
Abstract
Mesenchymal stem cells (MSCs) have significant immunomodulatory effects in the development of acute lung inflammation and fibrosis. However, it is still unclear as to whether MSCs could attenuate allergic airway inflammation in a mouse model of asthma. We firstly investigated whether exogenous MSCs can relocate to lung tissues in asthmatic mice and analyzed the chemotactic mechanism. Then, we evaluated the in vivo immunomodulatory effect of exogenous MSCs in asthma. MSCs (2 × 106) were administered through the tail vein to mice one day before the first airway challenge. Migration of MSCs was evaluated by flow cytometry. The immunomodulatory effect of MSCs was evaluated by cell counting in bronchoalveolar lavage fluid (BALF), histology, mast cell degranulation, airway hyperreactivity and cytokine profile in BALF. Exogenous MSCs can migrate to sites of inflammation in asthmatic mice through a stromal cell-derived factor-1α/CXCR4-dependent mechanism. MSCs can protect mice against a range of allergic airway inflammatory pathologies, including the infiltration of inflammatory cells, mast cell degranulation and airway hyperreactivity partly via shifting to a T-helper 1 (Th1) from a Th2 immune response to allergens. So, immunotherapy based on MSCs may be a feasible, efficient therapy for asthma.
Introduction
Asthma is an immunological disease that has increased dramatically in prevalence over the past two decades. Exaggerated T-helper 2 (Th2)-biased immune responses result in the development of asthma. 1 Immunotherapies that can regulate an allergen-specific Th2 immune response may provide long-lasting control of asthma. Recently, there have been an increasing number of studies demonstrating a therapeutic role of mesenchymal stem cells (MSCs) in rodent models of acute lung inflammation and fibrosis, which suggest that MSCs have significant immunomodulatory effects in the lung. 2 Indeed, MSCs can inhibit the proliferation and function of a broad range of immune cells, including T-cells, B-cells, natural killer cells and dendritic cells. 3 Whether MSCs can attenuate Th2-predominant allergic airway inflammation is a challenging hypothesis.
The immunoregulatory effects of MSCs are mainly mediated through direct cell-to-cell interactions and paracrine effects. 3 So, the recruitment of MSCs to the sites of inflammation and injury is very important. Chemokine and chemokine receptor are the major regulators of the trafficking of MSCs. SDF-1 (stromal cell-derived factor-1α), acting via its receptor, CXCR4, is a chemoattractant for MSCs. 4 Several studies have shown that the SDF-1-CXCR4 axis was implicated in the recruitment of MSCs to the liver and pancreatic islets. 5,6 Whether SDF-1-CXCR4 is also involved in the migration of MSCs to the lungs in an animal model of asthma is still undetermined.
The aim of this study was to observe whether exogenous MSCs can re-locate to lung tissues in asthmatic mice and suppress allergic airway inflammation, and further investigate the chemotactic and immunoregulatory mechanism of exogenous MSCs.
Materials and methods
Animals
Healthy female C57BL/6 mice (6–8 weeks of age) were purchased from the experimental animal center of Fourth Military Medical University. All mice were bred under pathogen-free conditions. All mouse protocols were approved by the Animal Experiment Administration Committee of Fourth Military Medical University.
Allergen sensitization and challenge protocol
Mice were sensitized intraperitoneally with 100 μg ovalbumin (OVA; A5503, Sigma-Aldrich, St Louis, MO, USA) adsorbed to 9% aluminum hydroxide hydrate (A1577, Sigma-Aldrich) on days 1 and 8. Then, mice were challenged with 100 μg OVA in phosphate-buffered saline (PBS) by an intratracheal route on days 15, 16 and 17. Control mice were treated with PBS.
Culture, labeling and transfer of MSCs
A frozen vial of murine bone marrow MSCs (isolated from female C57Bl/6J mice; Cyagen Biosciences, Guangzhou, China) was thawed and expanded according to the supplier's instructions. MSCs used in all experiments were between passages 3 and 4. MSCs were labeled with 1 μmol/L 5(6)-(N-succinimidyloxycarbonyl)-3′, 6′, 0, 0′-diacetylfluorescein (CFSE) on day 12. On day 14, labeled MSCs (2 × 106 cells in 0.1 mL PBS) were administered through the tail vein to mice. Control mice were treated with PBS. Some plates of MSCs were treated with AMD3100 (a CXCR4 antagonist, 200 ng/mL, Sigma-Aldrich) 24 h before cell transfer. 4 MSCs were washed to remove AMD3100 prior to cell transfer.
