
Abstract
This study was designed to investigate the hepatoprotective effects of magnesium chenoursodeoxycholic acid (Mg-CUD), a magnesium trihydrate salt of ursodeoxycholic acid and chenodeoxycholic acid, in carbon tetrachloride (CCl4)-induced liver fibrosis in rats. Rats were treated with CCl4 dissolved in olive oil (0.5 mL/kg, twice a week) intraperitoneally for eight weeks. Mg-CUD was administered orally at 15.625, 31.25, 62.5 and 125 mg/kg once a day. Chronic CCl4 administration induced increases in serum transforming growth factor-β1, hepatic hydroxyproline content and serum alanine aminotransferase activity. Mg-CUD attenuated these increases. The levels of α-smooth muscle actin protein and mRNA expression were increased by chronic CCl4 exposure and Mg-CUD attenuated these increases. Mg-CUD suppressed increases in matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-1 mRNA expression and elevation of oxidative stresses by attenuating lipid peroxidation and enhancing reduced glutathione/oxidized glutathione ratio. The overexpression of toll-like receptor 4 and increased nuclear translocation of nuclear factor-κB and phosphorylated c-Jun, a component of activator protein 1, were suppressed by Mg-CUD. Furthermore, CCl4 increased the levels of tumor necrosis factor (TNF)-α, interleukin (IL)-6, IL-10 and cyclooxygenase (COX)-2. Mg-CUD attenuated the levels of TNF-α, IL-6 and COX-2, while it augmented the level of IL-10. Our results suggest that Mg-CUD may prevent liver fibrosis by modulating collagen accumulation and inflammatory signaling pathways.
Introduction
Liver fibrosis, resulting from the wound-healing response to repeated injury, represents a major medical problem with high morbidity. The molecular mechanism of liver fibrosis has been under continuous investigation, and carbon tetrachloride (CCl4) has been used extensively as a xenobiotic for induction of liver fibrosis similar to that in humans. While administration of a single dose of CCl4 can result in centrizonal necrosis and steatosis, prolonged administration can result in liver fibrosis, cirrhosis and hepatocellular carcinoma. CCl4 is converted to a trichloromethyl radical (CCl3•) by cytochrome P450 2E1, resulting in lipid peroxidation and membrane damage followed by inflammatory responses in Kupffer cells. 1
As the primary source of fibrillar and non-fibrillar matrix protein, hepatic stellate cells (HSCs) are central to the process of fibrosis. 2 Migration and accumulation of activated HSCs occurs at sites of tissue repair, regulating synthesis of matrix protein and matrix degradation by matrix metalloproteinase (MMP) and tissue inhibitor of metalloproteinase (TIMP). 3 The pathogenesis of liver fibrosis is associated with a complicated network of liver, non-parenchymal cells, and balance between matrix production and degradation. 4
Liver fibrosis is highly linked to chronic hepatocellular damage and subsequent inflammatory response that produces inflammatory cytokines and recruits inflammatory leukocytes into the injured site. Clinically, all patients with liver fibrosis have symptoms of inflammation, which shows correlation with progression of fibrosis. The inflammatory environment stimulates the activation of HSCs through various mediators such as transforming growth factor (TGF)-β and platelet-derived growth factor. 5
Toll-like receptors (TLRs) are essential to innate immune defenses and after activation by foreign pathogens or host-derived molecules, TLR signal cascades induce inflammatory response. 6 As a receptor for lipopolysaccharide (LPS), TLR4 activates nuclear factor-κB (NF-κB); ultimately, a large number of proinflammatory mediators are synthesized and released. 7 According to a recent report by Seki et al., TLR4 plays a key role in progression of hepatic fibrogenesis. In addition, TLR4 was reported to enhance activation of HSCs triggered by TGF-β1, thereby implicating a novel molecular association of hepatic inflammation with fibrogenesis. 8
Patients with cholesterol-rich gallstones have been treated with ursodeoxycholic acid (UDCA) and chenodeoxycholic acid (CDCA), which are considered to be effective therapeutic agents. UDCA was reported to induce a delay in progression of liver fibrosis in primary biliary cirrhosis and its efficacy was also observed in a model of bile duct ligation-induced fibrosis.
9
Alternative therapies, such as combination treatment of UDCA with pegylated interferon α2b or retinoic acid, attenuated liver injury in an experimental fibrosis model.
10,11
Efficacy in dissolving of gallstones by CDCA was reported to be superior to that of UDCA,
12
and 6-ethyl CDCA, a semi-synthetic bile acid, was found to promote resolution of liver fibrosis by inhibiting TIMP-1 expression resulting from activation of farnesoid X receptor (FXR).
13
However, due to the various adverse effects of CDCA, a combination therapy of UDCA and CDCA, aiming for higher dissolution efficacy with minimal side-effects, has been suggested. Findings from our recent study demonstrated amelioration of
Therefore, this study was conducted to determine the efficacy of Mg-CUD for use in treatment of CCl4-induced liver fibrosis and to investigate its molecular mechanisms.
Materials and methods
Chemicals
Mg-CUD (C&U cap.®) was obtained from Myungmoon Pharm. Co, Ltd (Seoul, Korea). The composition of Mg-CUD is as follows: 45.6% UDCA, 45.6% CDCA and 8.8% magnesium. All chemicals used in the experiments were obtained from Sigma Chemical Co (St Louis, MO, USA), if not designated otherwise.
Animals and treatment regimen
Male Sprague-Dawley rats were supplied by Orient Bio Inc (Seongnam, Korea); weights of the animals ranged from 100 to 120 g. All animal procedures received approval from the Sungkyunkwan University Animal Care Committee and were conducted in agreement with the guidelines of the National Institutes of Health. Animals received intraperitoneal administration of CCl4, which was dissolved in olive oil (1:3, v/v), twice per week (0.5 mL/kg) for a period of eight weeks. During this period, animals received once-per-day oral administration of Mg-CUD suspended in 0.5% methylcellulose (vehicle). Animals were assigned in a random manner to six different groups, as follows: (a) vehicle-treated control; (b) vehicle-treated CCl4 (CCl4); and (c–f) Mg-CUD (15.625, 31.25, 62.5 and 125 mg/kg)-treated CCl4 groups. Rats were sacrificed at 24 h after the final treatment with CCl4 and blood (from inferior vena cava) and liver tissue were collected. Liver tissue was analyzed immediately for histological staining (sections from the left lobe) and the remaining portions of the liver tissues were frozen immediately in liquid nitrogen and kept at −75°C until biochemical analyses.
