
Abstract
Obesity is considered a systemic low-grade inflammatory state. Although the spleen is the main immune organ with a close anatomical relationship with the liver, its role in the progression of fatty liver disease remains uncertain. Therefore, we sought to clarify the functional role of the spleen in the development of steatohepatitis in high-fat (HF)-diet-induced obese rats. Male Sprague-Dawley rats were fed HF food and divided into two groups, a splenectomy (SPX) group and a sham-operation (Sham) group. The liver and abdominal white adipose tissue (WAT) were removed one and six months after surgery, and we evaluated the effects of SPX on WAT and HF-induced fatty liver. SPX rats exhibited worse dyslipidemia and inflammatory changes in WAT one month after surgery. Hepatic steatosis and inflammation were accelerated by SPX, based on the time after surgery. At one month after surgery, the tissue triglyceride content increased in SPX rats, compared with Sham controls (P < 0.05). The liver histology also showed a worsening of steatosis in those rats. At six months after SPX, dramatic inflammatory and fibrotic changes were observed in liver tissue sections. Hepatic carnitine palmitoyltransferase-1 was suppressed at one and six months after SPX (P < 0.05 for each). WAT and liver tissue levels of inflammatory markers such as tumor necrosis factor-α, and the expression of Kupffer cells were all increased at six months in SPX rats, compared with Sham controls (P < 0.05 for each). Our results indicate that the preservation of the spleen contributes to the prevention of the progression of hepatic steatosis to steatohepatitis in obese rats.
Introduction
Obesity is associated not only with fat accumulation but also with chronic inflammation, characterized by abnormal cytokine production and activation of a network of inflammatory signaling pathways. 1–5 The altered production of proinflammatory cytokines, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1 and IL-6, in adipocytes has been implicated in chronic inflammation in adipose tissue. 6 Furthermore, a marked increase of macrophage accumulation in omental white adipose tissue (WAT) in obese subjects also contributes to a series of metabolic disorders. 3,7,8
In addition to adipose tissue, the liver is also involved in obesity and related metabolic disorders. Previous studies have demonstrated that omental WAT macrophage accumulation is associated with marked hepatic inflammation in obese humans, 9 and intrahepatic fat accumulation accompanied by high-fat (HF) feeding leads to hepatic inflammation through activation of the nuclear factor-κB pathway and downstream cytokine production, including IL-1β, IL-6 and TNF-α in the liver. 10 These proinflammatory cytokines and fatty acids produced by hepatocytes in response to steatosis and released from visceral WAT activate liver Kupffer cells (KCs). KCs are macrophages resident in the liver and account for 80–90% of the total population of fixed tissue macrophages in the body. 11 Activated KCs produce hepatic proinflammatory cytokines, which further activate KCs in a positive-feedback mechanism. 12,13 In particular, hepatic TNF-α is a major inducer of obesity-related metabolic disorders such as insulin resistance 14 and steatohepatitis. 15,16
Non-alcoholic steatohepatitis (NASH) in humans is considered to be the hepatic manifestation of metabolic syndrome. 17 NASH progresses to more severe stages of liver injury, such as fibrosis, cirrhosis and even hepatocellular carcinoma. 2,18 Previous studies have stated that hepatic accumulation of inflammatory cells is generally greater in NASH than in simple steatosis, suggesting that activation of the immune system may contribute to the progression of simple steatosis. 11 Thus, the immune system plays an important role in the pathogenesis and/or progression of obesity-induced metabolic disorders, such as fatty liver disease.
As a major immunological organ, the spleen plays a crucial role in the primary host defense immunological response against bacterial invasion in humans and rodents. 19,20 The spleen contains up to 15% of the body's fixed macrophages as well as substantial numbers of T lymphocytes and natural killer cells, and it produces various cytokines during critical states. 21,22 The spleen has a close anatomical relationship with the liver, and cytokines released from the spleen flow directly into the liver, via the splenic and portal veins, enhancing natural killer cytotoxicity in the liver. 23,24 Previous animal studies have indicated that the spleen affects the immunological function of the liver. 23,25 Both positive 25–28 and negative 29,30 roles of the spleen for hepatic cytokine production under lipopolysaccharide (LPS) treatment or endotoxemia, investigated using the livers of splenectomized animals, have been demonstrated; however, these previous reports showed the effects of the asplenic state on hepatic immune function of normal liver derived from wild-type animals, not HF-diet-induced fatty liver from obese animals.
Indeed, there is, to our knowledge, no report describing a potential relationship between the spleen and hepatic complications of obesity, such as fatty liver disease, although the spleen is a crucial immune organ connected to the liver via the portal vein. Thus, we clarified the functional role of the spleen in the pathogenesis and progression of HF-diet- and obesity-induced hepatic steatosis by analyzing the livers of splenectomized obese rats. Additionally, we investigated the changes in WAT during progression of HF-diet- and obesity-induced hepatic steatosis.
Materials and methods
Animals and surgical procedures
Male Sprague-Dawley rats (KBT Oriental, Fukuoka, Japan), eight weeks of age, were housed in a light-, temperature- and humidity-controlled room (12/12-h light/dark cycle with lights on/off at 07:00/19:00; 21 ± 1°C; 55 ± 5% relative humidity). Rats were allowed free access to chow pellets and water during the experiment. All animals were treated in accordance with the Oita University Guidelines for the Care and Use of Laboratory Animals.
Twenty-four rats were fed 60% HF food (20% protein, 20% carbohydrate and 60% fat; 5.2 kcal/g; Research Diet, Tokyo, Japan) for four weeks and then divided into two groups, a splenectomy (SPX) group and a sham-operation group (n = 12 each).