Analysis of whole-lung cells by flow cytometry
On day 18, mice were sacrificed. Lungs were removed and used to obtain single-cell suspensions for flow-cytometric analysis. 7 Cells were labeled with anti-MAC-1-PE (557397; BD Biosciences, San Jose, CA, USA), anti-CXCR4-PE (551966; BD Biosciences), anti-CD44-PE (553134; BD Biosciences), anti-CD45-PE (553081; BD Biosciences), anti-CD90-PE (551401; BD Biosciences) or anti-Sca-1-PE (553108; BD Biosciences) and then analyzed using a flow cytometer (BD Calibur; BD Biosciences). The frequency of each cell subset was calculated.
Bronchoalveolar lavage fluid
Mice were sacrificed on day 18. Bronchoalveolar lavage fluid (BALF) was obtained by the slow injection of ice-cold saline (0.3 mL) into the trachea three times (total, 0.9 mL). Total cell number counting and differential counting in BALF, concentration of interleukin-5 (IL-5), IL-9, IL-4 and interferon-γ (IFN-γ) and β-hexosaminidase activity in BALF were performed as previously described. 8
Histology
Lung consecutive sections (5 μm) were stained with hematoxylin and eosin and randomly numbered. The degree of peribronchial and perivascular inflammation was evaluated by a pathologist blinded to the random numbers on a subjective scale of 0, 1, 2, 3 and 4 corresponding to none, mild, moderate, marked or severe inflammation, respectively. The lung inflammation scores were defined as the sum of peribronchial and perivascular inflammation scores.
Measurement of airway responsiveness
Airway responsiveness was assessed as described previously. 9 Briefly, mice were anesthetized, intubated and ventilated (120 breaths/min, 0.2 mL tidal volume). Then, mice were paralyzed with decamethonium and injected intravenously with 50 μg/kg acetylcholine. Airway responsiveness is expressed as the airway pressure–time index (APTI).
Reverse transcriptase-polymerase chain reaction analysis
Lung tissues were collected for detecting the expression of SDF-1 mRNA. The primers 5′ CACTTTCACTCTCGG TCCAC and 5′ CTGAAGGGCACAGTTTGGAG for SDF-1, and 5′ AACCCTAAGGCCAACCGTGAAAAG and 5′ GCAGGATGGCGTGAGGGAGAG for β-actin control were used.
Quantitative reverse transcriptase polymerase chain reaction
Quantitative reverse transcriptase polymerase chain reaction (RT-PCR) was performed as we previously described, 8 and the same primers for β-actin, IL-4 and IFN-γ were used.
Western blotting
Lung tissues were collected for detecting the expression of SDF-1 protein with anti-SDF-1 polyclonal antibody (sc-6193; Santa Cruz Biotechnology, Santa Cruz, CA, USA). Anti-β-actin polyclonal antibody (sc-1616; Santa Cruz) was used as control.
Statistical analysis
Data were expressed as mean ± SEM. Statistical analyses were performed using single-factor analysis of variance on the three groups and with a Student's unpaired t-test for comparisons of two groups. A P value of <0.05 was assumed to be significant.
Results
Migration of exogenous MSCs to lungs
To investigate whether exogenous MSCs can re-locate to lung tissues in asthmatic mice, we followed the fate of transferred MSCs bearing the CFSE marker, which permits transferred MSC identification in recipient mice. Mice were sensitized twice with OVA on days 1 and 8. Then, airway challenge was performed on days 15, 16 and 17 after MSC transfer on day 14. The frequency of CFSE+ cells in whole-lung cells was analyzed by flow cytometry on day 18.
