
Abstract
Selenium (Se) is an exogenous antioxidant that performs its role via expression of selenoproteins. Pathological changes of the structure of the vessel wall, elastin turnover and collagen production may lead to increased stiffness of the vessels with decreased blood flow to the peripheries. The level of anti-elastin antibodies (AEABs) may give information for elastin metabolism. The aim of the study is to investigate the influence of Se intake on the vessel wall changes and production of AEABs in spontaneously hypertensive rats (SHR). Twenty-four male, 32-week-old SHR were used, divided into three groups, G1, G2 and G3. Before blood and morphological testing, G1 received a low-Se diet for eight weeks, G2 received a diet with adequate Se content and G3 received a diet with Se supplementation. The Se nutritional status was assessed by determination of glutathione peroxidase-1 (GPx-1) activity in whole blood, using the ‘Ransel’ kit. The rats from group G3 showed higher GPx-1 activity and lower level of AEABs than the other groups (P = 0.021), and the aortic wall histology showed slight degenerative changes compared with other rats. A low-Se diet caused severe changes to the aortic wall's ultrastructure, whereas Se supplementation slowed the changes down. The morphometry revealed a thicker abdominal aortic wall in rats of G1 compared with the other groups, and reduced thickness of the wall of the left coronary artery in G3 compared with the other groups (P < 0.05). Our results have shown that low Se intake leads to severe changes in the vessel walls in SHR, whereas selenium supplementation slows down the elastin degradation and degenerative changes of the vessel walls.
Introduction
It is considered that the physiological effects of selenium (Se) are due mainly through the functions of selenoproteins (enzymes that incorporate Se in the form of one or more selenocysteine residues) as their expression are controlled by 25 selenoprotein genes. 1 Tissue expression of these enzymes depends on the daily Se intake, which occurs mainly by food. It has been established that for normal growth and reproduction in mammals, a diet containing 0.1 μg Se/g of food is enough. 2–4 Se-dependent cellular glutathione peroxidase-1 (GPx-1) is the most abundant intracellular isoform of the GPx antioxidant enzyme family, and it plays a major role in the control of reactive oxygen species (ROS), which participate in atherogenesis and in the pathogenesis of hypertension. As continuously exposed to a blood flow, endothelial cells are the primary target of an oxidant-induced injury. 5,6 Many studies are focused on the role of trace elements for normal endothelial function, but there is not enough evidence about their effect on the metabolism of the proteins of the underlying connective-tissue matrix. Elastin is the main component of the vessel wall. Pathological changes in elastin turnover and increased production of collagen may lead to a disturbance of the vessel's structure, increasing the stiffness of the vessel walls and decreasing blood supply to the organs. Autoantibodies to α-elastin (an elastin breakdown product) and tropoelastin (an elastin precursor) are found in the serum of healthy human individuals and correlate with their respective serum peptide concentrations. 7,8 They are considered as physiological autoantibodies and are assumed to be a part of a homeostatic mechanism, which clears altered elastin structures by in situ destruction or by opsonization of the products of degradation. Serum antibodies to tropoelastin (reflecting elastin synthesis) and α-elastin (reflecting elastin destruction) appear to correlate with the production and breakdown of the elastic tissue. An increased elastin turnover can be seen in several disorders, but its role is still not well known. In patients with systemic lupus erythematosus, scleroderma and polyarteritis nodosa, a marked increase in pathological anti-α-elastin autoantibodies level was found which suggests their role in the pathogenesis of autoimmune alterations. 9,10
Spontaneously hypertensive rats (SHR) are the best appropriate genetic model for studying essential hypertension in humans. These rats develop hypertension between the fourth and tenth week of life. It is evident that the older the SHR are and the longer the duration of their hypertension, the more difficult it is to induce a regression of hypertensive structural changes in their hearts and vessels. 11
There are no data about the influence of different levels of Se intake on the vessel wall structure and serum concentration changes of anti-α-elastin autoantibodies in SHR. Because of the close relations between the peptides and the presence of specific antibodies, the level of anti-elastin antibodies (AEABs) may give information about elastin metabolism and alterations to the vessels. 12
The aim of this study is to investigate the influence of different levels of Se intake on the changes to the vessel wall and serum concentrations of AEABs in SHR.
Materials and methods
The experiment was performed in accordance with the Animal Welfare Act Regulations and permission was obtained from the local ethical committee.
Experimental animals
Twenty-four male SHR (Okamoto–Aoki strain), 24 weeks old, were put on diets with different Se contents for eight weeks. They were divided into three groups: G1 (n = 8) – on a low-Se diet (0.05 μg Se/g of food); G2 (n = 8) – on an adequate Se diet (0.11 μg Se/g); and G3 (n = 8) – on a high-Se diet (0.25 μg Se/g). Se was given as selenomethionine. The systolic blood pressure was measured indirectly using a tail cuff (194 ± 6 mmHg). The rats were placed in the single chambers and had water and food ad libitum. The daily intake of food was checked for each rat.
