
Abstract
Chronic stress, as seen in post-traumatic stress disorder, can exacerbate existing diseases. Electroacupuncture (EA) has been proposed to treat chronic stress, although information on its efficacy or mechanism(s) of action is limited. While many factors contribute to the chronic stress response, the sympathetic peptide, neuropeptide Y (NPY), has been shown to be elevated in chronic stress and is hypothesized to contribute to the physiological stress response. Our objective was to determine if EA at acupuncture point stomach 36 (ST36) is effective in mitigating cold stress-induced increase in NPY in rats. Both pretreatment and concomitant treatment with EA ST36 effectively suppressed peripheral and central NPY after 14 d of cold stress (P < 0.05). The effect was specific, as NPY in Sham-EA rats was not different than observed in stress-only rats. Additionally, the effect of EA ST36 was long-lasting, as NPY levels remained suppressed despite early cessation of EA ST36, while exposure to cold stress was continued. In the paraventricular nucleus (PVN), it was notable that changes in NPY mirrored plasma NPY levels, and that the significant elevation in PVN Y1 receptor observed with stress was also prevented with EA ST36. The findings indicate that EA ST36 is effective in preventing one of the sympathetic pathways stimulated during chronic stress, and thus may be a useful adjunct therapy in stress-related disorders.
Introduction
Chronic stress has been linked with cardiovascular disease and obesity, 1–10 and has been shown to depress immune function, possibly influencing the course of cancer and infections. 11–13 Effectively blocking the stress pathways remains a challenge, and new therapies are continually being sought: acupuncture is a possible candidate. Acupuncture is the most common treatment modality in traditional Chinese medicine (TCM), 14 and electroacupuncture (EA), the electric stimulation of acupuncture needles, has been utilized by the Chinese for treating numerous diseases. According to the World Health Organization, acupuncture is useful as adjunct therapy in more than 50 disorders including chronic stress; 15 however, the mechanisms by which EA acts are unclear.
Traditionally, a common TCM acupoint, Zusanli (ST36), has been used in patients for a variety of conditions including stress. 14 However, reports on mechanisms of EA actions during stress are limited, and existing studies have focused on acute stress rather than chronic stress responses. 16–18 Experiments during acute stress show that EA ST36 blocks chronic stimulation of the hypothalamic–pituitary–adrenal axis (HPA), and thus might reduce the physiological effects of the acute stress. Indeed, Sun et al. 17 found that EA ST36 in their rat model was effective at reducing corticosterone concentrations, and modulated the expression of nitric oxide synthase1–3, possibly playing a role in protecting gastric mucosa from acute cold-induced stomach ulcers. Such studies, while intriguing, do not address the potential for EA in chronic stress states or its mechanism of action. Our broad hypothesis is that EA acts through several stress pathways including the HPA and sympathetic nervous system (SNS). Initially, we examined the effect of ST36 on one component of the SNS, neuropeptide Y (NPY).
NPY is primarily synthesized and released by sympathetic neurons both centrally and peripherally; it is also released from platelets in many species. 19–21 Importantly, plasma NPY is elevated in response to stress, and is proposed to contribute to the deleterious physiological effects of chronic stress. 22,23 Various stress models have provided evidence that NPY also increases in brain regions such as the amygdala and the paraventricular nucleus (PVN), 24,25 as well as in sympathetic ganglia. 26–28 These stress-induced effects have been mainly attributed to the action of NPY at the NPY Y1 receptor (Y1R). 25 Using a previously established model of non-habituating chronic cold stress, 29 we examined the effect of EA ST36 on central and peripheral NPY in rats.
Methods
Animals
All animal experiments were approved by the Georgetown University Animal Care and Use Committee. Adult male, 11–12-week-old, Sprague-Dawley rats (Harlan Laboratories, Inc, Dublin, VA, USA) weighing 290–420 g were included in this study. The rats were received with 23G round-tip indwelling jugular vein catheters (JVCs). On receipt, they were weighed and randomly assigned to one of either four or five groups depending on the experiment. Animals were assigned to one or two control groups; a stress-only group; a Sham-EA group that received the stress treatment and non-TCM EA; and finally a TCM-EA group.
Rats were housed one per cage in a controlled environment at a constant temperature (23°C) and maintained in a 12-h light/dark cycle with free access to water and regular rat chow. Catheter care was conducted on the day after the animals had arrived, and then again on days one and seven of the experiment. Maintenance consisted of drawing back on the catheter with a 1-cm3 sterile syringe utilizing a 23G blunt tip sterile stainless steel needle (Small Parts, Inc, Seattle, WA, USA), followed by instilling 0.1 mL of the Harlan recommended lock solution: 5 mL 100% sterile glycerin (Fisher Scientific, Pittsburgh, PA, USA) with 5 mL 1000 U/mL heparin sodium.