In PBS-challenged mice, the relative ratio and the absolute number of CFSE+ cells significantly increased after MSC transfer (Figure 1a, P < 0.05, P < 0.01 respectively), which indicated that administration of MSCs through the tail vein was feasible. When labeled MSCs were transferred into OVA-induced asthmatic mice, more CFSE+ cells were found in lung tissues (Figure 1a, P < 0.05, P < 0.01). Further analysis revealed that the phenotype of these CFSE+ cells was CD44−CD45−CD90+Sca-1+, which indicated that these CFSE+ cells should be CFSE-carrying MSCs (please see online Supplemental Figure at

Migration of exogenous MSCs to lungs. (a) The relative ratio and the absolute number of CFSE+ cells in whole-lung cells. Mice were sensitized twice with OVA or PBS on days 1 and 8. Labeled MSC transfer was performed on day 14. On days 15, 16 and 17, airway challenge was performed. Whole-lung cells were analyzed by flow cytometry on day 18. Labels indicate sensitization/treatment/challenge. P, M, O denotes PBS, MSCs, OVA, respectively. Representative flow cytometric analysis and percentages of cells in indicated squares are shown. Absolute cell numbers were calculated for total whole-lung cells. Horizontal bars denote paired experimental groups for which statistical significance is displayed in the figure. Data are the mean ± SD from three experiments (n = 15). (b) Expression of MAC-1 in CFSE+ cells. In vitro group: MSCs labeled with CFSE were cultured in vitro for four days and then expression of MAC-1 was analyzed. In vivo group: MSC labeled with CFSE were transferred to OVA-sensitized mice and then mice were challenged with OVA for three days. Four days later after MSC transfer, whole-lung cells was analyzed. Representative flow cytometric analysis is shown. Data are the mean ± SD from three experiments (n = 15). MSC, mesenchymal stem cell; PBS, phosphate-buffered saline; CFSE, 5(6)-(N-succinimidyloxycarbonyl)-3′,6′,0,0′-diacetylfluorescein; OVA, ovalbumin
Lung phagocytes, including macrophages, neutrophils and monocytes, etc., can phagocytize dead CFSE+ MSCs and be presented with CFSE+ phenotype. To exclude the influence of CFSE+ phagocytes, we observed the expression of Mac-1 both in cultured MSCs in vitro and in lung CFSE+ cells from MSC-transferred asthmatic mice. As shown in Figure 1b, no significant difference was observed in the relative ratio of Mac-1+ CFSE+ cells in total CFSE+ cells (Figure 1b, P > 0.05). Thus, exogenous MSCs can migrate to the sites of inflammation in asthmatic mice.
Role of the CXCR4/SDF-1 axis in the migration of exogenous MSCs