Following an intraperitoneal injection of sodium pentobarbital (50 mg/kg) and induction of deep anesthesia, a total SPX was aseptically performed through a left-sided lateral 1.5-cm subcostal incision. The splenic vessels were ligated using 4-0 silk suture, and the spleen was removed. The incision was closed in layers using running 4-0 nylon suture. A sham operation was performed through the same incision, but the spleen was returned to the abdominal cavity without ligation, and the wound was closed.
After surgery, each group was further divided into two subgroups. One subgroup was fed HF food for one month after surgery (1M) (1M-Sham, n = 6; 1M-SPX, n = 6), and the other was fed HF food for six months after surgery (6M) (6M-Sham, n = 6; 6M-SPX, n = 6). As a control group, 12 sham-operated rats were fed normal chow (CE-2; 25% protein, 50% carbohydrate and 5% fat; 3.4 kcal/g; CLEA Japan, Tokyo, Japan) from the beginning of the experiment and divided into two groups according to feeding period after splenic surgery: one month (1M-Cont, n = 6) or six months (6M-Cont, n = 6). Body weights of the rats were measured at both the time of splenic surgery and at the end of each experiment.
Measurements and blood sampling
After one or six months, the rats were anesthetized with sodium pentobarbital, as above, and perfused transcardially with isotonic phosphate-buffered saline (PBS), followed by 4% paraformaldehyde in 0.1 mol/L phosphate buffer. The liver and abdominal WAT around the epididymis were removed immediately, frozen in liquid nitrogen and stored at −80°C until protein extraction. Blood was withdrawn via cardiac puncture, and the serum was separated and immediately frozen at −80°C until analysis. Serum triglyceride (TG), free fatty acid (FFA), total cholesterol (TC), low-density lipoprotein cholesterol (LDL-c) and alanine aminotransferase (ALT) concentrations were measured using an automatic analyzer (SRL, Tokyo, Japan). The concentrations of serum high-sensitivity C-reactive protein (hsCRP), a marker of inflammation, were determined using commercial enzyme-linked immunosorbent assay (ELISA) kits (Helica Biosystems, Fullerton, CA, USA).
Histological analysis and immunohistochemistry
Liver tissue and abdominal WAT samples were fixed with 10% formalin and embedded in paraffin. Sections (5 μm thick) were cut with a vibratome, stained with hematoxylin and eosin (H&E) (both liver tissue and WAT) or silver (liver tissue) and examined under a microscope (Olympus, Tokyo, Japan).
Immunohistochemistry was performed to stain macrophages in liver tissue and WAT using an antibody against CD68 (AbD Serotec, Oxford, UK). Liver tissue and WAT slices, cut from the samples described above, were washed three times in PBS, incubated for 30 min in 0.3% H2O2 to quench endogenous peroxidase activity and transferred without rinsing to the primary antibody solution, which consisted of 5 μg/mL polyclonal anti-CD68 mouse antiserum. After 24 h at 4°C, the slices were washed three times in PBS and processed using the ABC method (Vector Laboratories, Burlingame, CA, USA). Slices were transferred to a biotinylated anti-mouse antibody solution for one hour, washed, transferred to avidin-biotinylated peroxidase for one hour, washed and then developed with diaminobenzidine substrate for 10 min. The slices were washed and mounted on slides with Permount (Dako Japan, Tokyo, Japan).
TG and cytokine contents in the liver
Liver tissue (100 mg) and abdominal WAT (300 mg) were homogenized in 2 mL of a solution containing 150 mmol/L NaCl, 0.1% Triton X-100 and 10 mmol/L Tris, using a BioMasher (Funakoshi, Tokyo, Japan) for one minute. 31 Liver TG contents were determined using a commercially available kit (Wako Chemical, Osaka, Japan). Commercial ELISA kits were used to measure the amounts of the TNF-α and IL-1β (Invitrogen, Camarillo, CA, USA) in liver tissue and abdominal WAT. Optical density readings of all samples were converted to pg/mL using standard curves generated with the recombinant cytokines supplied with the kits. Protein concentrations were determined using a protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA), to normalize the samples by protein concentration.
Western blotting
Frozen liver tissue and abdominal WAT preparations were homogenized with sodium dodecyl sulfate (SDS) sample buffer, clarified by centrifugation and boiled. The total protein concentration in each tissue sample was quantified by the Bradford method, and 10 μg of total protein per sample was loaded onto 10% SDS-polyacrylamide gels for electrophoresis. Separated proteins were transferred onto polyvinylidene difluoride membranes (Bio-Rad Laboratories). The membranes were incubated with primary antibody solution consisting of 5 μg/mL polyclonal antiserum with specificity for sterol regulatory element-binding protein-1 (SREBP-1), fatty acid synthetase (FAS), carnitine palmitoyltransferase-1 (CPT-1) (Santa Cruz Biotechnology, Santa Cruz, CA, USA) or CD68 (AbD Serotec). Immunoreactive bands were detected by enhanced chemiluminescence (Amersham Life Science, Buckinghamshire, UK) and quantified using National Institutes of Health imaging software (Scion Corporation, Frederick, MD, USA).
Statistical analysis
All data are expressed as means ± SE. Analysis of variance and a post hoc Bonferroni test were used to assess multiple comparison differences (StatView 4.0; SAS, Cary, NC, USA). A Mann-Whitney U test was used when appropriate.