Hepatic hydroxyproline content
The method of Jamall et al. 15 was used for measurement of hydroxyproline content in the liver. Following homogenization in 6 N hydrochloride (HCl), liver tissues underwent hydrolysis at 110°C for a period of 18 h. After cooling of the hydrolysate, chloramine T was added. Five minutes later, p-dimethylaminobenzaldehyde was added to the mixture, followed by incubation for a period of 30 min at 60°C. Using a reagent blank containing the complete system without added tissue, samples were read on a spectrophotometer at 560 nm (Hitachi, Tokyo, Japan).
Serum alanine aminotransferase activity
Serum alanine aminotransferase (ALT) activity was determined by spectrophotometric procedures, using the ChemiLab ALT assay kit (IVDLab Co, Uiwang, Korea).
Histological analysis
For histological analysis, sections were made of the anterior portion of the left lateral lobe of the liver. For fixation, the tissues were immersed in 10% neutral-buffered formalin, followed by paraffin embedding. Hematoxylin-eosin (H&E) staining was performed on sections of approximately 5-μm thickness. Histological changes were evaluated in a blind manner using an Olympus CKX 41 microscope (Olympus Optical Co, Ltd, Tokyo, Japan).
Level of hepatic lipid peroxidation and reduced glutathione/oxidized glutathione ratio
Using the method described by Buege and Aust, the steady-state level of malondialdehyde (MDA), the end product of lipid peroxidation, in liver homogenates was analyzed by spectrophotometric measurement of the level of substances that were reactive to thiobarbituric acid at 535 nm. 16 1,1,3,3-Tetraethoxypropane was used as the standard. Using the method described by Tietze, 17 the total level of glutathione was measured spectrophotometrically at a wavelength of 412 nm using yeast glutathione reductase, 5,5′-dithio-bis(2-nitrobenzoic acid) and nicotinamide adenine dinucleotide phosphate (NADPH). Using the same method, oxidized glutathione (GSSG) concentration was measured in the presence of 2-vinylpyridine. The difference between total levels of glutathione and GSSG was used for determination of the level of reduced glutathione (GSH).
Serum tumor necrosis factor-α, interleukin-6, interleukin-10 and TGF-β1 concentrations
Serum tumor necrosis factor-α (TNF-α), interleukin (IL)-6, IL-10 and TGF-β1 concentrations were individually detected using commercially available TNF-α, IL-6, IL-10 and TGF-β1 enzyme-linked immunosorbent assay (ELISA) kits (BD Biosciences, San Diego, CA, USA) according to the manufacturer's protocols.
Western blot
The procedure for Western blotting was based on a previous study, 18 which utilized 15 μg of whole protein for measurement of the content of α-smooth muscle actin (SMA), TLR4 and cyclooxygenase-2 (COX-2) and 15 μg of nuclear protein for measurement of the content of the NF-κB p65 subunit and phosphorylated-c-Jun (p-c-Jun). Densitometric evaluation of visualized immunoreactive bands was performed using ImageQuant™ TL software (Amersham Biosciences/GE Healthcare, Piscataway, NJ, USA). Primary antibodies used were as follows: α-SMA (Abcam, Cambridge, MA, USA; 1:1,000), TLR4 (Santa Cruz Biotechnology, Santa Cruz, CA, USA; 1:1,000), COX-2 (Cayman, Ann Arbor, MI, USA; 1:500), NF-κB/p65 (Santa Cruz Biotechnology; 1:1000) and c-Jun p39 phosphorylated on serine-63 (Santa Cruz Biotechnology; 1:500); signals were normalized to β-actin (Sigma Chemical Co; 1:1000) or lamin B1 (Abcam; 1:2500), respectively.
Reverse transcription polymerase chain reaction
Reverse transcription polymerase chain reaction (RT-PCR) was performed using liver tissues. Using a Takara RNA PCR kit (AMV) Version 3.0 (Takara Bio Inc, Shiga, Japan), reverse transcription was performed for extraction of total RNA and synthesis of the first strand of cDNA. For the PCR reaction, a 20-μL reaction volume was used with a diluted cDNA sample. The final concentrations for the reaction were as follows: sense and antisense primers, 0.5 μmol/L; dNTP mix, 0.25 mmol/L; 10× Ex Taq buffer, 1 μL; and Ex Taq (Takara Bio Inc), 0.25 U/reaction. The performance of PCR included an initial denaturation step at 94°C for five minutes, and a final extension step at 72°C for seven minutes using the GeneAmp 2700 thermocycler (Applied Biosystems, Foster City, CA, USA). The gene-specific primers used are shown in Supplementary Table 1 (please see
Statistical analysis
Results were reported as mean ± SEM. The overall significance of the results was determined by one-way analysis of variance using the SPSS Version 12.0 statistical software package (SPSS, Chicago, IL, USA). For determination of differences between groups, P < 0.05 was considered significant, and an appropriate Bonferroni correction was used for multiple comparisons.
Results
Serum TGF-β1 concentration
In the control group, the serum concentration of TGF-β1 was 764.7 ± 76.4 pg/mL. As a result of treatment with CCl4, the level of serum TGF-β1 showed a significant increase, to 5589.3 ± 457.0 pg/mL, which was attenuated by 125 mg/kg of Mg-CUD (1203.5 ± 72.5 pg/mL, P < 0.01) (Figure 1).