Chemokine receptors expressed on MSCs mediated their migration to tissues. Reportedly, CXCR4 was implicated in the recruitment of MSCs to the liver and pancreatic islets. 5,6 To observe whether CXCR4 is also involved in the migration of exogenous MSCs to the lungs, we detected the change of CXCR4 expression on the cell surface of MSCs. The relative ratio of CXCR4+ cells in total CFSE+ cells significantly increased in the lungs of MSC-transferred asthmatic mice compared with cultured MSCs in vitro (Figure 2a, P < 0.01). Thus, the migration of exogenous MSCs may be related with the up-regulation of CXCR4 expression.
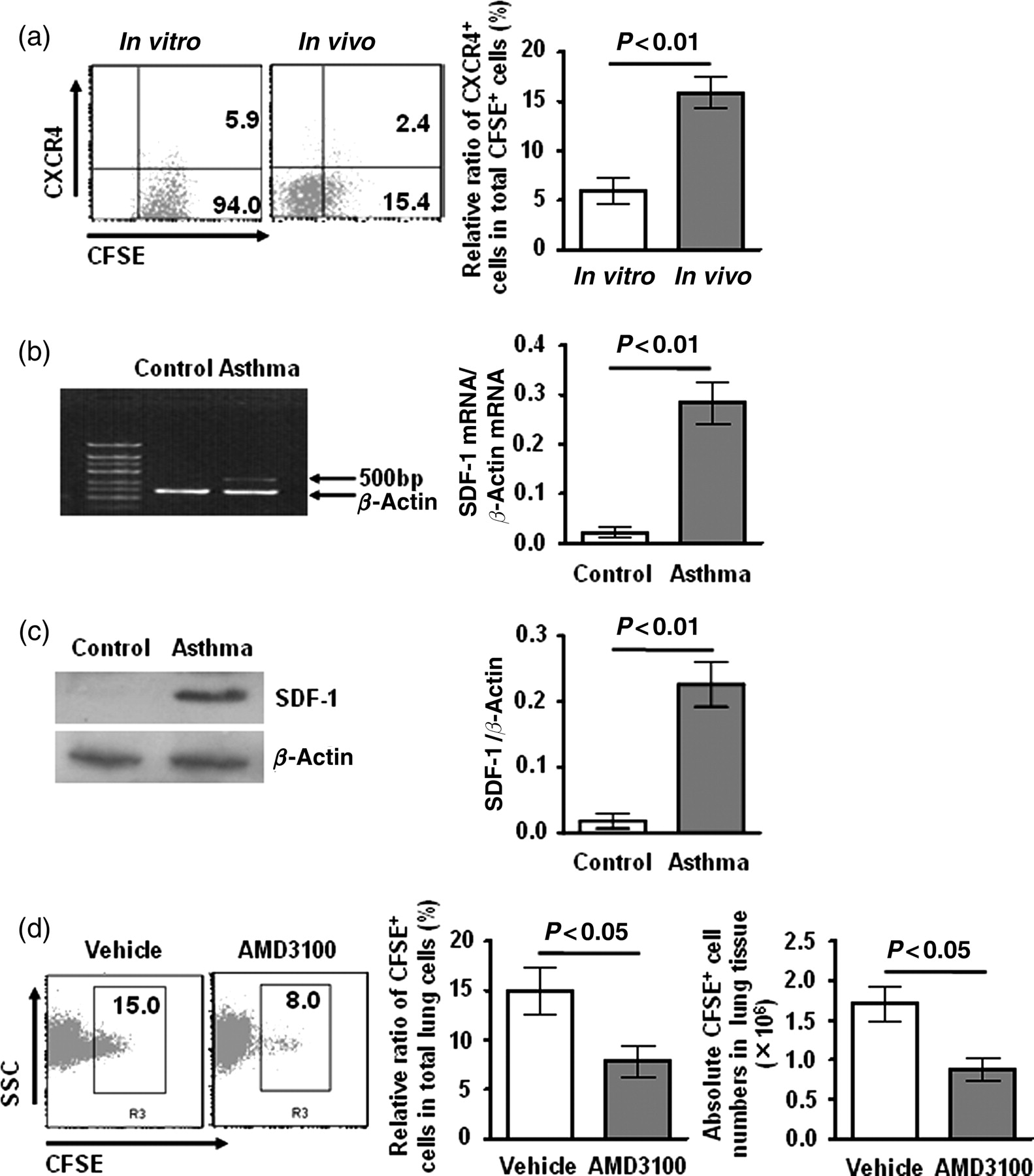
Role of the CXCR4/SDF-1 axis in the migration of exogenous MSCs. (a) Expression of CXCR4 in CFSE+ cells. Experimental groups, treatments and data analysis were identical with Figure 1b. (b) and (c) Expression of SDF-1 in lung tissues. Mice were sensitized twice with OVA on days 1 and 8, and then challenged with OVA on days 15, 16 and 17. Control mice were treated with PBS. Lung tissues were collected on day 18. RT-PCR and Western blotting were performed as described in ‘Methods’. Data are the mean ± SD from three experiments (n = 15). (d) AMD3100 treatment decreased the migration of MSCs to lungs in asthma. MSCs were treated with AMD3100 or vehicle in vitro prior to adoptive transfer. Transfer protocol and flow cytometric analysis were identical with Figure 1a. Data are the mean ± SD from three experiments (n = 15). MSC, mesenchymal stem cells; SDF-1, stromal cell-derived factor-1α; PBS, phosphate-buffered saline; CFSE, 5(6)-(N-succinimidyloxycarbonyl)-3′,6′,0,0′-diacetylfluorescein; OVA, ovalbumin
Expression of specific adhesion molecules or other proteins by the injured lung may be important for the recruitment of cells. 10,11 Next, we detected the expression of SDF-1, a ligand of CXCR4, in lung tissues. The results showed that the expression of SDF-1 significantly increased in OVA-induced asthmatic mice both in mRNA level and in protein level compared with PBS-treated control mice (Figures 2b and c, SDF-1 mRNA: P < 0.01; SDF-1: P < 0.01).
To further clarify the role of CXCR4/SDF-1 axis in the migration of exogenous MSCs to lungs in asthma, we used a non-toxic CXCR4 antagonist AMD3100, which is a tight-binding and slowly reversible antagonist. MSCs were treated with AMD3100 in vitro prior to adoptive transfer. As shown in Figure 2d, the relative ratio and the absolute number of CFSE+ cells significantly decreased in the AMD3100-treated group compared with the vehicle-treated group. The results indicated that blockage of CXCR4 decreased the migration of exogenous MSCs to lungs in asthmatic mice.