Results
Changes in body weight and serum lipid profile in SPX rats
Body weight before splenic surgery (before) and the end of experiment (after)
Data are expressed as means ± SD. Differences are assessed using t-tests for continuous variables or χ 2 tests for categorical variables
*P < 0.0001 versus Cont
Serum lipid profile after splenic surgery
TG, triglyceride; FFA, free fatty acid; TC, total cholesterol; LDL-c low-density lipoprotein cholesterol
Data are expressed as means±SD. Differences are assessed using t-tests for continuous variables or χ 2 tests for categorical variables
*P < 0.0001 versus Cont; † P < 0.0001 versus Sham; ‡ P < 0.0001 versus 1M-SPX
In contrast, SPX increased the concentrations of serum TG, FFA and TC at both one and six months after splenic surgery, compared with those of sham-operated rats fed on HF diets (P < 0.0001 for each). Additionally, these increases in splenectomized rats showed time dependency after SPX, indicating that levels in 6M-SPX rats were significantly higher than in 1M-SPX rats (P < 0.0001 for each). Serum LDL-c concentrations showed a similar profile, whereas there were no significant changes between 6M-SPX and 6M-Sham rats (Table 2).
Inflammatory changes in abdominal WAT of SPX rats
To investigate the inflammatory changes in abdominal WAT after SPX, we examined WAT histology (Figure 1a), by H&E staining, and measured TNF-α and IL-1β concentrations in abdominal WAT at one (Figure 1b) and six months (Figure 1c) after splenic surgery. In abdominal WAT derived from SPX rats, invasion of inflammatory cells into the stromal area was markedly increased, compared with Sham rats (Figure 1a). Proinflammatory cytokine concentrations in WAT were also higher in SPX rats than in Sham rats (P < 0.05 for each), indicating that significant increases were present in abdominal WAT one month after SPX (Figures 1b and c).
Inflammatory changes in abdominal white adipose tissue (WAT) of obese rats are accelerated in the splenectomized (SPX) state. (a) Histological findings of abdominal WAT sections from sham-operated control rats (first row; Cont), high-fat (HF)-fed sham-operated rats (second row; Sham) and HF-fed SPX rats (third row; SPX) at one (1M) and six (6M) months after surgery were assessed by hematoxylin and eosin staining (×400). (b, c) Abdominal WAT tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) concentrations at (a) one (1M) and (b) six (6M) months after splenic surgery (n = 6 rats per group). *P < 0.05 versus Cont, **P < 0.0001 versus Cont,†
P < 0.05 versus Sham. (A color version of this figure is available in the online journal)
In addition, we performed immunohistochemistry on abdominal WAT with an anti-CD68 antibody to evaluate the number of WAT macrophages (Figure 2a), and determined CD68 expression by Western blot (Figure 2b). SPX clearly increased the macrophage population (arrows) in the abdominal WAT of obese rats (Figure 2a). CD68 levels in abdominal WAT from SPX rats were higher than those in Sham rats at both one and six months after splenic surgery (P < 0.05 for each) (Figure 2b).
The number of abdominal WAT macrophages was higher in high-fat (HF) diet-induced obese rats in the splenectomized (SPX) state. (a) Representative immunohistochemical staining using antibody against the specific macrophage marker CD68 in abdominal WAT from rats at one (1M) and six (6M) months after splenic surgery (×400). (b) Protein expression in abdominal WAT from 1M and 6M rats was measured by Western blot (n = 6 rats per group). *P < 0.05 versus Cont, **P < 0.01 versus Cont, †
P < 0.05 versus Sham. (A color version of this figure is available in the online journal)
Changes in serum hsCRP concentrations in SPX rats
We next determined serum hsCRP concentrations, which is regarded as a serum marker of systemic inflammatory reactions, in rats (Figure 3). An increase in serum hsCRP concentrations was evident in SPX rats, compared with Sham rats at one month (P < 0.05); however, no significant differences were observed at six months.