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on serum transforming growth factor (TGF)-β1 concentration and hepatic hydroxyproline content. (a) The serum concentration of TGF-β1 was determined using an enzyme-linked immunosorbent assay. (b) Hepatic hydroxyproline content was measured as described in ‘Materials and methods’. Results are presented as mean ± SEM of 8–10 animals per group. **denotes significant differences (P < 0.01) compared with the control group; ++ denotes significant differences (P < 0.01) compared with the CCl4 group
Hepatic hydroxyproline content
Figure 1 shows the level of hydroxyproline in liver tissues. The level of hydroxyproline in the control group was 207.8 ± 34.0 μg/g liver. Compared with the control group, Mg-CUD (125 mg/kg) alone had no effect on the level of hydroxyproline (data not shown). However, in the CCl4 group, hydroxyproline content showed a marked increase, to 740.3 ± 71.9 μg/g liver, and 125 mg/kg of Mg-CUD was found to attenuate this increase (549.9 ± 7.1 μg/g liver, P < 0.01).
Serum ALT activity
As shown in Figure 2, the level of serum ALT activity was 34.5 ± 3.5 U/L in the control group. After administration of CCl4 for eight weeks, serum ALT activity showed a significant increase, to 437.7 ± 34.9 U/L. Compared with the control group, Mg-CUD (15.625, 31.25, 62.5 and 125 mg/kg) alone had no effect on serum ALT activity (data not shown). Increase in ALT activity was found to be significantly attenuated by 62.5 and 125 mg/kg of Mg-CUD (291.1 ± 21.6 U/L, P < 0.01 and 235.8 ± 23.2 U/L, P < 0.01, respectively).
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on serum alanine aminotransferase (ALT) activity. Rats received intraperitoneal injection of carbon tetrachloride (CCl4) (0.5 mL/kg) twice per week for a period of eight weeks. During this period, Mg-CUD or vehicle was administered orally once per day. Serum ALT activity was measured spectrophotometrically. Results are presented as mean ± SEM of 8–10 animals per group. **denotes significant differences (P < 0.01) compared with the control group; ++denotes significant differences (P < 0.01) compared with the CCl4 group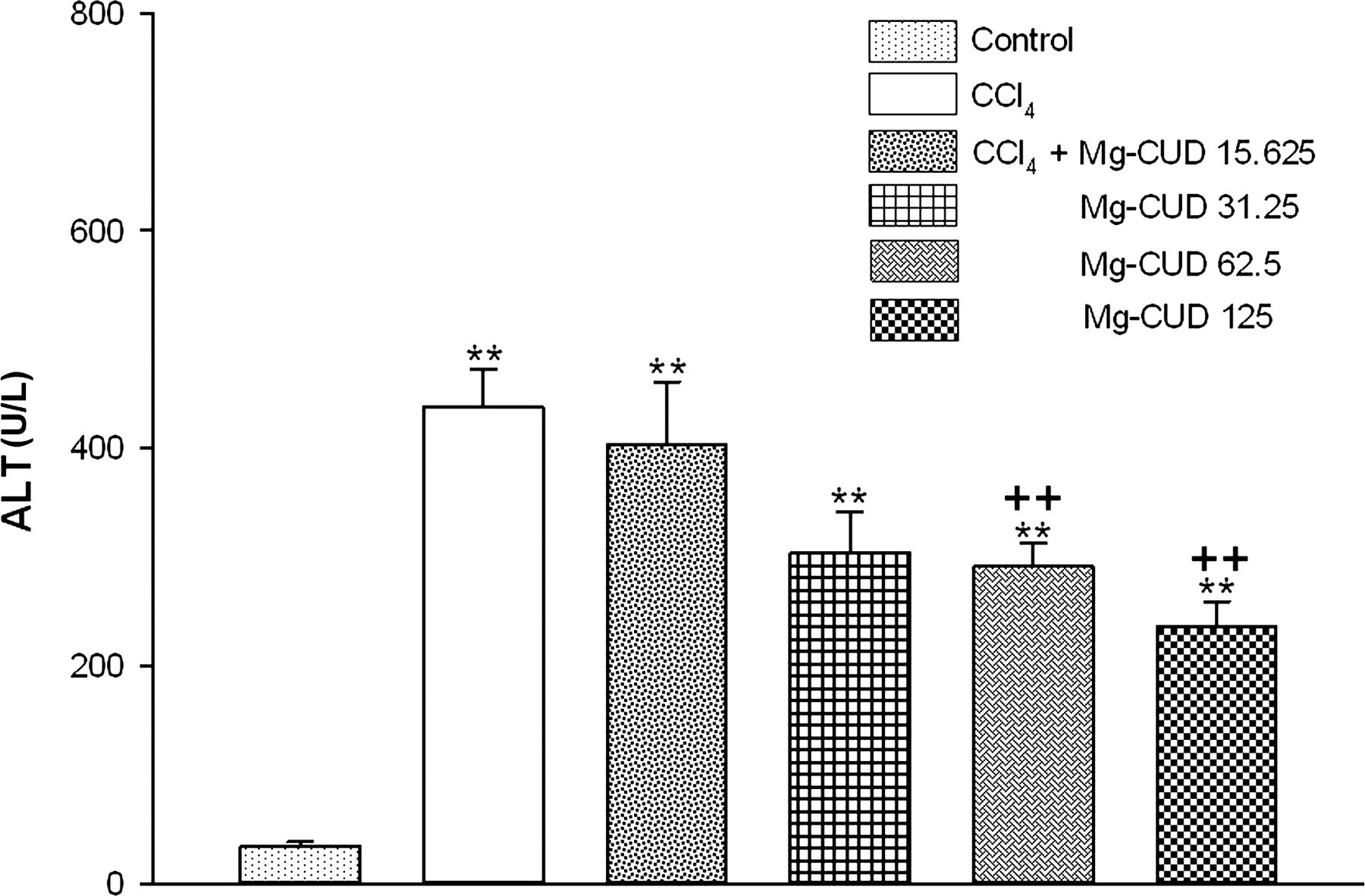
Pathological observation
Extensive accumulation of fat droplets, hepatocellular necrosis and inflammatory cell infiltration in the CCl4 group were observed by H&E staining. These pathological changes were ameliorated by Mg-CUD (Figure 3).