Assessment of selenium nutritional status
At the age of 32 weeks, under anaesthesia of pentobarbital sodium (40 mg/kg [intraperitoneally]), blood was taken from the abdominal aorta. The Se nutritional status was assessed by measuring GPx-1 activity in whole blood, using the ‘Ransel’ kit according to the protocols of Randox Laboratories Ltd (Antrim, UK) in the Central Clinical Laboratory of the University Hospital of Pleven. The hemoglobin (Hb) content of the blood was determined as well.
Determination of AEABs
AEABs were determined by the method of indirect home-made enzyme-linked immunosorbent assay (ELISA) in the Immunological Laboratory of the Biology Department of Medical University, Pleven. The plates (Greiner Microlon, Monroe, NC, USA) were coated with 100 μl rat lung α-elastin (Elastin Products Company, Owensville, MO, USA) (10 μg/mL) for two hours at room temperature and over the course of one night at 4°C. After washing with phosphate-buffered saline + 0.05% Tween-20 (PBS–Tween), the wells were blocked with 100 μL 0.1% bovine serum albumin in PBS–Tween-20 for one hour at 37°C. The tested serum samples (100 μL) diluted 1:5 were dropped, and incubated for one hour at 37°C. After washing, 100 μL anti-rat IgG peroxidase conjugate (Sigma-Aldrich, St Louis, MO, USA), at a dilution of 1:6400, were added. Incubation was performed for one hour at 37°C. After washing and incubation with substrate solution (0.8 mg/mL in 0.05 mol/L citrate buffer, рН 5.0, containing 0.01% H2O2), the reaction was stopped at the 20th minute by 50 μL 8N H2SO4. The absorptions were read at 492 nm by an automated ELISA reader. The following controls were used: (1) substrate control: assay buffer only, washing solutions and substrate added to the polystyrene wells coated with antigen α-elastin; (2) detection antibody control: detection antibody added directly to the wells coated with antigen; (3) negative control to assess the specificity of the reaction: antigen replaced by human albumin solution and serum samples; and the (4) positive control: the tested sample replaced by antibodies to rat α-elastin in assay buffer. All samples were analyzed in triplicates and averaged.