The levels of serum high-sensitivity C-reactive protein (hsCRP) increase in high-fat (HF)-diet-induced obese rats in the splenectomized (SPX) state at one month after surgery. Serum was collected from rats one (1M) and six (6M) months after splenic surgery, and hsCRP concentrations were measured. Data are shown as means ± SEM (n = 6 rats per group). *P < 0.0001 versus Cont, †
P < 0.05 versus Sham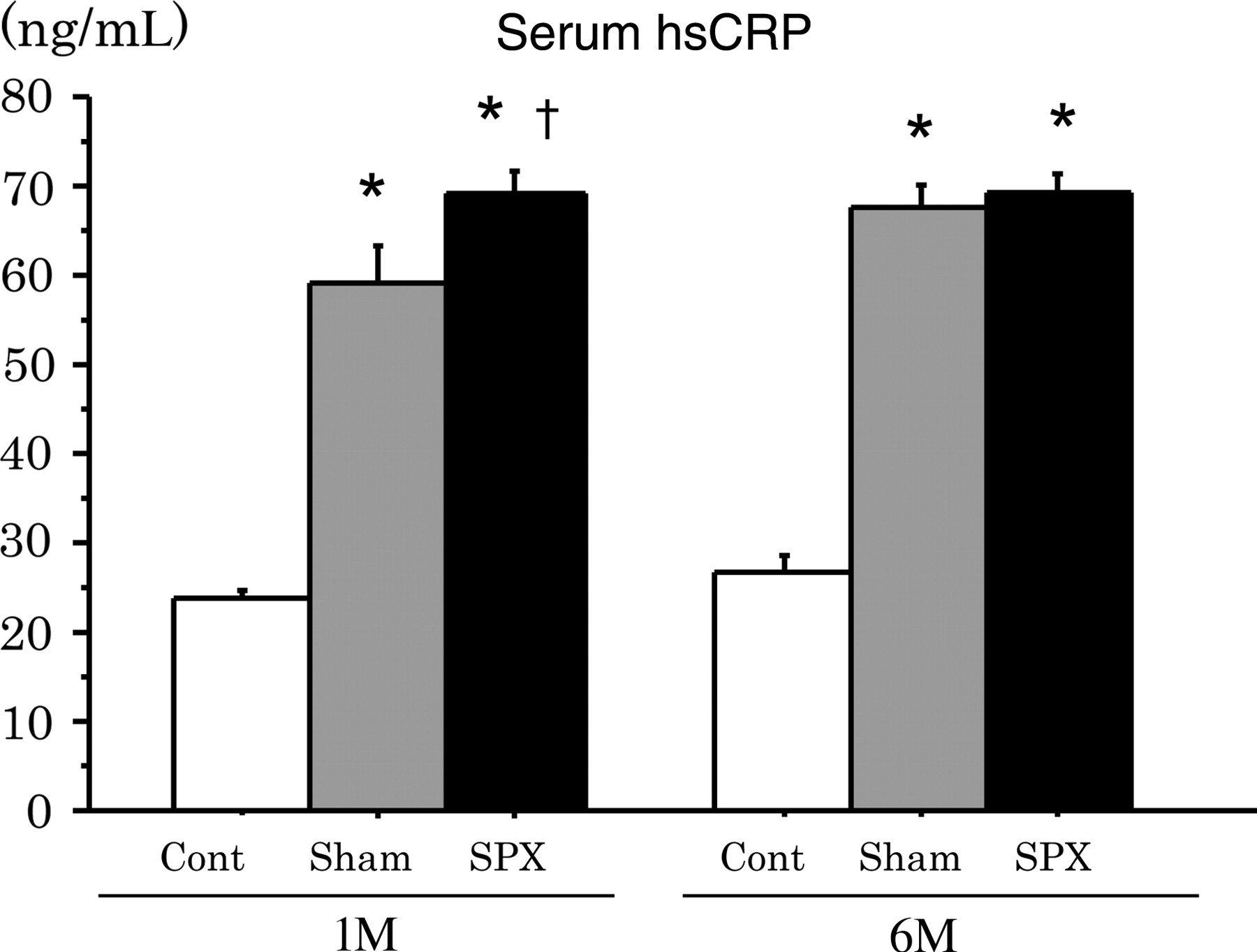
Changes in hepatic histology, liver TG content and serum ALT concentrations in SPX rats
Liver tissue histology (Figures 4a and b), as revealed by H&E staining, and hepatic lipid accumulation, as assessed by liver TG content (Figure 4c), were evaluated in rats at one and six months after SPX. At one month after SPX, a significant increase in liver TG content indicated hepatic lipid accumulation in the SPX rats compared with sham controls (P < 0.05), and liver histology showed a marked worsening of steatosis (Figure 4a). No specific histological change indicating inflammation or fibrosis was observed in the livers from 1M-SPX rats. However, the livers of 6M-SPX rats showed further lipid accumulation (P < 0.05) and dramatic inflammatory changes, such as abundant invasion by inflammatory cells and ballooning degeneration of hepatocytes (Figure 4b).
Intrahepatic lipid accumulation and liver inflammatory change in obese rats are accelerated in the splenectomized (SPX) state. (a) Histological findings of liver sections from sham-operated control rats (first row; Cont), high-fat (HF)-fed sham-operated rats (second row; Sham) and HF-fed SPX rats (third row; SPX) at one (1M) and six (6M) months after surgery were assessed by hematoxylin and eosin (H&E) staining (×400). (b) Marked inflammatory cell invasion and hepatocyte ballooning (arrow) were observed at higher (×400) magnifications in H&E-stained liver sections derived from 6M-SPX rats. (c) Hepatic triglyceride concentrations after splenic surgery. Data are shown as means ± SEM (n = 6 rats per group). *P < 0.0001 versus Cont, †
