
Abstract
Cardiovascular disease is characterized by impaired exercise capacity and endothelial dysfunction, i.e. reduced bioavailability of nitric oxide (NO). Phosphodiesterase-5 (PDE5) inhibition is a promising vasodilator therapy, but its effects on pulmonary and systemic hemodynamic responses to exercise in the absence, and particularly in the presence, of endothelial dysfunction have not been studied. We investigated the effects of PDE5 inhibitor EMD360527 in chronically instrumented swine at rest and during exercise with and without NO synthase inhibition (Nω
-nitro-
Introduction
Endothelial dysfunction is a prominent feature of cardiovascular disease, correlates inversely with exercise capacity and is characterized by a reduced nitric oxide (NO) bioavailability. 1,2 Treatment of various forms of cardiovascular disease therefore often involves NO supplementation; for instance, nitroglycerin is used in angina pectoris and inhaled NO is used to alleviate the symptoms of pulmonary hypertension. However, these therapies usually have a short duration in action and desensitization may occur. In recent years, phosphodiesterase (PDE)-5 inhibition has emerged as a promising novel vasodilator therapy. 3–6 The vasodilator influence of PDE5 inhibition is achieved by prolonging the half-life of cyclic guanosine monophosphate (cGMP), which is not only the main second messenger in the NO pathway but also in various other pathways, like natriuretic peptide and carbon monoxide signalling. 7 Thus, PDE5 inhibition may be a way to activate the vasodilator pathway of NO in a sustained manner, without evoking desensitization.
PDE5 expression is not uniform in the cardiovascular system. PDE5 is abundantly expressed in lung vascular smooth muscle 8 with less expression in the systemic vasculature. Since PDE5 inhibition may provide a powerful tool to selectively reduce pulmonary vascular resistance (PVR), while leaving systemic vascular resistance (SVR) unperturbed, 6 it has been used as therapy in patients with pulmonary hypertension, with minimal systemic effects. 4,9,10 However, some PDE5 inhibitors appear to be more selective for the pulmonary circulation than others, which may depend, in part, on the dosage used. 11,12 Therefore, the first aim of the present study was to assess the vasodilator effects of various dosages of the novel PDE5 inhibitor EMD360527 13 and to assess its selectivity in the pulmonary versus the systemic vascular bed of chronically instrumented, awake resting swine.
The pulmonary circulation is a low-pressure system due to a low resistance. As a result, it has a reduced capacity for additional vasodilation as compared with the systemic circulation, which is especially notable during exercise, when the increased cardiac output (CO) causes a marked increase in pulmonary artery pressure (PAP). 14,15 Since exercise can lead to exaggerated increases in pulmonary pressure 16 and/or a paradoxical increase in PVR in patients with pulmonary hypertension, 17 which contributes to the limited exercise capacity of these patients, 18 it is important to know the efficacy of vasodilator therapy not only under resting conditions but also during exercise. We have previously reported that NO contributes to exercise-induced pulmonary vasodilation. 19 Since NO is considered as one of the major sources of cGMP, 20 we hypothesized that the effect of PDE5 inhibition increases during exercise and depends on the presence of NO. Consequently, the second aim of the present study was to investigate the pulmonary and systemic vasodilator effect of PDE5 inhibition during exercise.
Since cardiovascular disease, and pulmonary hypertension in particular, have been proposed to be associated with endothelial dysfunction and reduced endothelial NO synthase (eNOS) activity, 21–23 it is important to determine to what extent the efficacy of PDE5 inhibitors depends on the activity of eNOS. Consequently, the third aim of the study was to evaluate the vasodilator effect of PDE5 inhibition in the presence of NOS inhibition. Since our results indicated a paradoxical increase in the pulmonary and systemic vasodilator effects of PDE5 inhibition in the presence of NOS inhibition, we hypothesized that this increased effect was due to sensitization of the NO–cGMP pathway, and tested this hypothesis by investigating the pulmonary and systemic vasodilator responses to the NO donor sodium nitroprusside (SNP) and the cGMP analog 8-bromo-cGMP.
Methods
Studies were performed in accordance with the ‘Guiding Principles in the Care and Use of Laboratory Animals’ as approved by the Council of the American Physiological Society, and with approval of the Animal Care Committee of the Erasmus MC Rotterdam. Twenty-five crossbred Landrace × Yorkshire swine of either sex (2–3 months old) were included in the study. Daily adaptation of animals to laboratory conditions started one week before surgery and continued during the first week after surgery. Adaptation included running on the treadmill for 15 min at 2–3 Kmh−1.
