future studies which congener(s) of PBDEs are most biologically active.
Proper functioning of the macrophage through chemical signals such as cytokines and eicosanoids is essential for the innate immune response and host defense. Our results suggest that chronic exposure to PBDEs could compromise the effectiveness of alveolar macrophage signaling, that in the short term, could lead to increased risk of respiratory infection, and in the long term, possibly to autoimmunity or allergic disease. 33 Clearly, more research in this area is warranted.
Footnotes
ACKNOWLEDGEMENTS
The authors thank Anne Ronan for her excellent technical support throughout the study, as well as Dr Paul Tsang for his generous gift of 17β-estradiol. This study was supported, in part, by the New Hampshire Agriculture Experiment Station, Project 11H495. This is Scientific Contribution Number 2434 from the New Hampshire Agricultural Experiment Station.