P < 0.05 versus Sham, ††
P < 0.01 versus Sham. (A color version of this figure is available in the online journal)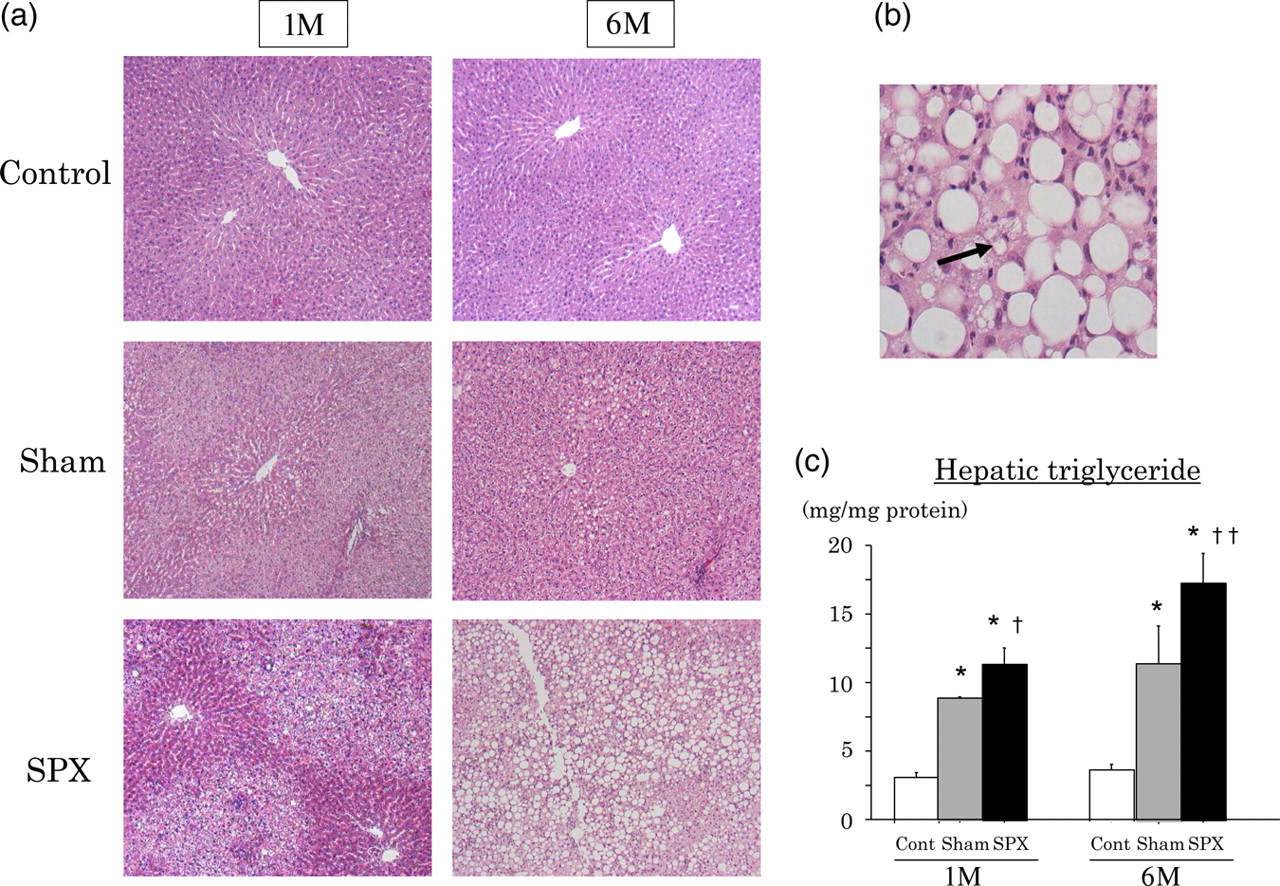
To determine the degree of liver fibrosis, silver-stained tissue slices were observed. As shown in Figure 5a, pericellular fibrosis in the area of pronounced fat deposition had developed in the livers of 6M-SPX rats, but was not seen in 1M-SPX rats. Moreover, we investigated hepatocyte injury by measuring serum concentrations of ALT in SPX rats. Serum ALT concentrations in 1M-SPX rats tended to be higher than those in 1M-Sham rats, but the difference was not significant. However, the serum ALT concentrations were increased significantly in 6M-SPX rats compared with 6M-Sham rats (P < 0.05) (Figure 5b).
Liver injury with fibrosing steatohepatitis is induced in high-fat (HF)-diet-induced obese rats in the splenectomized (HPX) state six months after surgery. (a) Representative liver sections stained with silver at one (1M) and six (6M) months after surgery (×200). First row: livers from sham-operated control rats (Cont). Second row: livers from HF-fed sham-operated rats (Sham). Third row: livers from HF-fed SPX rats (SPX). Pericellular fibrosis (arrows) was observed in the livers of 6M-SPX rats. (b) Serum sample was collected from the rats one (1M) and six (6M) months after splenic surgery, and alanine aminotransferase (ALT) concentrations were measured. Data are shown as means ± SEM (n = 6 rats per group). *P < 0.01 versus Cont, †
P < 0.05 versus Sham. (A color version of this figure is available in the online journal)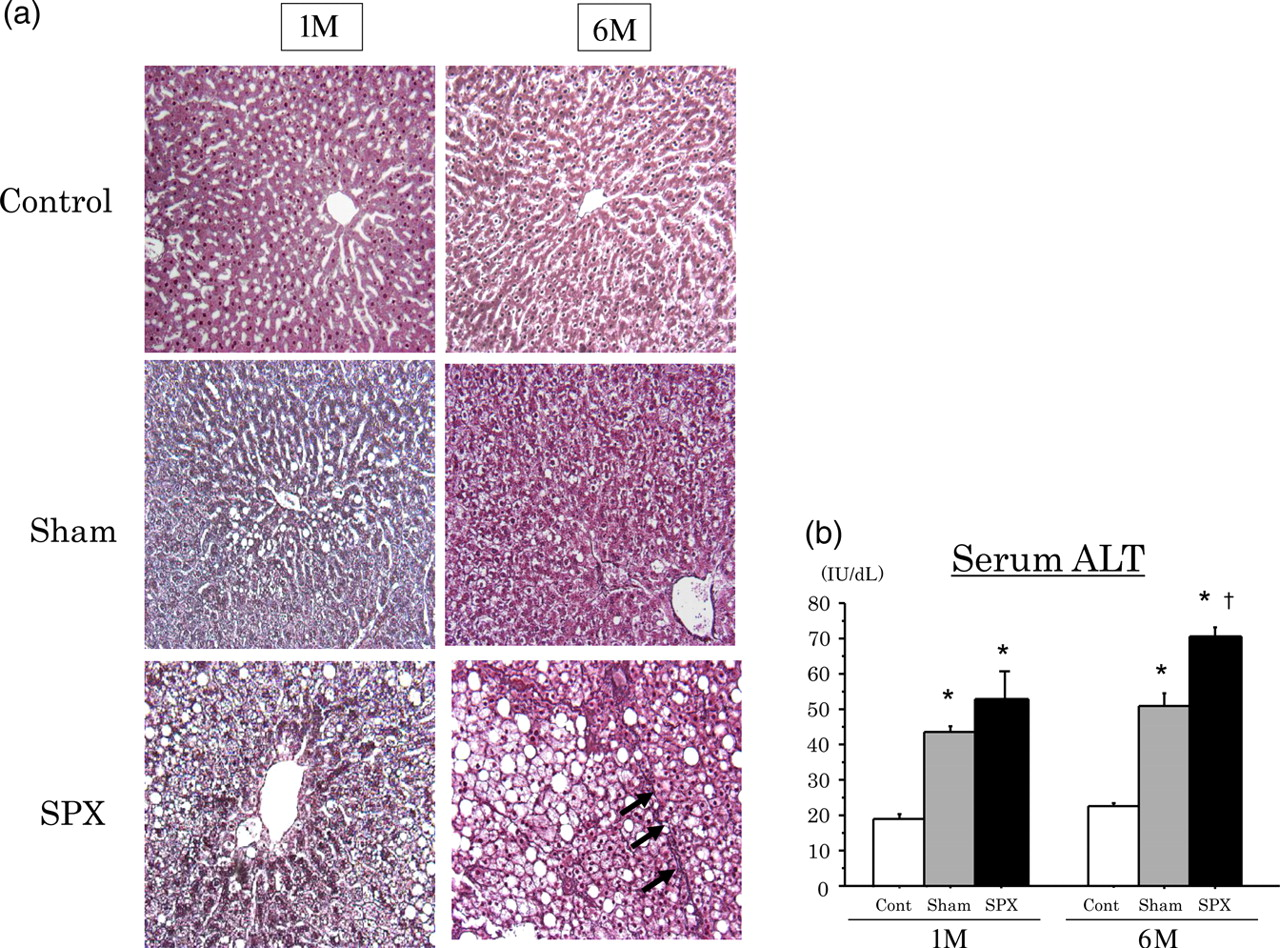