All animals were acclimated similarly for four days before the onset of interventions. Acclimation included three minutes of touch and placing the animal twice a day in a cotton sock, which was subsequently used in the study to briefly hold the animal during the acupuncture procedures. The control animals received no further treatments once the experiment was begun.
Acupuncture
Sham-EA needling
Acupuncture treatment for the Sham-EA group consisted of placing the animal into the sock that had been utilized for acclimation. Sterile, stainless steel, acupuncture needles measuring 34G (0.22 mm) and 1 inch (25 mm) (Millennia, Shanghai, China) were inserted bilaterally into a randomly designated non-TCM acupuncture point on the back, 2 cm lateral to the tail region.
TCM acupuncture needling
The rats in the TCM EA group received acupuncture treatments utilizing the same sock and needles as described above. However, in this cohort, the location of the needles was at TCM point Zusanli (Stomach, ST36) on bilateral hind legs of the animal. These points were identified using rat mapping for TCM acupuncture (kindly provided by the laboratory of Dr Lixing Lao, University of Maryland School of Medicine, Baltimore, MD, USA).
EA treatments
Once the acupuncture needles described above were inserted bilaterally, they were attached to electrodes. Animals were then singly placed in new cages with electrodes threaded through the cage lid. Electrodes were then attached to a power source (Model AWQ-104L, purchased from UPC Medical Supplies, St Gabriel, CA, USA) and the points were stimulated for 20 min at a frequency of 10 Hz with 2 mA output, and a pulse width of 0.1 s for 20 min, based on previous findings. 30,31 The animals did not receive food or water for the duration treatment. All acupuncture treatments occurred between 09:00 and 12:00 depending on the experiment, and were administered either 30 min before the stressor or 30 min after the animals had acclimated to ambient room temperature, in their home cages.
Two EA ST36 paradigms were assessed: PRE (EA prior to initiation of stress) – to determine whether pretreatment with EA could diminish the stress response, animals received 20 min of either Sham or EA ST36 treatment for five days prior to 10 d of cold stress. Control (n = 7), Sham-EA (n = 10), Stress (n = 7), EA ST36 (n = 10); and POST (EA begins following exposure to stress) – to determine whether EA treatment after initial exposure to stress effectively diminishes the stress response, rats received Sham or EA ST36 treatment for 20 min following one hour of cold stress for 14 d. Control (n = 7), Sham-EA (n = 13), Stress (n = 7), EA ST36 (n = 13). In this model, animals received one-hour cold stress, were then returned to their home cage for 30 min at room temperature, followed by 20 min of Sham or EA ST36. To assess whether the effects of EA ST36 were long lasting, we ceased treatment in four animals from the POST Sham-EA and EA ST36 groups four days before the experiment ended, while continuing with stress.
Stress
Animals were randomly assigned to treatment groups. Cold stress was carried out as previously described. 32 Briefly, animals were placed in cages with 1 cm deep crushed ice for one hour per day for 14 d. Following cold stress, animals were returned to their home cage.
NPY enzyme-linked immunosorbent assay
Blood was obtained via the JVCs using 23G blunt stainless steel needles (Small Parts Inc) in 3-mL sterile syringes. In general, 1.3 mL of whole blood was collected in 1.5-mL ethylenediaminetetraacetic acid tubes and centrifuged at 4°C at 400
Immunohistochemistry analysis
Animals were anesthetized using 30–50 mg/kg pentobarbital (intraperitoneally). Brain hemispheres were harvested and cut in half for immunohistochemistry and quantitative reverse transcription polymerase chain reaction (RT-PCR) analysis. Tissues were post-fixed with 10% buffered formalin, paraffin-embedded and prepared as 5 μm-thick sagittal sections. Slides were deparaffinized with xylene and rehydrated through a graded alcohol series, starting with 100% EtOH and ending with 60% EtOH. Slides were immersed in 10 mmol/L citrate buffer (pH 6.0) with 0.05% Tween at 98°C for 20 min. Horseradish peroxidase (HRP)-conjugated antirabbit secondary antibody (Envision-Plus; DAKO USA, Carpinteria, CA, USA) was applied, and the HRP was detected using DAB chromagen (DAKO USA). Similarly treated serial sections with the primary antibody omitted were used as negative controls.