Histological features of liver sections stained with hematoxylin and eosin. Representative photomicrographs are shown (original magnification × 200). Arrows indicate fat droplets, hepatocellular necrosis and inflammatory cell infiltration. (a) Control, normal hepatic architecture, (b) chronic CCl4 treatment, necrotic area and a large amount of inflammatory cell infiltration surrounding the centrilobular veins of the liver, (c) CCl4 + Mg-CUD 125 mg/kg, with no significant necrosis. Mg-CUD, magnesium chenoursodeoxycholic acid; CCl4, carbon tetrachloride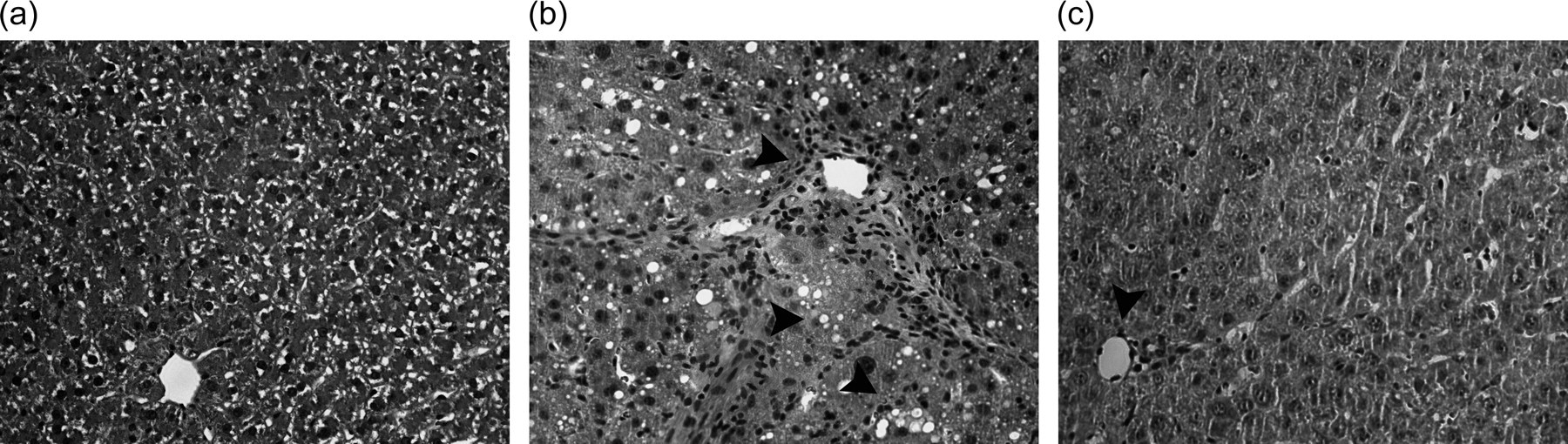
α-SMA protein and mRNA expression
Figure 4 shows changes in the levels of α-SMA protein and mRNA expression. After repeated injections of CCl4, levels of α-SMA protein and mRNA expression showed a significant increase, to 1.6- and 3.1-times the control value, respectively. Mg-CUD at a concentration of 125 mg/kg was found to significantly attenuate these increases (69.2% of CCl4 group, P < 0.01 and 66.1% of CCl4 group, P < 0.01, respectively).
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on protein and mRNA expression of α-smooth muscle actin (SMA). Results are presented as mean ± SEM of 8–10 animals per group. (a) Western blot analysis for α-SMA was performed on whole extracts from liver; (b) α-SMA mRNA expression in liver was measured by reverse transcription polymerase chain reaction analysis. **denotes significant differences (P < 0.01) compared with the control group; ++denotes significant differences (P < 0.01) compared with the CCl4 group
MMP-2 and TIMP-1 mRNA expression
Compared with the control group, levels of expression of MMP-2 and TIMP-1 mRNA were 2.0- and 3.2-fold higher in the CCl4 group, respectively. These increases were significantly suppressed by 125 mg/kg of Mg-CUD (46.4% of CCl4 group, P < 0.01 and 52.7% of CCl4 group, P < 0.01, respectively) (Figure 5).
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on matrix metalloproteinase (MMP-2) and tissue inhibitor of metalloproteinase (TIMP-1) mRNA expression. Results are presented as mean ± SEM of 8–10 animals per group. MMP-2 and TIMP-1 mRNA expression in liver were measured by reverse transcription polymerase chain reaction analysis 24 h after carbon tetrachloride (CCl4) injection. **denotes significant differences (P < 0.01) compared with the control group; ++ denotes significant differences (P < 0.01) compared with the CCl4 group
Hepatic lipid peroxidation concentration and GSH/GSSG ratio
Effects of Mg-CUD on oxidative stress
MDA, malondialdehyde; GSH, reduced glutathione; GSSG, oxidized glutathione; Mg-CUD, magnesium chenoursodeoxycholic acid
Rats received intraperitoneal injection of CCl4 (0.5 mL/kg) twice a week for eight weeks. During the period, Mg-CUD (125 mg/kg) or vehicle was administered orally once a day. The results are presented as mean ± SEM of 8–10 animals per group
**Significant differences (P < 0.01) compared with control group
+,++Significant differences (P < 0.05, P < 0.01) compared with CCl4 group
TLR4 protein expression and nuclear translocation of NF-κB and activator protein 1
TLR4 protein expression in the CCl4 group showed a significant increase, to 1.8-fold, compared with the control group. This increase was attenuated by Mg-CUD (68.4% of CCl4 group, P < 0.01). Nuclear levels of p65, a subunit of NF-κB, and phosphorylated c-Jun (p-c-Jun) showed a marked increase to 2.2- and 2.0-fold in the CCl4 group, compared with the control group, respectively. Mg-CUD at a concentration of 125 mg/kg was found to attenuate these increases (71.3% of CCl4 group, P < 0.01 and 70.3% of CCl4 group, P < 0.05, respectively) (Figure 6).