Reduced allergic airway inflammation and airway hyperreactivity by exogenous MSCs
Having confirmed the migration of MSCs to lungs, we investigated the functional role of MSCs in asthma. Firstly, we studied the effect of MSCs transfer on the airway environment, having established that no changes in airway cell composition or BALF cytokine secretion in mice occurred as a result of MSC transfer per se (data not shown). Mice were sensitized twice with OVA on days 1 and 8. Then, airway challenges were performed on days 15, 16 and 17 after MSC transfer on day 14. Recovery of BALF was performed on day 18. Administration of exogenous MSCs significantly reduced airway cellular infiltrates (Figure 3a, P < 0.05). Differential cell counting revealed a profound reduction of airway eosinophilia in MSC-transferred mice (Figure 3b, P < 0.05).
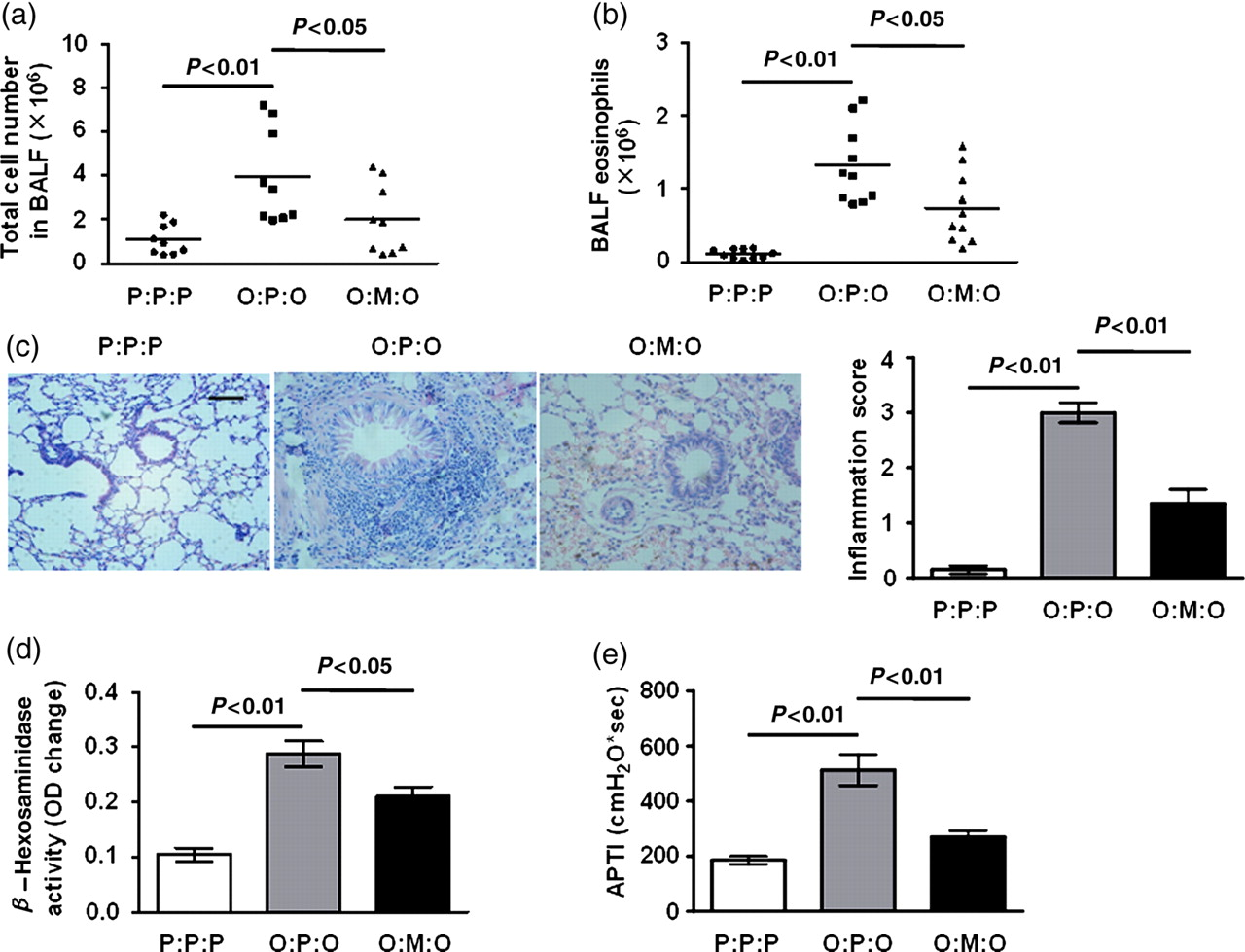
Airway cell infiltration, tissue pathology and AHR were inhibited by exogenous MSCs. Mice were sensitized twice with OVA or PBS on days 1 and 8. MSC transfer was performed on day 14. On days 15, 16 and 17, airway challenge was performed. 24 h after final airway challenge (day 18), measurement of airway responsiveness, collection of BALF and histological sections of lung tissues were performed. Data are the mean ± SD from three experiments (n = 10). (a) Total lavage cell numbers. (b) Eosinophil numbers. (c) Formalin-fixed lung 5-μm sections stained with H&E and semiquantitative peribronchial inflammation scores. Scale bar, 40 μm. (d) β-Hexosaminidase in BALF. (e) AHR on acetylcholine challenge. AHR, airway hyperreactivity; H&E, hematoxylin and eosin; BALF, bronchoalveolar lavage; APTI, airway pressure-time index; OVA, ovalbumin. (A color version of this figure is available in the online journal)