Changes in hepatic molecular markers related to lipid mobilization in SPX rats
To look for factors causing the acceleration of hepatic steatosis after SPX, we measured protein expression levels of the molecular markers related to fatty acid mobilization in the liver (Figure 6). Expression of CPT-1, a marker of mitochondrial β-oxidation, was significantly suppressed in 1M-SPX and 6M-SPX rats, compared with the respective sham controls (P < 0.05 for each) (Figure 6a). In contrast, the expression of liver SREBP-1 (Figure 6b) and FAS (Figure 6c), markers of lipid synthesis, showed no marked change in 1M-SPX rats, but were lower in 6M-SPX rats compared with the respective sham controls (P < 0.05 for each).
A mitochondrial β-oxidation disorder caused by suppression of hepatic carnitine palmitoyltransferase-1 (CPT-1) expression develops in high-fat (HF)-diet-induced obese rats in the splenectomized (SPX) state. At one (1M) and six (6M) months after splenic surgery, hepatic protein concentrations of (a) CPT-1, (b) sterol regulatory element-binding protein-1 (SREBP-1) and (c) fatty acid synthetase (FAS) were assessed by Western blot (n = 6 rats per group). *P < 0.05 versus Cont, **P < 0.01 versus Cont, †
P < 0.05 versus Sham
Changes in the liver KC population in SPX rats
To examine the changes in liver KCs in SPX rats, we determined the number of KCs (Figure 7a) by immunohistochemistry and the CD68 protein concentration by Western blot (Figure 7b) in rat livers. Immunohistochemistry with an anti-CD68 antibody revealed a lower number of KCs and lower expression levels of the CD68 protein in the livers from 1M-SPX rats compared with 1M-Sham rats (P < 0.05). In contrast, the number of CD68-positive KCs, assessed by immunohistochemistry, increased, and CD68 protein concentrations were significantly higher in the livers from 6M-SPX rats than 6M-Sham rats (P < 0.05).