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on toll-like receptor 4 (TLR4) protein expression (a) and nuclear translocation of nuclear factor (NF)-κB and activator protein-1 (AP-1) (b). Results are presented as mean ± SEM of 8–10 animals per group. Western blot analysis for TLR4 and NF-κB and p-c-Jun was performed on whole and nuclear extracts from liver, respectively. *, **denote significant differences (P < 0.05, P < 0.01) compared with the control group; +, ++denote significant differences (P < 0.05, P < 0.01) compared with the CCl4 group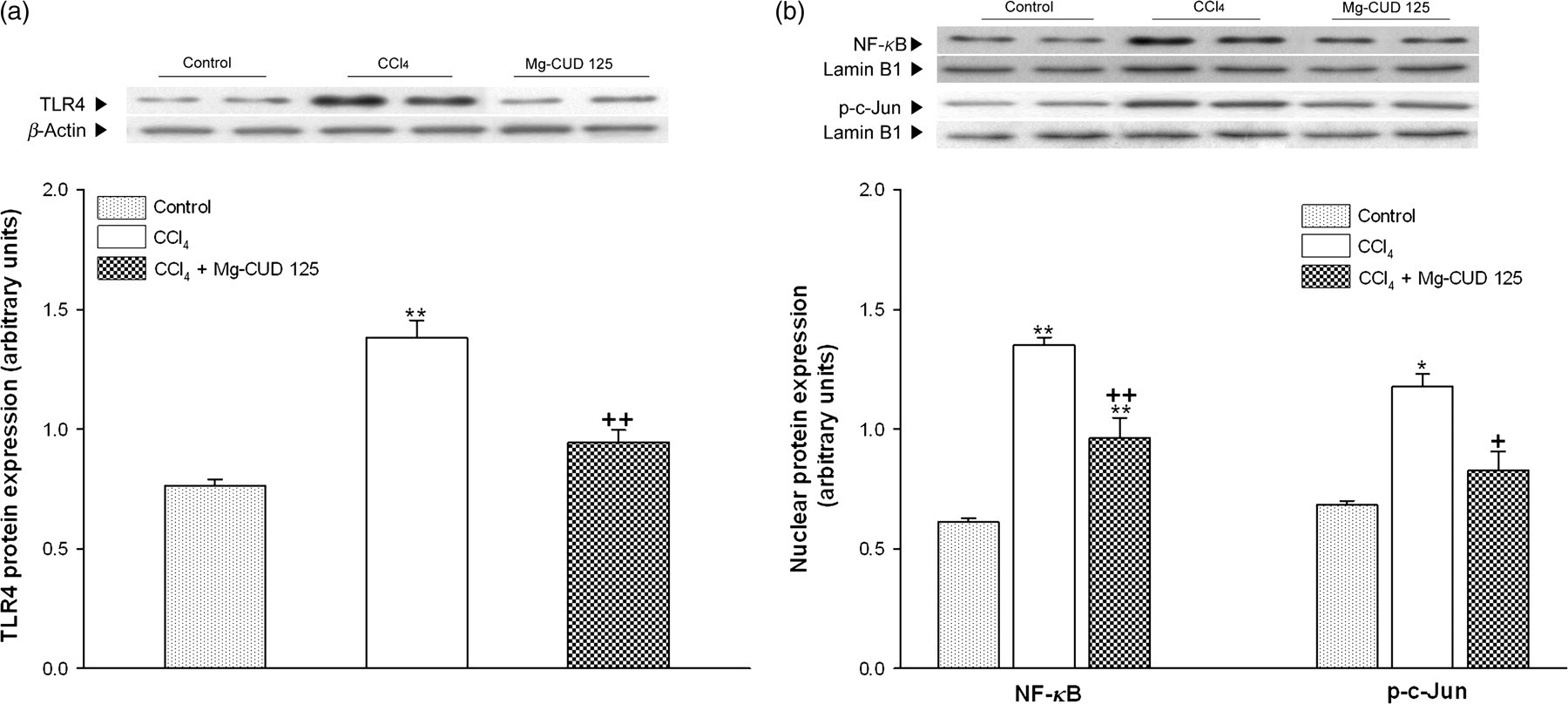
Serum concentrations and mRNA expression of TNF-α, IL-6 and IL-10
Effects of Mg-CUD on serum inflammatory cytokines
TNF, tumor necrosis factor; IL, interleukin; Mg-CUD, magnesium chenoursodeoxycholic acid
Rats received intraperitoneal injection of CCl4 (0.5 mL/kg) twice a week for eight weeks. During the period, Mg-CUD (125 mg/kg) or vehicle was administered orally once a day. The serum concentration of TNF-α, IL-6 and IL-10 was determined using enzyme-linked immunosorbent assay. The results are presented as mean ± SEM of 8–10 animals per group
*,**Significant differences (P < 0.05, P < 0.01) compared with control group
+,++Significant differences (P < 0.05, P < 0.01) compared with CCl4 group
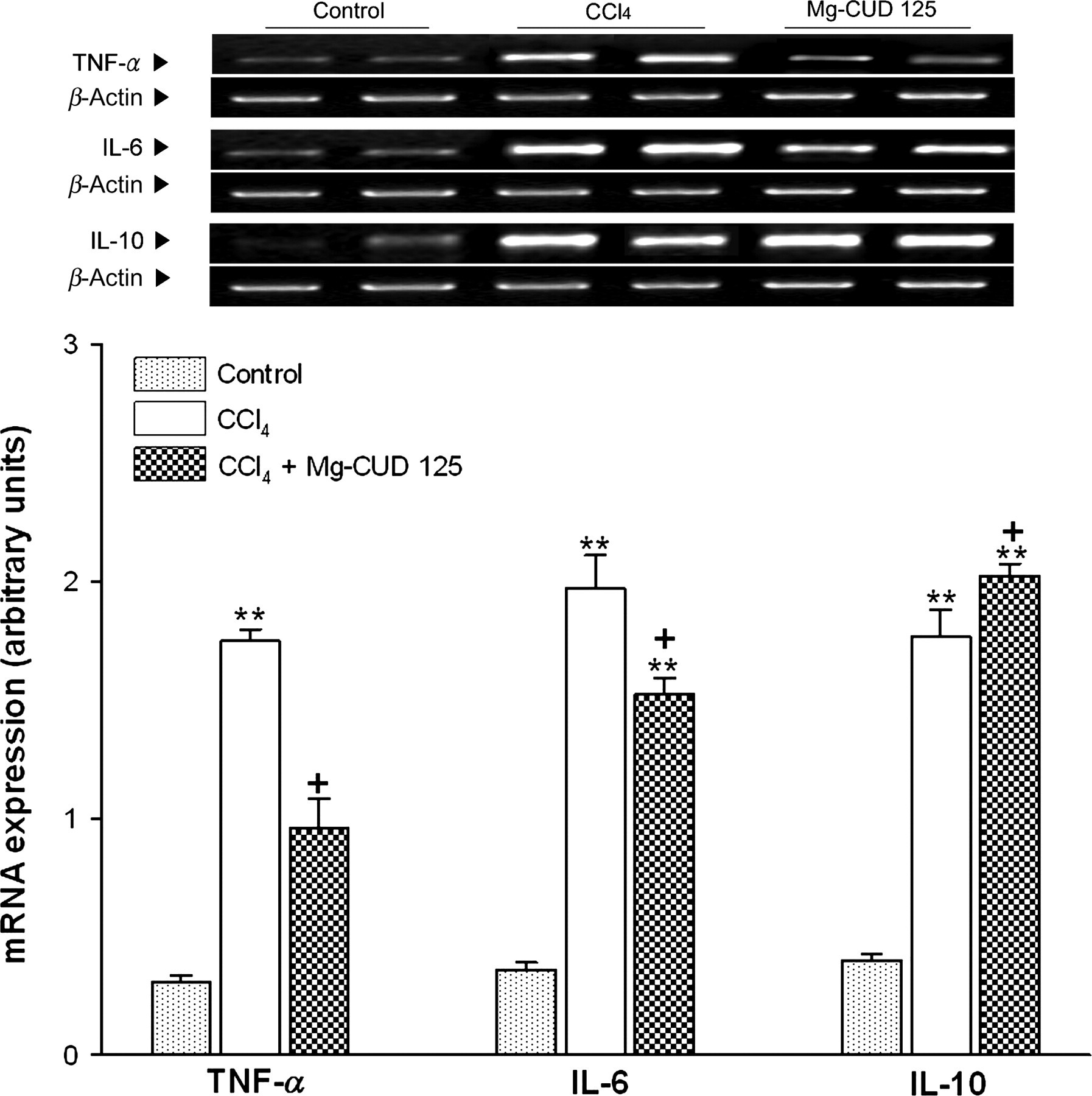
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on mRNA expression of inflammatory cytokines. Results are presented as mean ± SEM of 8–10 animals per group. Tumor necrosis factor (TNF)-α, interleukin (IL)-6 and IL-10 mRNA expression in liver were measured by reverse transcription polymerase chain reaction analysis. **denotes significant differences (P < 0.01) compared with the control group; +denotes significant differences (P < 0.05) compared with the CCl4 group
COX-2 protein and mRNA expression
Figure 8 shows changes in the levels of COX-2 protein and mRNA expression. After repeated injections of CCl4, levels of COX-2 protein and mRNA expression showed a significant increase to 2.1- and 9.2-times the control value, respectively. Mg-CUD at a concentration of 125 mg/kg was found to significantly attenuate these increases (63.5% of CCl4 group, P < 0.01 and 57.5% of CCl4 group, P < 0.01, respectively).
Effects of magnesium chenoursodeoxycholic acid (Mg-CUD) on protein and mRNA expression of cyclooxygenase 2 (COX-2). Results are presented as mean ± SEM of 8–10 animals per group. (a) Western blot analysis for COX-2 was performed on whole extracts from liver; (b) COX-2 mRNA expression in liver was measured by reverse transcription polymerase chain reaction analysis. **denotes significant differences (P < 0.01) compared with the control group; ++denotes significant differences (P < 0.01) compared with the CCl4 group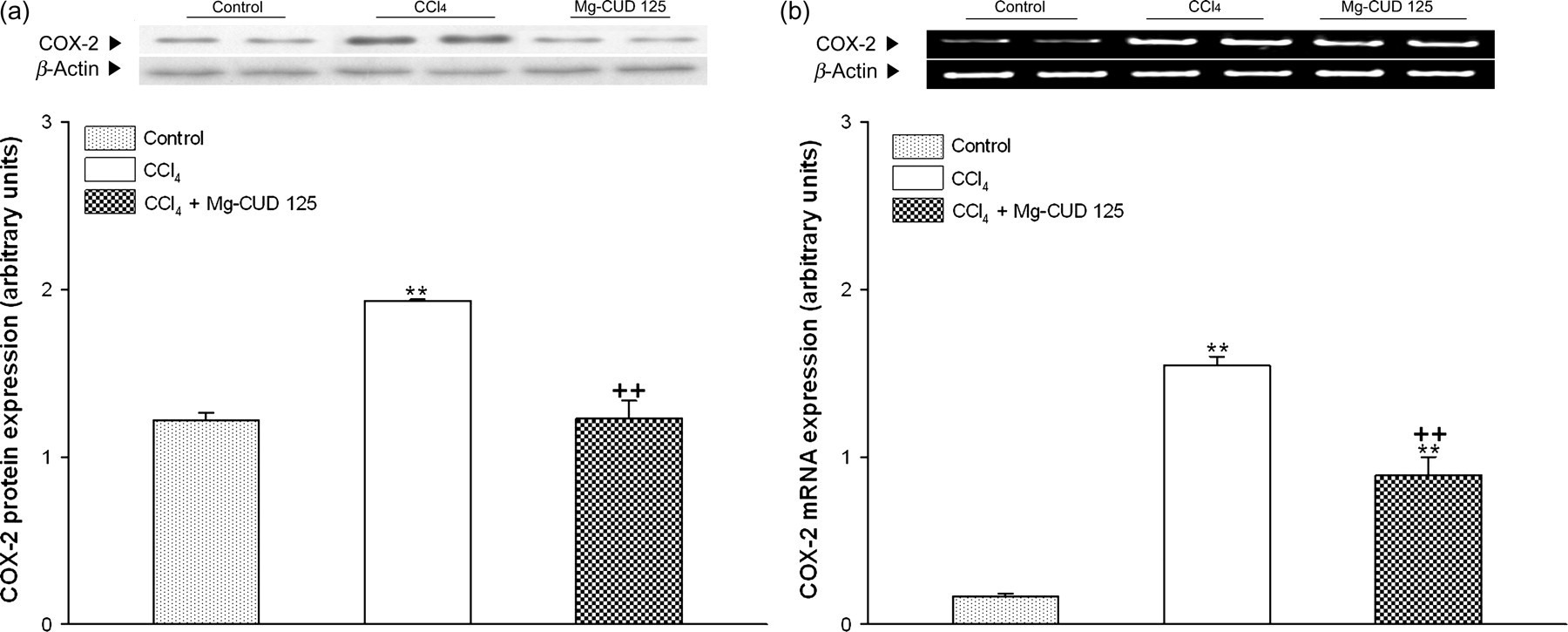