Surgical procedures
Swine were sedated with ketamine (30 mg kg−1 intramuscular), anesthetized with thiopental (10 mg kg−1 intravenous), intubated and ventilated with a mixture of O2 and N2O (1:2) to which 0.2–1% (vol vol−1) isoflurane was added. 19,24,25 Anesthesia was maintained with midazolam (2 mg kg−1 + 1 mg kg−1 per hour intravenous) and fentanyl (10 μg kg−1 per hour intravenous) and the depth of anesthesia was checked regularly using a pain stimulus (toe pinch). Under sterile conditions, the chest was opened via the fourth left intercostal space and a fluid-filled polyvinylchloride catheter was inserted into the aortic arch and pulmonary artery for blood pressure measurement (Combitrans® pressure transducers; Braun, Melsungen, Germany), blood sampling and infusion of drugs. An electromagnetic flow probe (14–15 mm; Skalar, Delft, The Netherlands) was positioned around the ascending aorta for measurement of CO. A polyvinylchloride catheter was also inserted into the left atrium to measure pressure. Catheters were tunnelled to the back and animals were allowed to recover, receiving analgesia (0.3 mg buprenorphine intramuscular) for two days and antibiotic prophylaxis (25 mg kg−1 amoxicillin and 5 mg kg−1 gentamycin intravenous) for five days. Studies were performed 1–2 weeks after surgery. After completing all experimental protocols, animals were killed by an intravenous overdose of pentobarbitone sodium.
Experimental protocols
Schematic representation of the overlap of swine used in the different protocols
PDE5i, PDE5 inhibition; Ex, exercise; NLA, Nω
-nitro-
Numbers in bold indicate total number of animals in each protocol
Dose response of EMD360527 in resting swine
With swine (n = 10) resting quietly in a cage, the vasodilator responses in the systemic and pulmonary circulation to the PDE5 inhibitor EMD360527 13 were studied. EMD360527 demonstrates at least 45-fold selectivity for PDE5 (IC50 = 0.007 μmol L−1) compared with PDE6 (IC50 = 0.32 μmol L−1), 94-fold selectivity for PDE1 (IC50 = 0.66 μmol L−1), 137-fold selectivity for PDE10 (IC50 = 0.96 μmol L−1) and >1400-fold selectivity for PDE2, PDE3, PDE4 and PDE7 (all IC50 > 10 μmol L−1) (Dr Beier, unpublished observations). EMD360527 was administered in consecutive 15-min infusions of 10, 30, 100 and 300 μg kg−1 min−1 intravenously. At the end of each infusion period, heart rate, CO, and aortic, pulmonary arterial and left atrial blood pressures were recorded. Arterial blood samples were subsequently withdrawn for determination of plasma concentrations of EMD360527. EMD360527 was dissolved in a final concentration of 100 μg kg−1 mL−1 in a 1:1 (vol vol−1) mixture of saline and NaHCO3 (4.2%). We have previously shown that saline infusions do not affect hemodynamic variables in awake resting swine. 26
Effect of PDE5 inhibition during treadmill exercise
On a different day, hemodynamic measurements as well as blood samples were obtained, and swine (n = 11) were subjected to a five-stage exercise protocol (1–5 km h−1) on a motor-driven treadmill. Hemodynamic variables were continuously recorded and blood samples collected during the last 60 s of each three-minute exercise stage, at a time when hemodynamics had reached a steady state. After 90 min of rest, the PDE5 inhibitor EMD360527 was infused continuously in a dose of 300 μg kg−1 min−1 intravenous, and 10 min after starting the infusion, the exercise protocol was repeated. We have previously observed excellent reproducibility of two consecutive control exercise protocols. 19,25
Effects of PDE5 inhibition during treadmill exercise in the presence of NOS inhibition
On a different day, the NOS inhibitor Nω
-nitro-
Effects of sodium nitroprusside in the presence of NOS inhibition
Resting swine (n = 7) received consecutive 10-min intravenous infusions of the NO donor SNP in doses of 0.5, 1.0, 2.0 and 3.0 μg kg − 1 min − 1 before and after NLA (20 mg kg − 1 min − 1 intravenous) and systemic and pulmonary hemodynamic responses were studied.
Effects of 8-bromo-cGMP in the presence of NOS inhibition
Resting swine (n = 5) received consecutive 10-min intravenous infusions of the stable cGMP analog 8-bromo-cGMP in doses of 3, 10 and 30 μg kg−1 min−1 27 before and after NLA (20 mg kg−1 min−1 intravenous), and systemic and pulmonary hemodynamic responses were studied.