To determine whether suppressed airway cellular infiltration represented a more general down-modulation of pathology, lung histological sections were compared. Hematoxylin and eosin staining was used to characterize cellular infiltrates. In addition, mast-cell degranulation was estimated by measuring levels of β-hexosaminidase in BALF. In asthmatic control mice, airway challenge leads to a dense peribronchial and perivascular inflammatory infiltrate of lymphocytes, and mononuclear and polymorphonuclear cells with epithelial shedding and extended columnal cells (Figure 3c). In MSC-treated mice, however, tissue inflammation was greatly reduced with significantly less peribronchial and perivascular cellular infiltration (Figure 3c). In addition, the allergen-induced increase in BALF β-hexosaminidase, indicating mast-cell mediator release, was also attenuated in MSC-treated mice (Figure 3d, P < 0.05). Thus, exogenous MSCs can protect mice against a range of allergic airway inflammatory pathologies, including the infiltration of inflammatory cells and mast cell degranulation.
We also examined the capacity of exogenous MSCs to counterbalance airway hyperreactivity (AHR), as assessed by APTI. Figure 3e shows that asthmatic control mice expressed significant AHR. However, administration of exogenous MSCs significantly reduced AHR.
Suppression of local type 2 effector cytokines and reversion of Th2 bias by MSCs
To further analyze the mechanism of the inhibitory effect of MSCs, we measured local cytokine IL-5 and IL-9 levels in BALF. The levels of IL-5 and IL-9 were both elevated after airway challenge in the BALF of asthmatic control mice (Figures 4a and b). However, IL-5 was significantly decreased by MSC transfer (Figure 4a, P < 0.05). Reductions in the key agents in the mobilization and extravasation of eosinophils provided a mechanistic explanation for the dramatically reduced airway eosinophilia in MSC-treated mice. Similarly, the elevation of IL-9 level was also reversed by MSC transfer (Figure 4b, P < 0.05), which may explain the decrease of mast-cell mediator release in MSC-treated mice.