Discussion
Liver fibrosis results from excessive accumulation of extracellular matrix (ECM) in response to chronic injury. Improvement of hepatocyte ultrastructure by UDCA in liver fibrosis has been demonstrated. 19 UDCA decreased intrahepatic resistance, which mostly acted through the inhibition of hepatic thromboxane A2 production and lipid peroxidation and led to the prevention of hepatic fibrosis and cirrhosis. 20 UDCA also induced a reversal in development of hepatic fibrosis through its effect on the signaling pathway of TGF-β1/Smad by inhibiting expression of TGF-β1, Smad3 and cAMP response element (CREB) binding protein and increasing expression of Smad7. 21 A study with CDCA, the most active natural ligand of FXR, provided evidence that in vivo activation of FXR induces a decrease in expression of TIMP-1, as well as an amelioration of liver fibrosis. 13 FXR is expressed by HSCs, and FXR ligands function as negative regulators of α1(I)-collagen synthesis both in vitro and in vivo in rodent models of hepatic fibrosis. 22,23
In respect of clinical use, Podda et al. concluded that a combination treatment of equimolar doses of UDCA and CDCA can be recommended for treatment of gallstones since it has greater effects on bile cholesterol saturation than either alone, is better tolerated than CDCA (less diarrhea and hypertransaminasemia), and is less expensive than UDCA.
24
These advantages could be applied for the treatment of hepatic fibrosis requiring long-term use of drugs to avoid development of cirrhosis. In our previous studies, Mg-CUD outperformed its half dose of UDCA or CDCA in α-naphthylisothiocyanate-induced cholestasis and ethionine-induced fatty liver rat models (data not shown). In addition, we recently reported that Mg-CUD was found to protect the liver from
TGF-β is one of the significant fibrogenic cytokines involved in activation of HSCs from a state of quiescence to formation of myofibroblasts, the most significant source of collagen-producing cells in liver injury. Collagens are constructed from hydroxyproline residues; therefore, ultimate accumulation of ECM can be detected by hydroxyproline content in hepatic tissue, a marker for liver fibrosis. 26 We confirmed induction of liver fibrosis in rats by an increase in serum TGF-β1 concentratrion and hepatic hydroxyproline content after repeated CCl4 exposure, and this was suppressed by Mg-CUD. We assessed histological lesions and all of the animals were found to possess fibrotic features even though the variations of hepatic damage severity existed among the animals in each group. Extensive areas of fatty change and gross necrosis, significant infiltration of