Data analysis
Digital recording and offline analysis of hemodynamics have been described previously. 19,25 SVR was computed as mean aortic blood pressure (MAP) divided by CO. PVR was defined as mean PAP minus mean left atrial pressure (LAP) divided by CO.
Plasma concentrations of EMD360527 were determined using a high performance liquid chromatography (HPLC)-tandem mass spectrometry assay, involving an Agilent 1100 Series HPLC system with an HTC CTC PAL Autosampler coupled to the turboion spray source of a mass spectrometer of the API 4000 series from Applied Biosystems (Carlsbad, CA, USA). The plasma concentrations of EMD360527 were calculated by interpolation of the peak area ratios of analyte/internal standard versus the ratio of their nominal concentrations into the regression line obtained from the calibration standards. The calibration curve used covered a concentration range of 0.04–46.06 μmol L−1.
Statistical analysis
The effects of EMD360527 in resting swine were assessed using analysis of variance (ANOVA) for repeated measures followed by post hoc testing using Scheffe's test. The effects of EMD360527 during exercise were analyzed using two-way ANOVA (exercise level × EMD360527) for repeated measures. Post hoc testing for exercise and drug effects was performed using Scheffe's test. The effect of NOS inhibition on the responses to EMD360527 during exercise were tested using three-way ANOVA (exercise level × EMD360527 × NLA) for repeated measures. Linear regression analysis was used to test the dependency of the effect of EMD360527 on the initial level of pressure and resistance in the pulmonary and systemic vascular bed. Finally, the effect of NOS inhibition on the vasodilator responses to SNP and 8-bromo-cGMP were assessed by two-way ANOVA (dose×NLA). Statistical significance was accepted when P < 0.05. Data are presented as mean ± SEM.