Altered Kupffer cell numbers and subsequent hepatic proinflammatory cytokine production in high-fat (HF)-diet-induced obese rats according to time after removal of the spleen. (a) Representative immunohistochemical staining using an antibody against the specific macrophage marker CD68 in livers of rats one (1M) and six (6M) months after splenic surgery (×400). (b) Hepatic protein expression in 1M and 6M rats measured by Western blot (n = 6 rats per group). (c, d) Tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) concentrations at (c) one (1M) and (d) six (6M) months after splenic surgery (n = 6 rats per group). *P < 0.05 versus Cont, **P < 0.01 versus Cont, †
P < 0.05 versus Sham. (A color version of this figure is available in the online journal)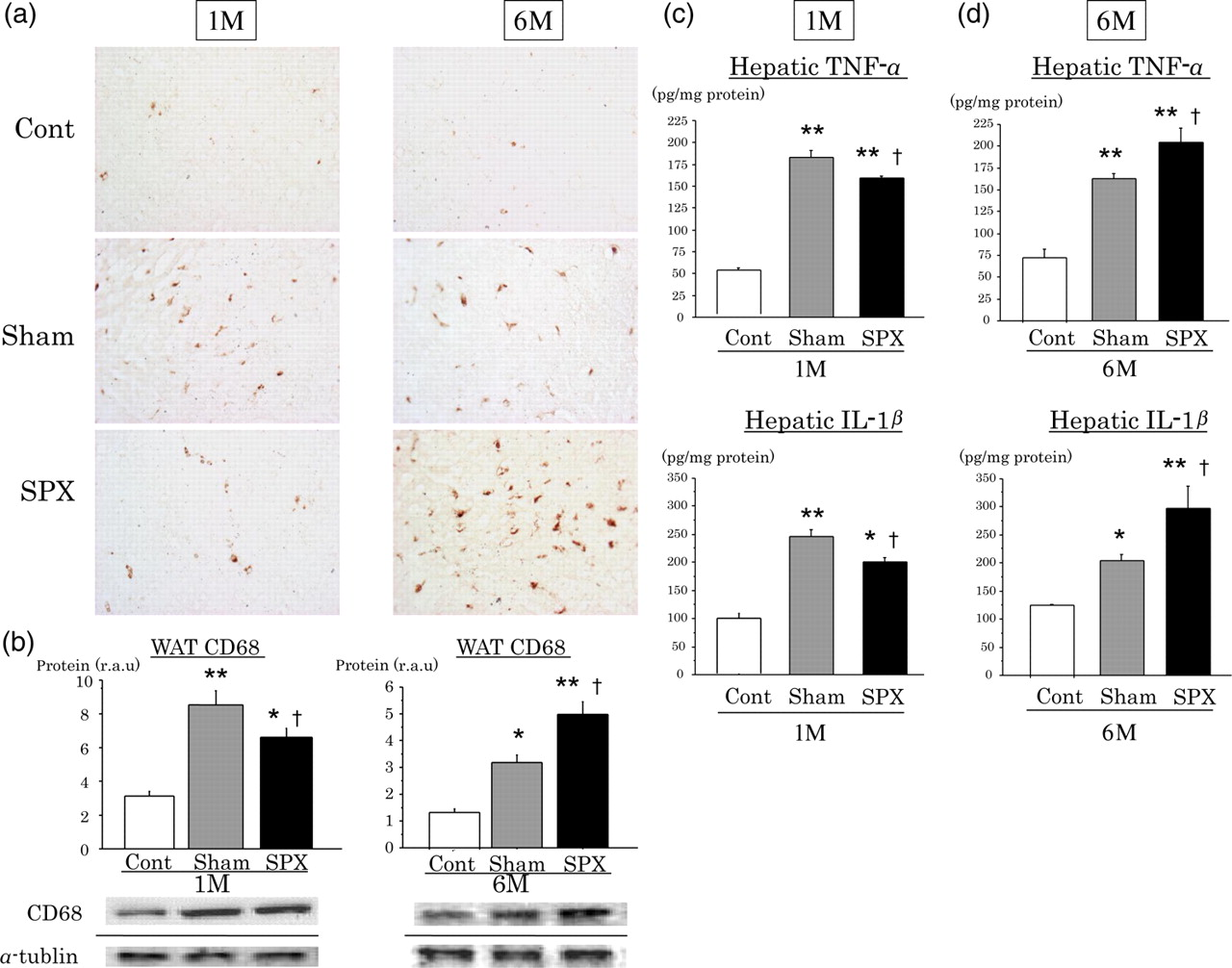
Changes in hepatic proinflammatory cytokine content in SPX rats
Next, the concentrations of the proinflammatory cytokines, such as TNF-α and IL1-β, were measured in the livers of the rats (Figures 7c and d). Proinflammatory cytokine concentrations were lower in the livers of 1M-SPX than 1M-Sham rat livers (P < 0.05 for each) (Figure 7c), whereas they were higher in 6M-SPX rats, compared with 6M-Sham rats (P < 0.05 for each) (Figure 7d). The change in cytokine concentrations occurred with the same time course, between one and six months after SPX, as the hepatic inflammatory changes observed by liver histology and the increase in the KC population.
Discussion
Previous studies reported no difference in mortality after LPS or Escherichia coli injections in mice with and without splenectomy. 28 In this study, all rats, including Sham and SPX rats fed on HF diets, survived for the one- or six-month study period (100% survival), indicating that SPX does not worsen survival in HF-fed rats during a natural time course after surgery.
It is now widely accepted that the immune system displays a close relationship with the development of obesity. Evidence over the last decade has indicated that the number of macrophages in WAT is elevated in obese subjects and that the activation of subsequent inflammatory pathways is involved in the metabolic complications of obesity. 3,5,9
The spleen is the largest lymphoid organ, and plays an important role in host immune functions. Although several studies have shown that the macrophage response, including that involving hepatic KCs, to bacterial infection is affected by spleen removal, only a few have demonstrated the involvement of the spleen in metabolic disorders such as fatty liver disease. Some previous reports demonstrated a relationship between the spleen and lipid metabolism, indicating that SPX-induced dyslipidemia was caused by a loss of splenic macrophages, which play a role in lipid storage. 32 In this study, in addition to the worsening of dyslipidemia after SPX, we clearly demonstrated that intrahepatic fat accumulation caused by HF feeding was increased in the splenectomized state. Here, we focused on fatty acid metabolism in the liver to identify factors important for the progression of hepatic steatosis after removal of the spleen.