Histological techniques
The aortas of the rats, separated into two parts – thoracic and abdominal – were extirpated and then fixed in a 10% solution of formaldehyde, embedded into paraffin blocks. The hearts were also extirpated and cut below the auricula. Longitudinal slices (4 μm thick) were prepared and stained by hemalaun–eosin. The morphological changes were examined and described under the light microscope. Measurements of the thickness of the aortic walls and left coronary artery of each rat were performed at 10 points (including the thickest and the thinnest areas), using an ocular micrometer at the Laboratory of Pathomorphology of the National Center of Public Health Protection, Sofia, Bulgaria.
Electron microscopy (EM) analysis was performed, using an electron microscope (Hitachi HS7S, Tokyo, Japan), after appropriate fixation and staining of the aortas and hearts.
The computer statistics program SPSS, V.15 (SPSS Inc, Chicago, IL, USA) was used for data analysis. The tests used for conclusions were: Fisher's F test (analysis of variance) and multiple range tests (least significant difference) at the P < 0.05 confidence level. For simple regression analysis, the P < 0.1 confidence level was used.
Results
The GPx-1 activity of whole blood in rats from G1, as the means ± SEM, was 453 ± 73 U/g Hb; in G2 – 450 ± 73 U/g Hb; and in G3 – 559 ± 78 U/g Hb. The rats from G3 had higher GPx-1 activity than the other groups (P = 0.021) (Figure 1).
GPx-1 activity of whole blood in G1, G2 and G3 as the means ± SEM (U/g Hb). GPx-1, glutathione peroxidase-1; Hb, hemoglobin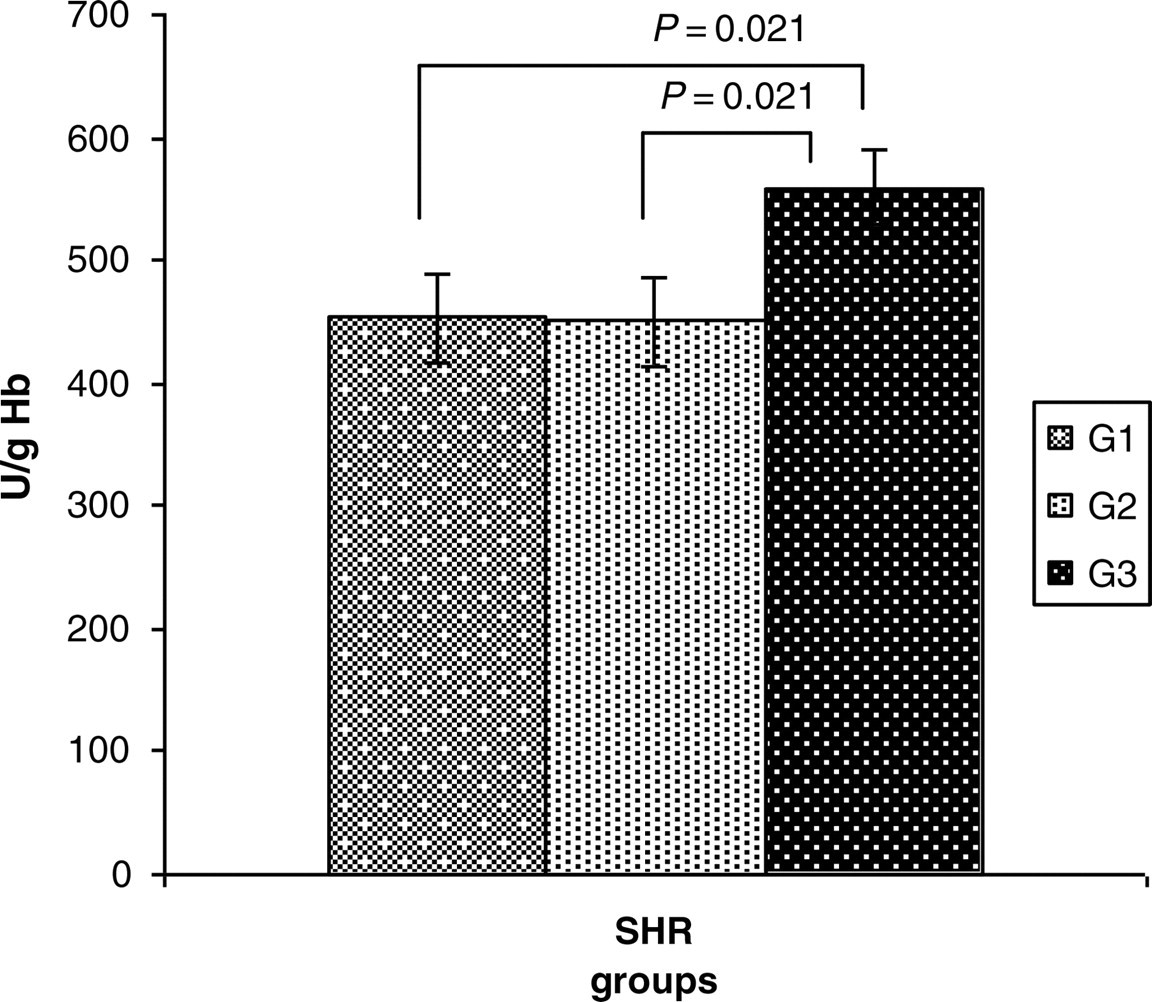
Serum concentrations of AEABs as the means ± SD were lower in rats from G3 than the other groups (P = 0.021): G1 = 0.1173 ± 0.03; G2 = 0.1172 ± 0.01; and G3 = 0.0922 ± 0.02 OD 492 nm (Figure 2).
Serum concentration of anti-elastin antibodies in G1, G2 and G3 as the means ± SD (OD 492 nm)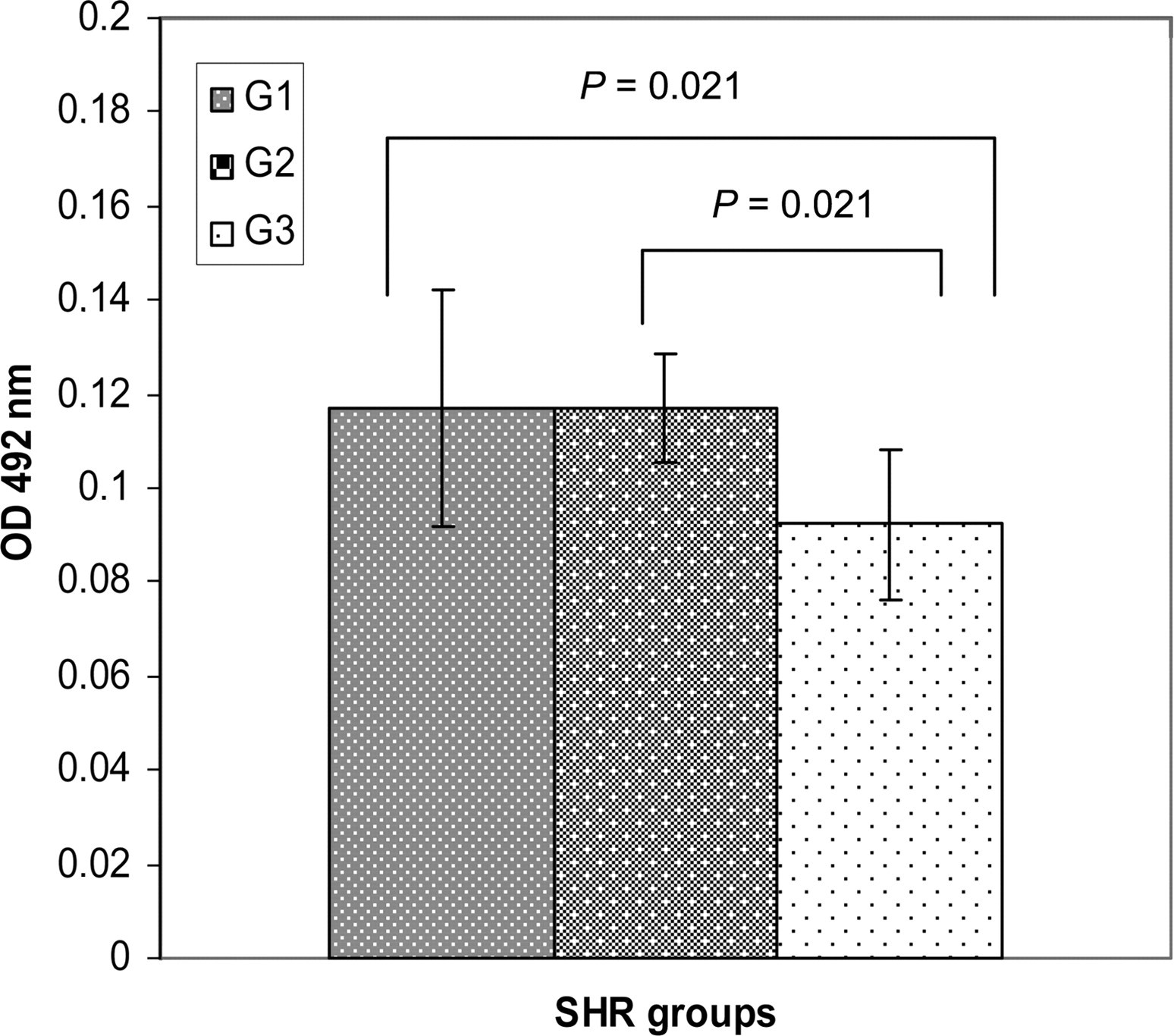
These data were supported by the histological and EM analysis and the morphometry of aortic walls of the experimental animals.