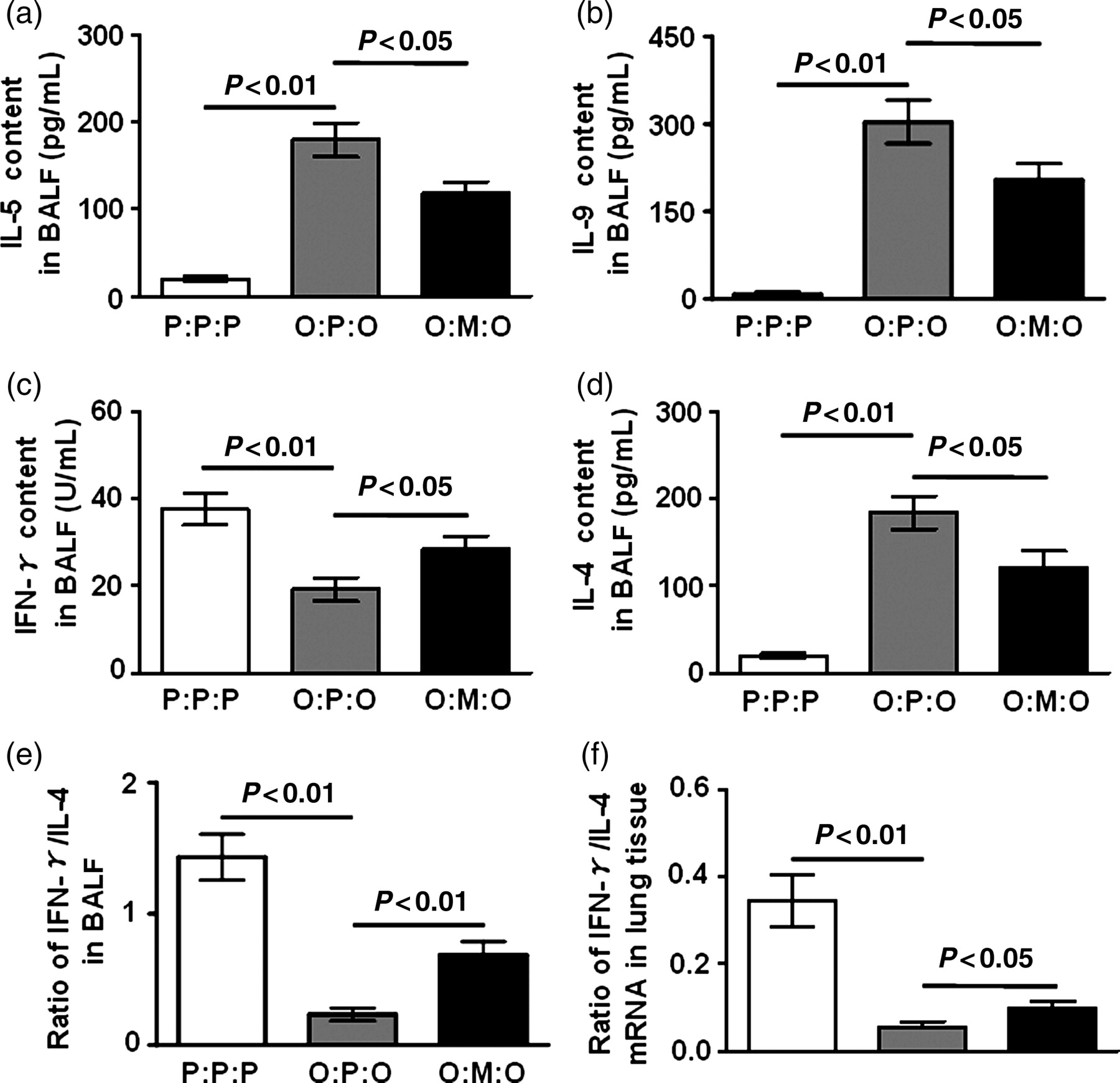
Suppression of local type 2 effector cytokines and reversion of Th2 bias by MSCs in asthmatic mice. Data are the mean ± SD from three experiments (n = 10). (a), (b), (c) and (d) Cytokine responses in BALF. BALF from three group mice were assayed for the indicated cytokines. (e) and (f) Reversion of Th1/Th2 bias by MSCs in asthmatic mice. (e) The ratio of IFN-γ/IL-4. (f) The levels of IFN-γ and IL-4 mRNA isolated from lung tissue were quantified by the realtime PCR SYBR Green System (Takara, Dalian, China). The results were presented as the expression of the individual mRNAs with normalization to β-actin, using the cycle threshold method. BALF, bronchoalveolar lavage fluid; IFN-γ, interferon-γ; IL, interleukin
MSCs can inhibit the proliferation and function of T-cells, B-cells and dendritic cells. 3 To examine the effect of exogenous MSCs on Th2 bias in asthma, IFN-γ and IL-4 levels in BALF were measured. As shown in Figures 4c and d, decreased IFN-γ level and enhanced IL-4 responses were observed in asthmatic control mice. Meanwhile, the ratio of IFN-γ/IL-4 was lower in asthmatic control mice (Figure 4e, P < 0.01). However, compared with asthmatic control mice, elevated IFN-γ level (Figure 4c, P < 0.05) and decreased IL-4 level (Figure 4d, P < 0.05) were shown in MSC-treated mice. Correspondingly, the ratio of IFN-γ/IL-4 was elevated in MSC-transferred mice (Figure 4e, P < 0.01).