lymphocytes, Kupffer cell hyperplasia and fibrous septa were observed by H&E staining after repeated CCl4 exposure. Mg-CUD 125 mg/kg was shown to be most effective to alleviate the histological damages induced by CCl4. Serum ALT activity, a representative marker of liver function, also increased after chronic CCl4 administration. Increase in ALT activity by repeated CCl4 injection was found to be significantly attenuated by 62.5 and 125 mg/kg of Mg-CUD while 15.625 and 31.25 mg/kg of Mg-CUD did not affect the serum ALT activity. Therefore, Mg-CUD at 125 mg/kg was selected as the optimal effective dose for evaluating the molecular mechanisms of Mg-CUD against CCl4-induced hepatotoxicity in this study.
During liver fibrosis, the quiescent HSCs become activated with the excess production of α1(I)-collagen and α-SMA. While losing their typical star-shape, they acquire a myofibroblast-like phenotype such as proliferative, motile, profibrogenic and contractile functions. 27 Thus, changes in the amounts of α-SMA, a fibroblastic cell hallmark in liver tissues, were evaluated. Mg-CUD was found to attenuate an increase in the levels of expression of α-SMA protein and mRNA after chronic exposure to CCl4, suggesting Mg-CUD suppressed the activation of HSC.
Activated HSCs express the essential elements required for matrix degradation; they are also an important source of MMPs, a family of zinc-dependent enzymes, which include collagenases, gelatinases and stromelysins. 28 Changes in MMP expression and activity result in a re-structuring of the hepatic ECM during liver injury, leading to both direct and indirect acceleration of further HSC activation. These MMPs are regulated primarily by TIMPs (TIMPs 1–4), which demonstrate substrate- and tissue-specific binding to MMPs, resulting in blockade of their proteolytic activity. 29 Levels of TIMP mRNA and protein show a dramatic increase during fibrosis, so that levels of MMP either show a modest increase or remain essentially unchanged. 30 However, during fibrogenesis, the level of MMP-2, a 72-kDa type IV collagenase, shows an exceptional increase. MMP-2 degrades normal subendothelial ECM, accelerating its replacement by fibril-forming collagen, which stimulates HSC growth and MMP-2 itself in a positive feedback loop. 31 Thus, the net effect of a rapid increase in ECM synthesis, in combination with increased TIMP activity, accentuates formation of the fibrotic scar. In our study, treatment with Mg-CUD was found to attenuate an increase in the levels of expression of MMP-2 and TIMP-1 mRNA following chronic administration of CCl4, which suggests inhibition of collagen synthesis and enhanced fibrolytic activity by Mg-CUD through decreasing levels of MMP-2 and TIMP-1.