The histology findings showed the degenerative changes at different stages of development in each of the groups, and highlighted the difference in aortic wall engagements – from endothelial hyperplasia and proliferation of myocytes to fibrosis and hialinosis.
The rats from G1 showed severe degenerative changes, and many atherosclerotic spots spread throughout the thoracic to the abdominal aorta. In the thoracic aorta, the plaques with a cellular proliferation predominated. The plaques with fibrosis and hialinosis predominated in the abdominal aorta, and were located near the branches of the arteries. They had confluenced, deforming the lumen of the aorta and causing stenosis. Deep endothelial erosions, hyperplasia of myocytes, fibrosclerotic plaques with multiple necroses and lipid drops in the media were found in the aortas of rats from G1. The whole wall of the vessel, from the endothelium to adventitia, was found to be involved. The internal elastic membrane was thick and pushed out by growing collagen that caused edema and separation between the intima and media of the wall. Degeneration and vacuolization of smooth muscle cells were seen. They were functionally transformed to fibrocytes. This process led to thickening and loss of elasticity in the vessel wall (Figure 3).
A representative photomicrograph of the aortic wall of rats from G1. HE: Aorta – deep endothelial erosions (thin arrow), hyperplasia of myocytes, fibrosclerotic plaque with multiple necroses and lipid drops in media (thick arrow). The whole wall of the vessel, from the endothelium to adventitia, is engaged (magnification ×20). EM: Aorta – endothelial cells with deformed nuclei and lipid drops, below them (thin arrow), high thickness of the elastic membrane and powerful growing collagen (thick arrow) (magnification ×4000). HE, hemalaun–eosin; EM, electron microscopy. (A color version of this figure is available in the online journal)