We also detected the levels of IFN-γ and IL-4 mRNA in lung tissue by realtime PCR, and the ratio of IFN-γ/IL-4 mRNA was analyzed. The data show that it was significantly higher in MSC-treated mice than in asthmatic control mice (Figure 4f, P < 0.05). These data indicate that MSCs could induce a shift from Th2 to Th1 response in the animal model of asthma and altered the highly polarized type 2 cytokine environment both in the local airway and in whole lung tissue.
Discussion
Several studies have shown that MSCs have significant immunomodulatory effects in several lung injury animal models induced by physical and chemical factors. Intratracheal or systemic administration of MSCs immediately after intratracheal bleomycin administration decreased subsequent lung collagen accumulation and fibrosis. 10 Intratracheal administration of MSCs decreased pulmonary hypertension and monocrotaline-induced pulmonary vascular injury. 12 Our current report describes that exogenous MSCs can reduce the infiltration of inflammatory cells, mast cell degranulation and the production of type 2 cytokines in a mouse model of allergic asthma. Our results expand our understanding of the immunomodulatory role of MSCs to lung allergic inflammation.
Firstly, we confirmed exogenous MSCs can relocate to lung tissues in asthmatic mice, which hinted that exogenous MSCs might have some functional role in the development of asthma. Then, we analyzed the chemotactic mechanism of MSCs. CXCR4 expression on the cell surface of MSCs was up-regulated in asthma. Simultaneously, OVA challenge increases lung expression of SDF-1. Using a CXCR4 antagonist, we found that though only 6% of MSCs express CXCR4 in vitro, inhibition of CXCR4 still decreased the migration of MSCs to lungs. Thus, we presented direct evidence about the role of the CXCR4/SDF-1 axis in the recruitment of exogenous MSCs to lungs in a mouse model of asthma. MSCs also express cell surface adhesion molecules including vascular cell adhesion molecule-1 and CD44. Increased expression of the CD44 ligand hyaluronan is reported to provide a homing signal for CD44-expressing MSCs in bleomycin-induced lung injury in mice. 10 However, no significant change of CD44 expression was observed in MSCs in our study (data not shown).
Then, we found that exogenous MSCs can attenuate OVA-induced airway inflammatory pathologies, including the infiltration of inflammatory cells, mast cell degranulation and AHR. Downregulation of airway inflammation and AHR by exogenous MSCs was associated with a significant reduction of IL-5 and IL-9 production and a reversion of Th2 bias. The data indicate that MSCs may directly or indirectly inhibit the exaggerated Th2 response in asthma in vivo. Our results are consistent with a very recent report, which showed that adipose tissue-derived stem cells could inhibit eosinophilic inflammation partly via a shifting from Th2 to Th1 immune response in an experimental allergic rhinitis model. 13 However, MSCs may also shift Th1 immune responses to Th2 responses in allografts, 10 multiple sclerosis 14 and acute kidney injury. 15 Thus, the immunomodulatory effects of MSCs may be influenced by the inflammatory environment, which seems to be in an immunosuppressive manner overall, irrespective of the type of T-cell immune response. Very recently, Nemeth et al. 16 reported that bone marrow stromal cells could effectively suppress allergic response in a mouse model of ragweed-induced asthma, which was accompanied with more CD4+Foxp3+ Tregs presented in lung tissue. However, the origin and the exact role of the Treg subtype are still unknown. So, further studies are needed to clarify the exact mechanism of a Th2 to Th1 shift in our study.
In conclusion, this study showed that exogenous MSCs can migrate to sites of inflammation in a mouse model of asthma through an SDF-1/CXCR4-dependent mechanism. Administration of a single dose of MSCs can attenuate allergic airway inflammation and AHR partly via a shift from a Th2 to Th1 immune response. Some clinic trials have demonstrated safety of autologous or allogeneic MSC administration in treatment-resistant patients with Crohn's disease and graft-versus-host disease, in which no significant adverse effects have been observed. 17,18 Furthermore, MSCs are also increasingly described as vehicles for delivery of therapeutic genes and proteins in several lung diseases, including pulmonary hypertension 19 and acute lung injury 20 . Taken together with our findings, immunotherapy based on MSCs may also be a feasible, efficient therapy for asthma.
Footnotes
ACKNOWLEDGeMENTS
The work was supported by grants from the National Natural Science Foundation of China (30770927, 30900653).