Sections were visualized with the CRi Nuance FX microscope and pictures were taken with the CRi Nuance v2.6.0 camera (Caliper Life Sciences, Hopkinton, MA, USA). The PVN was identified using rat brain map, 34 in consultation with a neuroanatomist (Dr Nabil Azzam), and semiquantitatively analyzed for positive NPY staining via the MDS Analytical Technologies Metamorph v7.5.5.0 (Sunnyvale, CA, USA).
Quantitative realtime RT-PCR
Total RNA was isolated from resected brain PVN using the previously described phenol–chloroform extraction method. 35 One microgram of RNA per sample was used for cDNA synthesis via the iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA). Realtime RT-PCR was done using the ICycler iQ Detection System (Bio-Rad). The PCR reactions were carried out using TaqMan Universal PCR Master Mix and predesigned primers and fluorescein-labeled probes (Applied Biosystems, Foster City, CA, USA). The primers were as follows. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH): 5′ CCTTCATTGACCTCAACTAC 3′, 5′ GGAAGGCCATGCCAGTGAGC 3′; NPY: 5′ CAGAGGCGCCCAGAGCAG 3′, 5′ CAGCCCCATTCGTTTGTTACC 3′; Y1R: 5′ CTCTTGCTTATGGRGATGTGA 3′, 5′ CTGGAAGTTTTTGTTCAGGAAYCCA 3′. The results were calculated by the comparative threshold method using GAPDH as an endogenous reference gene, according to the Applied Biosystems ABI PRISM 7700 User Bulletin no. 2.
Statistical analysis
Data are presented as mean ± SEM and analyzed by one-way analysis of variance with Tukey multiple t-test or Kruskal–Wallis post-test depending on the sample size to compare between treatment groups using GraphPad Prism v4 (GraphPad Software, La Jolla, CA, USA). P < 0.05 was considered statistically significant for the indicated n per group.
Results
Effect of EA on plasma NPY in chronically stressed rats
Control values for platelet-poor plasma NPY were comparable (within normal levels) in both PRE and POST groups on the pretreatment day (day 1, Figure 1a, left and right panels). By day 14, NPY was significantly higher in the stress-only and Sham-EA groups compared with controls (Figure 1a). In contrast, EA at ST36 prevented the stress-induced increase in NPY. Indeed, NPY levels in the EA ST36 rats were not different from controls, but were significantly lower than the Sham-EA and stress-only animals (Figure 1a). These findings were consistent in both the PRE and POST EA ST36 animals, indicating that EA ST36 is effective in preventing the rise in plasma NPY, whether given following or prior to the stress.