All rats from G2 had endothelial hyperplasia, and light erosions. Myocyte proliferation, single necrobiotic changes and small lipid drops in the media were seen. EM also revealed a moderate thickness of elastic membrane, hyperplasia of smooth muscle cells with altered integrity and optical empty spaces (Figure 4).
A representative photomicrograph of the aortic wall of rats from G2. HE: Aorta – proliferation of endothelial cells and light erosions (thin arrow), single necrobiotic changes and small lipid drops in media (thick arrow) (magnification ×10). EM: Aorta (part of media) – moderate thickness of elastic membrane (thin arrow), hyperplasia of smooth muscle cells with altered integrity and optical empty spaces (thick arrow) (magnification ×6000). HE, hemalaun–eosin; EM, electron microscopy. (A color version of this figure is available in the online journal)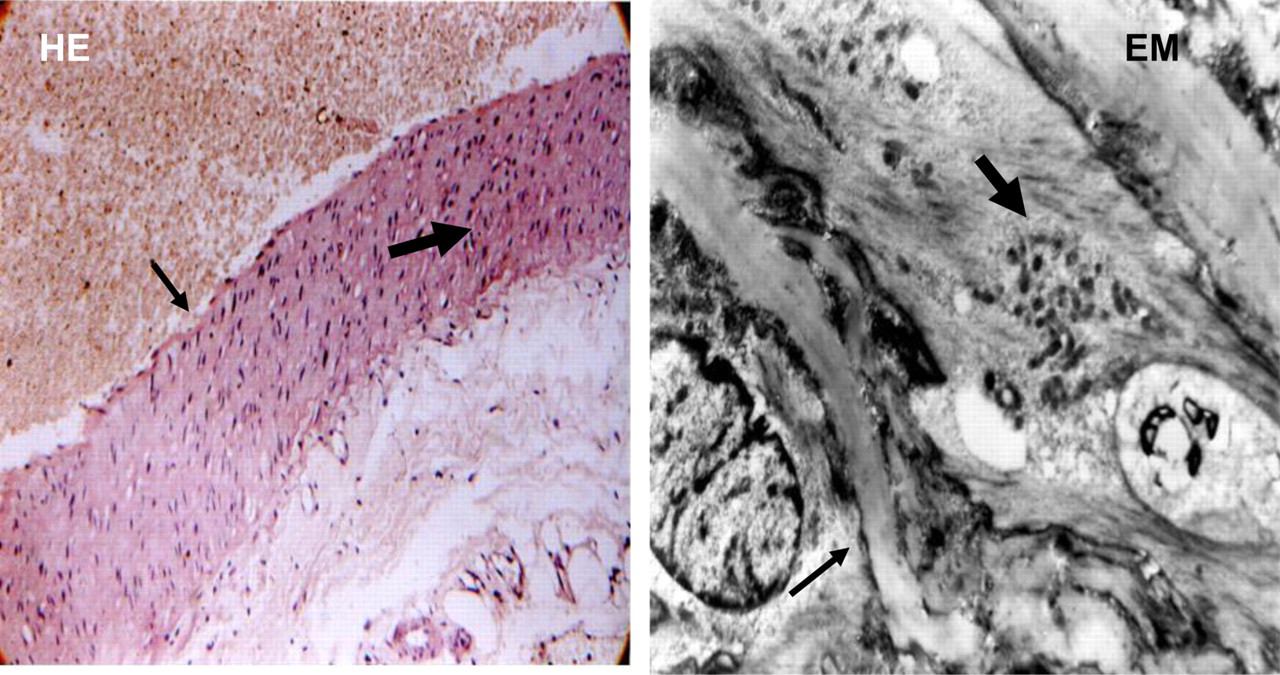
The histology of the aortic walls showed a small number of degenerative changes in G3, which evolved slowly over time. A uniform thickness of the aortic wall, moderate endothelial proliferation and erosions, light myocyte hyperplasia and small lipid drops in the media were seen. EM analysis supported the light microscope findings, revealing a normal ultrastructure of the smooth muscle cells, and a slow rate of production of young collagen in SHR of G3. This distinguished G3 from the others, in which mature collagen, lipid drops and an altered integrity of the media predominated (Figure 5).