The main transcriptional factor controlling lipid synthesis in the liver is SREBP. The SREBP-1c isoform is most relevant in rodent and human livers, and is generally recognized as a key modulator of the enzymes involved in fatty acid and TG synthesis 33 SERBP-1c is regulated by various factors, including TNF-α. 15 CPT-1 is a key enzyme controlling mitochondrial β-oxidation, a crucial process in fatty acid metabolism in the liver. CPT-1 is localized in the outer mitochondrial membrane with its active site exposed to the cytosolic space of the mitochondrion. 34,35 Feeding with an HF diet leads to a marked increase in hepatic CPT-1 gene expression in rats. 36 Our results showed that liver CPT-1 expression was initially suppressed at one month after removal of the spleen and that FAS and SREBP-1c protein expression levels were suppressed later. We suggest that the initial reduction in hepatic CPT-1 expression may induce fat accumulation through impairment of fatty acid β-oxidation. However, these data are insufficient to identify the mechanism of liver CPT-1 suppression in SPX rats; so, further investigations are needed to clarify the mechanism using other methods, such as knocking down of CPT-1 using siRNA.
The present study also showed that hepatic inflammation was accelerated in SPX rats, as indicated by an increased number of KCs, invasion of inflammatory cells and a dependent increase in intrahepatic proinflammatory cytokine production. KCs have previously been demonstrated to affect lipid metabolism and lipid storage in hepatocytes. 37 Conversely, KC depletion was shown to result in a reduction of liver steatosis in mice chronically fed on an HF diet, indicating a stimulatory effect of KCs on hepatic TG storage. 12,38 Considering that expression of liver lipogenetic markers, such as SREBP-1 and FAS, was also suppressed in the 6M-SPX group, the most likely factor contributing to liver steatosis in the chronic phase after SPX was the development of hepatic inflammation, including increased KCs and proinflammatory cytokine production.
In addition to the peripheral dyslipidemia caused by SPX, liver steatosis is influenced by different factors during the acute (one month) versus the chronic (six months) phases after spleen removal. An acceleration of hepatic steatosis in the 1M-SPX group was induced by a β-oxidation disorder due to the suppression of liver CPT-1. In the 6M-SPX group, further lipid accumulation in the liver may have been regulated not only by the suppression of hepatic CPT-1, but also the increase in KC number and subsequent proinflammatory cytokine production.
Previous reports have demonstrated that removal of the spleen alters the function of macrophages, including KCs, in organs such as the liver. 25,28,39 KCs are derived from circulating monocytes. In experimental animals, spleen removal reduced the number of KCs and suppressed KC activity, with a decrease in hepatic IFN-γ production 29 after LPS stimulation in the acute phase, although these parameters were increased during the late phase. 28 Splenic macrophages infiltrate the liver and become hepatic KCs after an inflammatory stimulus. 40,41 Thus, it is conceivable that the population of KCs is smaller in livers from splenectomized rats versus control rats during the early phase after removal of the spleen. Our results are consistent with previous reports and indicate that the initial changes in the liver during the early period after spleen removal were caused by the loss of the spleen as an immune organ that provides mononuclear phagocytic cells and cytokines. However, during the later chronic period, the liver may begin to play a role as an immune organ, compensating for the loss of the immunological function of the spleen. With these considerations, we suggest that the continuous activation of KCs for an extended period after removal of the spleen leads to the production of proinflammatory cytokines, inducing the development of steatohepatitis. Furthermore, these cytokines activate KCs via a positive feedback mechanism. However, these data are insufficient to confirm the influence of this increase in KC numbers after SPX on the progression to steatohepatitis. Huang et al. 12 demonstrated that depletion of liver KCs by administration of gadolinium chloride (GdCl3), a selective toxicant for KCs, prevented development of hepatic steatosis and insulin resistance in HF and high-sucrose diet-induced obese rats. Thus, further investigations are needed to clarify this relationship using KC-depletion techniques, such as liposome-encapsulated Cl2MDP and GdCl3.
In contrast, during the early phase when hepatic inflammatory changes were not yet observed, inflammation in WAT, evaluated by an increase in the number of inflammatory cells, macrophages and tissue proinflammatory cytokines and high levels of serum hsCRP, was already induced by loss of the spleen. We speculate that the splenectomized state led first to inflammatory changes in WAT, which may have led to worsening of hepatic steatosis and development of inflammatory changes in the liver. This progression is similar to that of metabolic syndrome in humans.
Thus, our results demonstrate that the liver changes with time in the splenectomized state. Interestingly, the process of steatohepatitis development after removal of the spleen occurred in two phases: an early phase with the worsening of steatosis and a later phase with the development of hepatic inflammation and fibrosis. This series of effects may cause a progression from simple steatosis to the development of fibrosing steatohepatitis that was similar to the liver histological findings in human NASH. The current popular theory of NASH pathogenesis is the ‘two-hit’ hypothesis. 42 According to this theory, the first hit leads to hepatic fat accumulation, and the second leads to steatohepatitis, in which proinflammatory cytokines, such as TNF-α, as well as oxidative stress and subsequent lipid peroxidation, contribute to NASH pathogenesis. In this study, the loss of the immunological function of the spleen seemed to act as the second stimulus to induce the resulting steatohepatitis.
In conclusion, our results indicate that loss of the spleen may induce the progression of simple hepatic steatosis to steatohepatitis in obese subjects. That is, preservation of spleen function may be an important factor regulating the progression of fatty liver disease. Although we were unable to identify the mechanisms by which the asplenic state modulated immune cell function and the metabolic system in various compartments of the body, this study provides a novel insight into the pathophysiological role of the spleen in the functional linkage between adiposity and hepatic inflammation.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by Grants-in-Aid from the Japanese Ministry of Education, Science and Culture and Research Grants for Intractable Diseases from the Japanese Ministry of Health and Welfare.