Oxidative stress has been observed in the majority of chronic liver diseases, and different etiologies and progressions of fibrosis in rats have been associated with a decrease in antioxidant defenses. 32 Release of reactive oxygen species (ROS) that stimulate HSC proliferation and transformation into α-SMA-positive myofibroblast-like cells by hepatocytes undergoing oxidative stress has been reported. 33 Glutathione and glutathione-related enzymes play detoxifying and antioxidative roles through conjugation with glutathione or free-radical reduction in living tissue; these components are easily inactivated by lipid peroxides or ROS during CCl4 exposure. 34 In this study, Mg-CUD was found to impair CCl4-mediated oxidative stress as a result of decreased production of free-radical derivatives, which was demonstrated by the decrease in MDA concentration. In addition, Mg-CUD was found to suppress the decrease in hepatic GSH/GSSG ratio after repeated injections of CCl4. Increase in the hepatic GSH/GSSG ratio, which was observed in Mg-CUD-treated rats, might have occurred as a result of its effect on de novo synthesis of glutathione, its regeneration or both. Consequently, maintenance of the hepatic GSH/GSSG ratio at levels sufficient for neutralization of the increase in formation of free radicals, as in the case of CCl4 toxicity, could be achieved. These findings demonstrate the possibility that antioxidant properties may be a mechanism of alleviation of liver damage induced by chronic CCl4 exposure by Mg-CUD.
Growing evidence has demonstrated a close association between inflammation and chronic hepatic injury, as well as the consequent process of wound-healing. 8 Recent studies in both experimental models and humans have implicated TLRs as a pivotal regulatory system of inflammatory cascades; it has also been found to mediate cross-talk occurring between inflammation and fibrosis in association with chronic liver injury. 35 Expression of TLR4 on Kupffer cells, hepatocytes, HSCs and dendritic cells for maintenance of homeostasis of the hepatic environment is known to occur in the liver. 36 TLR4 signaling is pivotal to activate the HSCs. Intestinal microflora has been regarded as an important source of LPS, a potent ligand of TLR4 in liver fibrosis. TLR4 signaling drives recruitment of inflammatory cells and induces down-regulation of TGF-β pseudoreceptor BMP and the activin membrane-bound inhibitor (Bambi) for fibrogenic response. 8 TLR4 signaling triggers activation of NF-κB and JNK/AP-1 (activator protein 1) pathways through two adapter molecules, myeloid differentiation factor 88 and toll-receptor-associated activator of protein of interferon; 37 modulation of TGF-β signaling by a TLR4-NF-κB axis affords a novel target for proinflammatory and profibrogenic signals. 8 AP-1, an inducible transcription factor, has also been associated with diverse cellular processes, including cell proliferation, differentiation, apoptosis and neoplastic transformation. 38 Upon activation, AP-1 binds to a palindromic sequence, inducing activation of downstream target genes, including type I collagen and TGF-β1, which may result in deposition of ECM. 39 Our study showed that TLR4 protein expression increased following chronic CCl4 exposure; Mg-CUD was found to attenuate this increase. Mg-CUD was found to suppress the increase in nuclear expression of NF-κB and p-c-Jun. In addition, levels of proinflammatory mediators, including TNF-α and IL-6, were increased by repeated administration of CCl4; levels of TNF-α and IL-6 were found to be attenuated by Mg-CUD. Collectively, the protective effect of Mg-CUD is partially the result of suppression of the TLR4-mediated inflammatory signaling pathway.
Synthesis of IL-10, a potent anti-inflammatory cytokine involved in inhibition of synthesis of proinflammatory cytokines by T helper type 1 cells, mono/macrophages and neutrophils, can be performed by several cell types within the liver; these include Kupffer cells and hepatocytes. Modulation of inflammatory response and limitation of hepatotoxicity by IL-10 have been demonstrated in several models of liver injury. 40 According to previous reports, IL-10 exerts antifibrogenic effects through down-regulation of profibrogenic cytokines, including TGF-β1 and TNF-α; exogenous IL-10 can suppress mRNA expression of MMP-2 and TIMP-1 in vivo. 41 After eight weeks of CCl4 administration, an increase in the serum concentration of IL-10 and its mRNA expression was observed, and Mg-CUD was found to augment these increases, implying that the antifibrotic and anti-inflammatory effects of Mg-CUD are possibly the result of its up-regulation of IL-10.
COX-2 is induced primarily by various types of inflammatory stimuli, including LPS, TNF-α and IL-1β. 42 COX-2 is associated with chronic liver diseases, such as hepatitis C infection, and has been found to be up-regulated in association with necro-inflammatory injury in experimental models. 43,44 Enhancement of the synthetic level of TGF-β and its receptors both in vitro and in vivo by prostaglandin E2, a product of COX-2 activity, has been demonstrated. 45 Chavez et al. 46 reported on celecoxib, a selective COX-2 inhibitor, which was found to exert antifibrotic and fibrolytic properties against liver damage induced by CCl4. Mg-CUD was found to attenuate an increase in the levels of expression of COX-2 protein and mRNA after chronic exposure to CCl4, which translates to occurrence of an antifibrotic effect via inhibition of COX-2 at the transcriptional level.
In conclusion, Mg-CUD demonstrated a protective effect against CCl4-induced fibrosis, which could be attributed to its inhibition of collagen deposition and down-regulation of inflammatory response through the TLR4 signaling cascade. Findings from this study demonstrate the potential for development of Mg-CUD as a novel therapeutic agent for use in prevention of liver fibrosis.
Footnotes
ACKNOWLEDGEMENTS
J-WK has received ‘Global PhD Fellowship Program’ support from the National Research Foundation of Korea (2011-0006724).