Results
Dose–response of EMD360527 in resting swine
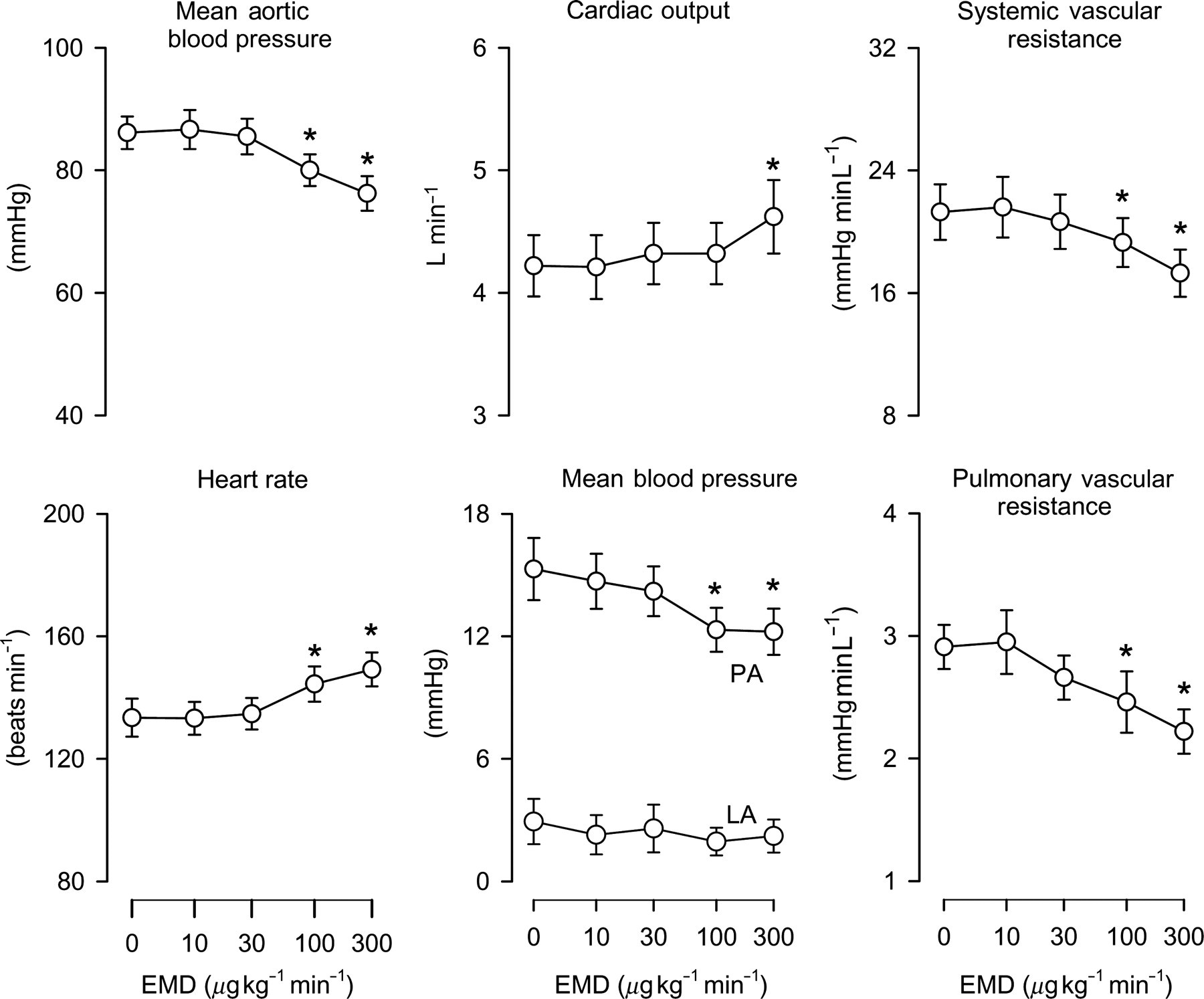
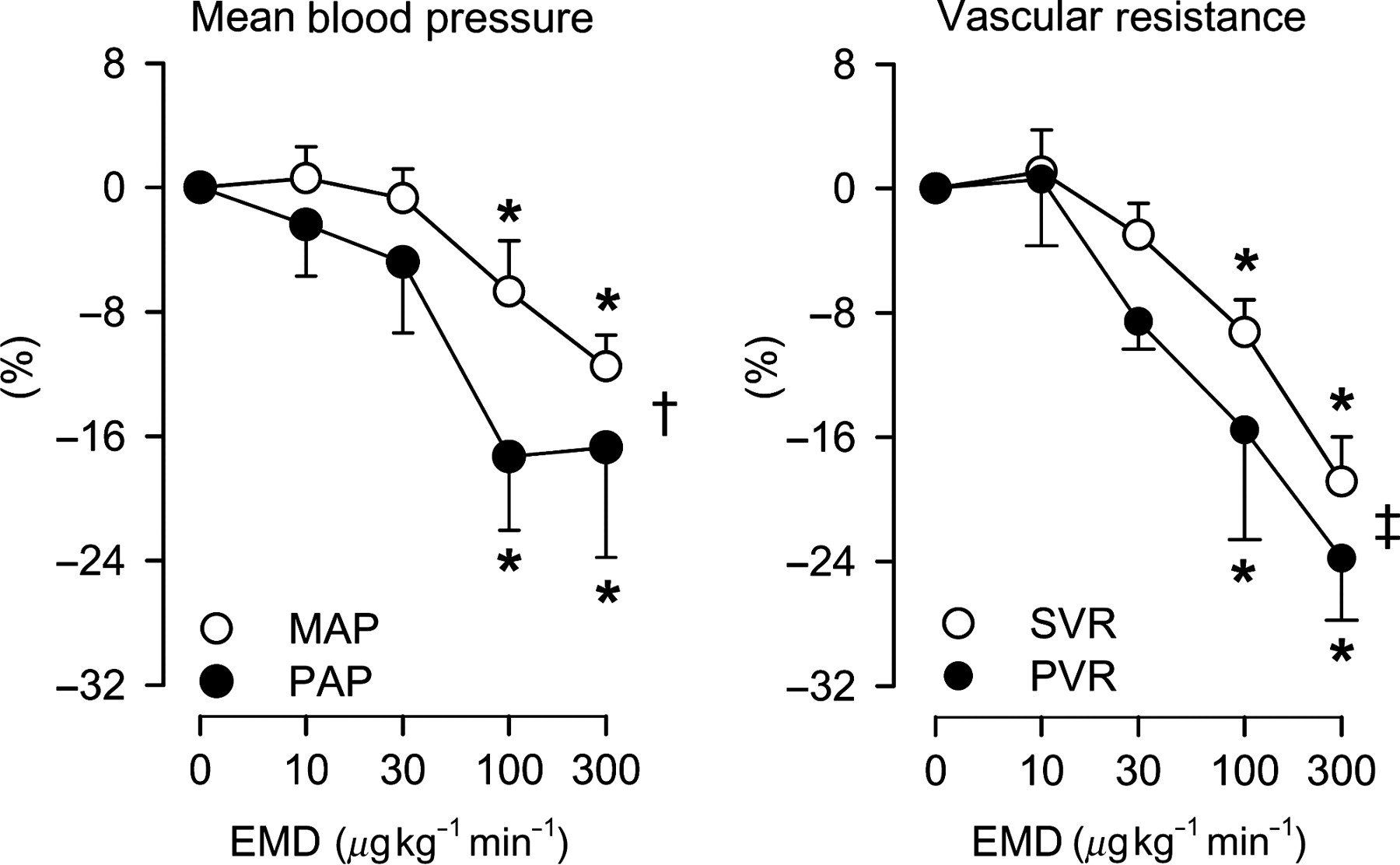
Consecutive 15-min infusions of 10, 30, 100 and 300 μg kg−1 min−1 intravenous EMD360527 resulted in dose-dependent increases in its plasma concentrations of 0.35 ± 0.04, 1.26 ± 0.12, 4.8 ± 0.5 and 15.5 ± 1.5 μmol L−1. EMD360527 caused a decrease in MAP of up to 12 ± 2%, which was due to a 19 ± 3% decrease in SVR, as CO increased by up to 10 ± 3% (Figure 1); the latter was the result of a (probably baroreflex-mediated) increase in heart rate. EMD360527 also caused a dose-dependent decrease in PAP of up to 17 ± 7%, which was due to a decrease in PVR of up to 24 ± 4%, as LAP was not affected and CO increased (Figure 1). The relative decreases in arterial pressure and vascular resistance in response to EMD360527 tended to be slightly larger in the pulmonary than in the systemic vascular bed (P < 0.05 for pressure, P = 0.053 for vascular resistance, Figure 2).
Hemodynamic effects of PDE5 inhibition in resting swine. Hemodynamic effect of consecutive intravenous infusions of PDE5 inhibitor EMD360527 (EMD) in 10 awake resting swine. PA, pulmonary artery; LA, left atrium; PDE5, phosphodiesterase-5. *P < 0.05 versus baseline (0 μg kg−1 min−1) Pulmonary versus systemic vasodilator effect of PDE5 inhibition in swine. Relative effect of PDE5 inhibitor EMD360527 (EMD) on hemodynamic parameters in awake swine. MAP, mean arterial pressure; PAP, pulmonary artery pressure; SVR, systemic vascular resistance; PVR, pulmonary vascular resistance; PDE5, phosphodiesterase-5. *P < 0.05 versus baseline (0 μg kg−1 min−1); †
P < 0.05 by two-way analysis of variance (ANOVA), effect EMD on MAP versus PAP; ‡
P = 0.053 by two-way ANOVA, effect EMD on SVR versus PVR for dosages 30–300 μg kg−1 min−1