Effect of EA given before and after stress on plasma NPY in chronically stressed rats. (a) Plasma NPY levels on days 1 (pre-treatment), and 14, respectively. PRE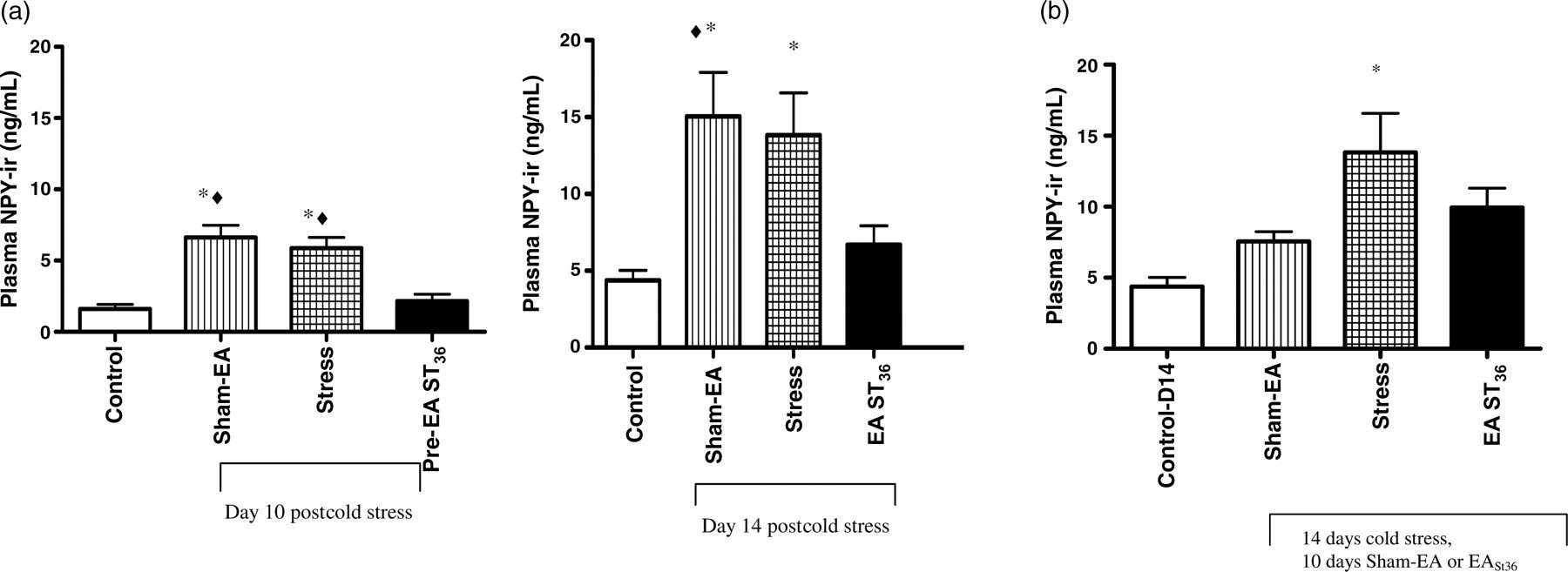
To investigate if the effects of acupuncture on the plasma NPY levels are transient or long lasting, we also analyzed NPY levels in animals where EA treatments were stopped during the last four days of cold stress in POST animals. At day 14, as previously seen, NPY in the stress-only animals was significantly greater than seen in the control, non-stressed group (P < 0.05), while EA ST36 was not different from controls (Figure 1b)
Effects of EA on NPY Y1R expression in the PVN
Central NPY immunoreactivity was examined by immunohistochemistry in tissue from the PVN. There were significant increases in NPY in the PVN of stress-only and Sham-EA groups (Figure 2a), and the effect of EA ST36 to prevent the increase in NPY protein was similar to effects observed in PVN NPY mRNA (Figure 2b). The changes in gene expression mirrored the effects observed in the plasma (Figure 1). However, these changes in the brain NPY were different than those observed in the adrenal glands, where NPY mRNA expression was not different between the groups Effect of EA on NPY protein and message in the PVN of chronically stressed rats. (a, b) NPY immunohistochemistry in the PVN. NPY immunoreactivity was significantly higher in the stress + Sham-EA and stress-only animals when compared with the control group (*P < 0.0001). EA ST36 prevented this increase. (c) NPY mRNA expression in the PVN. NPY mRNA (normalized by GAPDH) was also significantly higher in stress and stress + Sham-EA than either control or stress + EA ST36 animals (♦P < 0.05). NPY, neuropeptide Y; EA, electroacupuncture; PVN, paraventricular nucleus; GAPDH, glyceraldehyde-3-phosphate dehydrogenase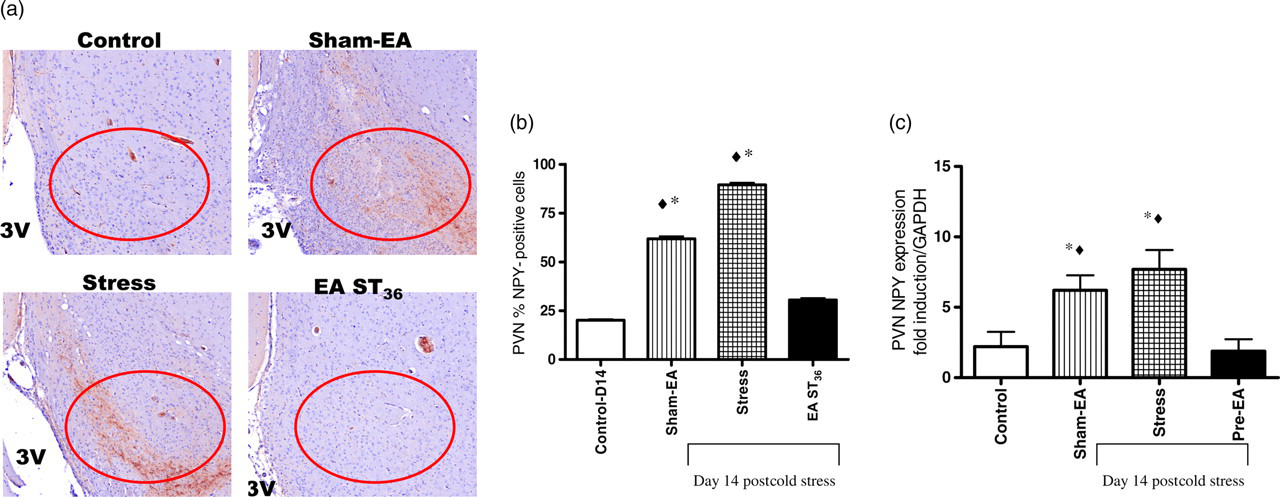
As the Y1R is considered the predominant receptor for NPY action in the PVN during chronic stress,
17
Y1R mRNA expression was examined in the PVN from control and stressed animals. There was a significant induction of Y1R mRNA in the PVN of stress-only animals (P < 0.05 from controls), which was prevented by EA ST36 (Figure 3). Y1R mRNA was also significantly increased by Sham-EA (P < 0.05 from controls), further confirming the specificity of the EA ST36. The Y2 receptor did not vary between the groups, supporting the findings that the Y1R effect was specific (data not shown).