A representative photomicrograph of the aortic wall of rats from G3. HE: Aorta – uniform thickness of aortic wall, moderate endothelial proliferation with light erosions (thin arrow), light hyperplasia of myocytes and small lipid drops in media (magnification ×10). EM: Aorta (part of media) – young smooth muscle cells with normal ultrastructure (thin arrow), light increase of elastic membrane thickness and scanty quantity of collagen (thick arrow) – (magnification ×7000). (A color version of this figure is available in the online journal)
No difference was found in the thoracic aortic wall thickness between the groups. However, there was a statistically significant difference in the thickness of the abdominal aortic walls. The data as the means ± SD in microns are: for thoracic aorta – G1 = 15.11 ± 0.92, G2 = 14.95 ± 1.13 and G3 = 14.63 ± 0.80 (P > 0.05) and for abdominal aorta – G1 = 14.69 ± 0.79, G2 = 13.45 ± 0.49 and G3 = 13.42 ± 0.50 (P = 0.0008). These results are presented graphically in Figure 6.
Thickness of thoracic and abdominal aorta walls (microns) of rats in G1, G2 and G3 as the means (micron)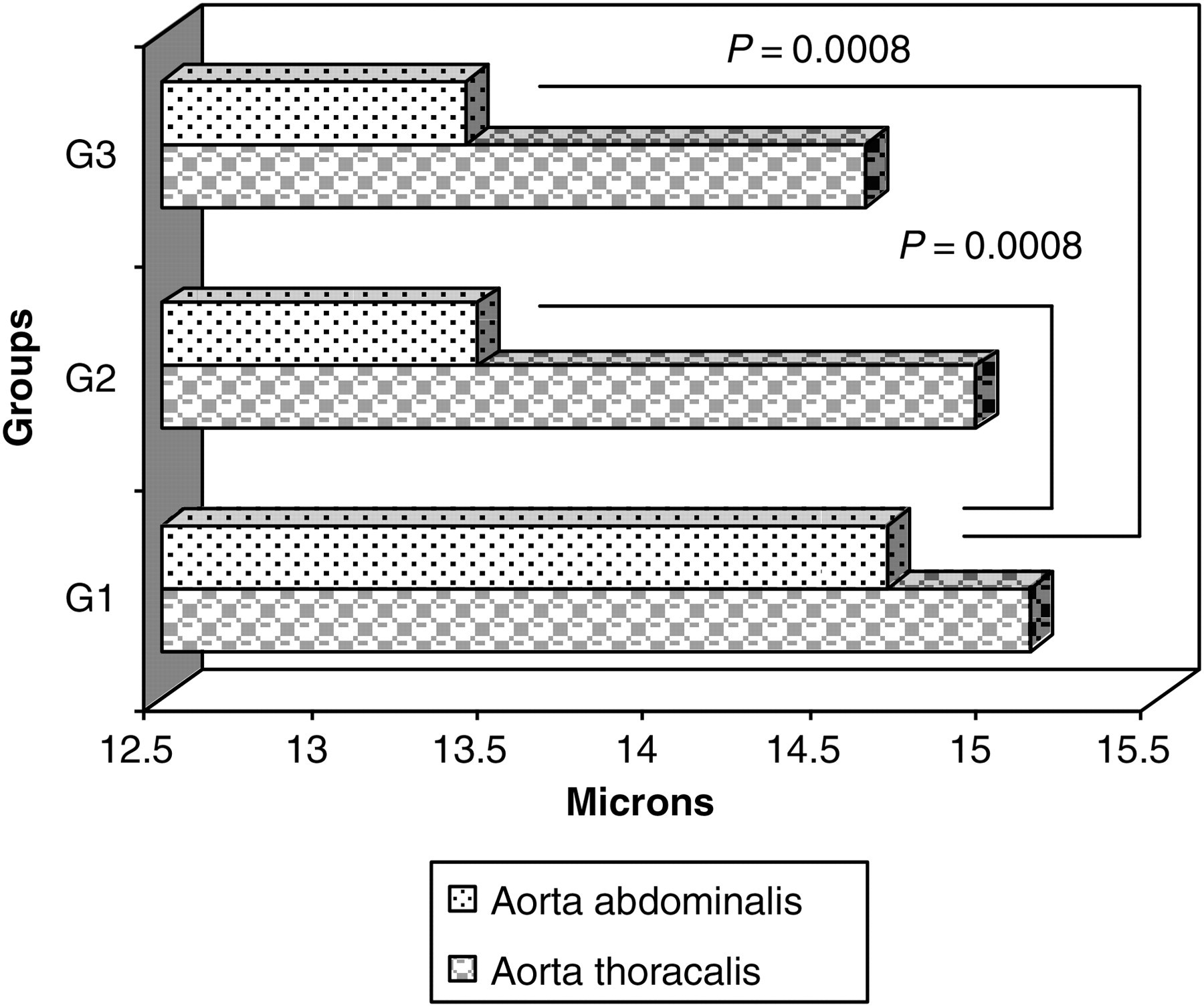
The morphometry of the left coronary artery showed reduced thickness in G3, compared with the other groups (P = 0.005). The results for the thickness of the left coronary artery walls as the means ± SD in microns were: G1 = 3.22 ± 0.38; G2 = 3.32 ± 0.31; and G3 = 2.72 ± 0.34 (Figure 7).
Thickness of vessel wall of the left coronary artery of rats in G1, G2 and G3 as the means (micron)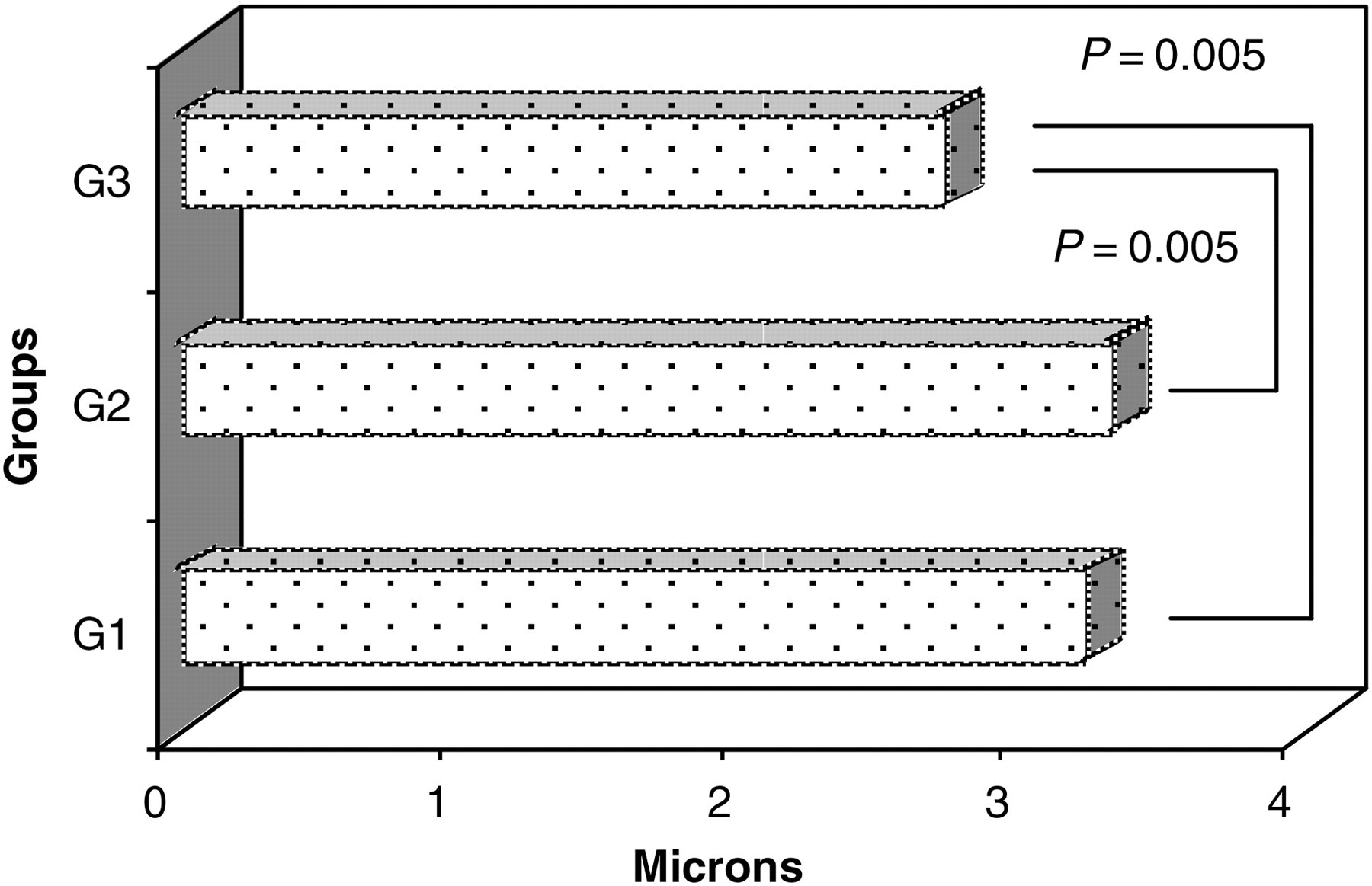
A relatively week positive relationship between the serum concentration of AEABs and thickness of wall of the left coronary artery was described (r = +0.395; R 2 = 15.60%; P = 0.056).
In conclusion, the rats with Se supplementation have a higher GPx-1 activity and lower level of AEABs; a lower degree of development and spread of the degenerative changes to the aortic wall; and a presence of young smooth muscle cells, a low quantity of collagen, and a thinner elastic membrane and thinner vessel walls than the rats from the other groups. Rats that received a low-Se diet showed severe degenerative changes and increased thickness of the examined vessel walls, a high rate of collagen production and thickened elastic membranes – events that act to increase the stiffness of the vessel walls, change the lumen and may lead to the chronically ischemic disturbances of the organ's blood supply.
Discussion
Selenoproteins play an important biological role in mammals, acting to mediate optimal endocrine and immune function, and participating in the prevention of cardiovascular and neoplastic diseases. They are grouped into three families of enzymes: GPx, thioredoxin reductases and iodothyronine 5I deiodinases. Six isoenzymes of GPx that participate in antioxidative defense of mammalian organisms have been evaluated. The thioreductases control cellular redox systems. Three iodothyronine deiodinases have been identified, which catalyze a release of iodine from the 5 or 5I position of the iodothyronin substrates and play an important role for activation and inactivation of the thyroid hormones in all tissues. 1
The cellular antioxidant enzymes such as GPx-1 and superoxide dismutase play a major role in the control of ROS, which participate in the atherogenesis and pathogenesis of hypertension. GPx catalyze reduction of hydrogen peroxide (H2O2) and organic peroxides. GPx-1 plays the most important role for elimination of peroxides from the cells. H2O2 modulates the different aspects of the function of endothelial cells: their growth and proliferation; apoptosis; endothelium-dependent vasodilation; barrier function; and endothelial inflammatory answer and endothelial control of vessel re-modeling. 13,14 Using animal models, it was evaluated that the abnormal redox status of the aortic wall was of primary importance for the development of arterial hypertension and atherosclerosis and had a determining effect on the disturbed control of vasomotor tone. 15 Endothelial dysfunction precedes the development of lesions in the vessels. The presence of chronically increased blood pressure increases the oxidative stress of arteries through the activation of the protein kinase-C-dependent NAD(P)H-oxidase pathway and through the activation of the local renin–angiotensin system. 16
GPx-1 deficit, which is due to low Se intake, directly induces an increase in oxidative stress, leading to an endothelial dysfunction. 17 The determined GPx-1 activity of whole blood in this experiment was only at a normal level for the rats in G3 (559 ± 78 U/g Hb). Se supplementation increased GPx-1 activity by 18%, whereas low Se intake did not significantly decrease it. It was established that for the Wistar rats under a diet of Se content 0.1 μg Se/g of food, the normal level of GPx-1 activity of whole blood was 528 ± 20 U/g Hb and it was varied in the different organs. GPx-1 expression was the highest in the liver, erythrocytes, lungs and heart. 18 Se supplementation increased the synthesis of selenoproteins because of the increased level of Sec-tRNA, but low Se intake drastically decreased the levels of GPx-1 and selenoprotein P. 19 Our data showed that the recommended Se intake was not enough to ensure normal GPx-1 activity in SHR at this age of life.