Effect of EA on NPY Y1 receptor mRNA expression in the PVN of chronically stressed rats. NPY Y1 receptor expression (normalized by GAPDH) was significantly higher in the stress + Sham-EA and stress-only animals when compared with the controls (*P < 0.05). EA ST36 prevented the stress-induced increase in NPY Y1 receptor. NPY, neuropeptide Y; EA, electroacupuncture; PVN, paraventricular nucleus; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
Discussion
The current study demonstrates that EA specifically at ST 36 is effective in preventing the chronic stress-induced increase in circulating and central NPY. In separate experiments, the cold stress-induced three-fold increases in plasma NPY were blocked by both pre- and post-treatment with EA at ST36 (but not with Sham-EA). In addition, this effect appears to be long lasting, providing suppression of NPY for four days after EA ceased. Finally, these changes in peripheral NPY were also observed in NPY protein in the PVN of the hypothalamus. This ability to block NPY has elucidated a potential mechanism of action for acupuncture's efficacy in treatment of chronic stress. We have therefore added to the literature by reporting for the first time that both PRE- and POST- EA ST36 are effective in preventing the chronic stress-induced increases in plasma and PVN NPY, and that the effect may be long lasting.
The plasma NPY stress-induced response was noted at 14 d, but not at seven days (data not shown), supporting the role of NPY as contributing to the chronic response to stress. These findings support the previous work showing that cold stress increases NPY in plasma. 36–38 EA has been demonstrated to decrease NPY in other physiological stress conditions, including diabetes and chronic food restriction. Lee et al. 39 found that acupuncture at ST36 decreased NPY expression in the hypothalamus of rats with streptozotocin diabetes, reducing diabetic hyperphagia. Tian et al. 40 reported that EA decreased expression of gastric ghrelin and hypothalamic NPY in chronic food-restricted rats when compared with their controls. Our studies extend these reports by showing that EA ST36 is also effective at blocking central and plasma NPY during chronic stress. In addition, we determined that when EA was withdrawn during cold stress, NPY continued to be low in EA ST36 rats. No other studies have demonstrated these potentially long-lasting effects of EA ST36.
Another important aspect of this work is the central effects of EA ST36 on NPY in the PVN, which mirrored that of the NPY in the plasma. The chronically stressed animals had significantly higher Y1R mRNA expression when compared with the control animals; the increase was prevented by EA ST36. These central effects of EA ST36 make the overall effects more compelling and relevant, since changes in brain NPY have also been postulated in humans during times of stress. 41
It is unclear at what point the EA ST36 may block the central and peripheral response to release NPY during chronic stress; however, we speculate that EA ST36 may act to block one or more pathways. Indeed, other studies in our lab indicate that EA ST36 is also effective in blocking the HPA (unpublished results), which may also modulate the NPY response. This, in addition to the potential effect of EA ST36 to block the SNS, may comprise an important mechanism to suppress HPA and sympathetic outflow in response to chronic stressors.
In our model of chronic cold stress, peripheral and central NPY levels were significantly increased, which is in accordance with other reports. 17–20 Thus, one of the mechanisms by which EA reduces chronic stress may be by suppressing the NPY pathway and reducing the stress-induced increase in central Y1R; however, in the absence of the protein data, conclusions on the receptors have to be viewed with caution. This study suggests that EA ST36 is effective in preventing one of the sympathetic pathways stimulated during chronic stress, and thus may be a useful adjunct therapy in treating chronic stress-related disorders.
Footnotes
ACKNOWLEDGEMENTS
The research was funded by the American Association of Nurse Anesthetists (AANA) doctoral fellowship award to LE and NIH/NCCAM grant no. K07-AT001193–02 NIH/NCCAM to HA.