The low GPx-1 activity, increasing oxidative stress, decreases bioavailable nitric oxide (NO), which is synthesized by the endothelium by two different mechanisms. The first one is the uncoupling of endothelial nitric oxide synthase, because of oxidation of its co-factor tetrahydrobiopterin and thus decreasing the NO formation and increasing the O2 − production. The inactivation of NO occurs also by a reaction of NO− radical with some other ROS. Data have previously been published reporting that up-regulation of arginase by H2O2 impairs endothelium-dependent NO-mediated dilation of the coronary arterioles. 20 NO contributes to vascular tone, preserves endothelial integrity, and inhibits smooth muscle cell migration and proliferation. Endothelial dysfunction causes an imbalance in the synthesis of the vasodilatator NO and the vasoconstrictor endothelin-1, resulting in increased resistance in the vessels. 5,17,21
Elastin is the major component of the vessel wall, is responsible for the elasticity of the big vessels and is synthesized and split continuously. Elastin peptides derived from its degradation are present in the circulating blood and they stimulate increased production of antibodies against these modified self-structures. AEABs are physiologically connected with breakdown products of elastin metabolism and enter circulation through the monocyte–macrophage system. Significant changes in elastin metabolism play a role in alterations to the integrity of the elastin-rich organs (vessels, lung, skin, muscles). Relatively high levels of AEABs (class IgG) were established in the serum of children, which reach even higher values in the serum of 18–20-year-old individuals. Their levels then stabilize in the serum of 30–60-year-old persons and gradually decrease thereafter. An increase in serum concentrations of AEABs indicates the presence of changed structures in the vessel wall. 7–10,12
Previous studies in SHR have shown that aortic collagen and elastin biosynthesis follow a biphasic pattern. Both collagen and elastin synthesis have been found to exceed Wistar–Kyoto rat control levels in the prehypertensive period (at the age of 4 weeks). They then decrease in the development of hypertension (to the age of 14–16 weeks), and then increase again in the period of the established hypertensive state (beyond the age of 16 weeks). Several studies showed that this second increase in connective tissue proteins may be prevented, depending on the choice of antihypertensive therapy. 22,23
SHR show early degenerative changes to the vessels, because it is evident that hypertension is the risk factor for development of atherosclerosis. Excessive production of collagen, low elastin production and high elastin degradation are considered as causes for increased arterial rigidity. Data have been published showing that endothelial dysfunction is a significant factor for the increase of arterial rigidity in patients with isolated systolic hypertension and during senescence. 24
The turnover rate for collagen and elastin is low in healthy arteries, but vascular pathology upsets the regulatory pathways that maintain this balance. The effects of hypertension on elastic fibers are fatigue and accelerate degradation, leading to loss of arterial wall resilience. Except these well-known effects, the individual components of elastic fibers and their degradation products can act as signal transducers and regulate cellular proliferation, migration, phenotype and extracellular matrix (ECM) protein degradation and in this way are associated with hypertension progression. In response to hypertension, the over-expression of both proinflammatory and proteinase-inhibitory molecules dramatically increases arterial ECM protein synthesis. At the same time, elastin and elastic fibers are progressively degraded by enzymatic processes, involving an imbalance between antiproteases and proteases – elastin degrading enzymes, i.e. elastases, including several matrix metalloproteases (MMP), such as MMP-2 and MMP-9. 25,26
Our results support these findings. The rat's genetic brand used had a long period of established hypertension and altered elastic structures. The serum concentration of AEABs was significantly lower in rats with Se supplementation than the other groups. We speculate that improved redox status may be the cause for poor alterations of the vessel wall and decreased activity of proteinases.
EM analysis of the aortic wall revealed a normal ultrastructure in the smooth muscle cells, and a slow rate of young collagen production in SHR of G3. In the rats from G1 and G2, the internal elastic membranes were thicker and damaged by the growing collagen.
Morphological data collected by our experiment showed that alterations in the vessel walls began from the thoracic aorta to the abdominal aorta and propagated to the arteries. Selenium supplementation could not repair the already existed changes, but could slow down their propagation along the arterial tree.
It is known that growth, apoptosis, inflammation and fibrosis contribute to vascular re-modeling in hypertension. 27 Our findings showed that low GPx-1 activity affects normal functions of the endothelial cells, thus altering control of the vasomotor tone, causing proliferation of smooth muscle cells, changes to elastic membranes, accumulation of collagen and fibrosis. As a consequence, the endothelium becomes rough, and the media thickens and encroaches to the lumen. These changes may be in a position to cause turbulent blood flow, susceptibility to atherothrombosis, low perfusion and ischemic disturbances in the organs.
Hypertension-induced target organ damage (TOD) is one of the leading causes of morbidity and mortality. Regression of arterial hypertrophy depends on the specific antihypertensive treatment. The activity of the antioxidant defense system of the organism is lowered with the age. A low Se intake worsens the antioxidant status and is a risk factor for the development of cardiovascular diseases (CVD), whereas Se supplementation has a positive effect in reducing TOD. Our results have revealed important evidence in support of this hypothesis, because the therapeutic benefit of Se administration in the prevention and treatment of CVD still remains insufficiently documented. 28 Cross-sectional and longitudinal population studies have demonstrated a negative relation between systolic blood pressure and serum Se. In addition to antihypertensive drug therapy, enhancement of the endogenous defense against ROS by a supranutritional level of Se is more safe and effective than supplementation of other antioxidants for the reduction of hypertensive TOD, because the protective effects of GPx are of particular importance when the organism is exposed to additional stress factors. 6,29,30
The data presented here show that selenium supplementation, reducing oxidative stress in SHR, slowed down elastin degradation and development of degenerative changes of the vessel wall. We suggest that it will be important in the future to undertake more specific clinical investigations to determine the efficacy of Se supplementation in the treatment of CVD, complementing existing drug therapies.
Footnotes
Acknowledgements
This study was supported by Medical University, Pleven, Bulgaria. We thank Professor Roumen G Roussev for helpful discussion and for the improvement of the English presentation.