Effects of PDE5 inhibition during treadmill exercise
Hemodynamics of swine at rest and during exercise before and after administration of EMD360527 and NLA
HR, heart rate; MAP, mean arterial pressure; CO, cardiac output; SVR, systemic vascular resistance; PAP, mean pulmonary arterial pressure; LAP, mean left atrial pressure; PVR, pulmonary vascular resistance; NLA, NO synthase inhibitor Nω
- nitro-
In accordance with its effects in the dose–response protocol, EMD360527, in a dose of 300 μg kg−1 min−1, resulted in a significant decrease in systemic and pulmonary pressures and vascular resistances (Table 2). During the subsequent exercise protocol, the vasodilator effect of PDE5 inhibition on the pulmonary vasculature was not significantly altered, although a trend towards an increased effect (P = 0.08) could be observed, while the vasodilator effect in the systemic vasculature decreased (P < 0.05 by ANOVA), compared with the effects of the PDE5 inhibition during resting conditions (Figure 3).
Effect of PDE5 inhibition in swine during exercise in absence and presence of NLA. Effects of the PDE5 inhibitor EMD360527 alone and after prior administration of the NO synthase inhibitor NLA in swine on mean arterial blood pressure (MAP, n = 11 without NLA and n = 9 with NLA), systemic vascular resistance (SVR, n = 9 without NLA and n = 6 with NLA), pulmonary artery pressure (PAP, n = 11 without NLA and n = 9 with NLA) and pulmonary vascular resistance (PVR, n = 9 without NLA and n = 6 with NLA) at rest (0L, lying and 0S, standing) and during exercise (1–5 km h−1) are shown. Dot inside symbol P < 0.05, effect of EMD; *P < 0.05 by ANOVA, effect of EMD is different during exercise; **P = 0.08 by ANOVA, effect of EMD is different during exercise; †
P < 0.05 by ANOVA, effect of EMD is different after NLA. PDE5, phosphodiesterase-5; NLA, Nω
-nitro-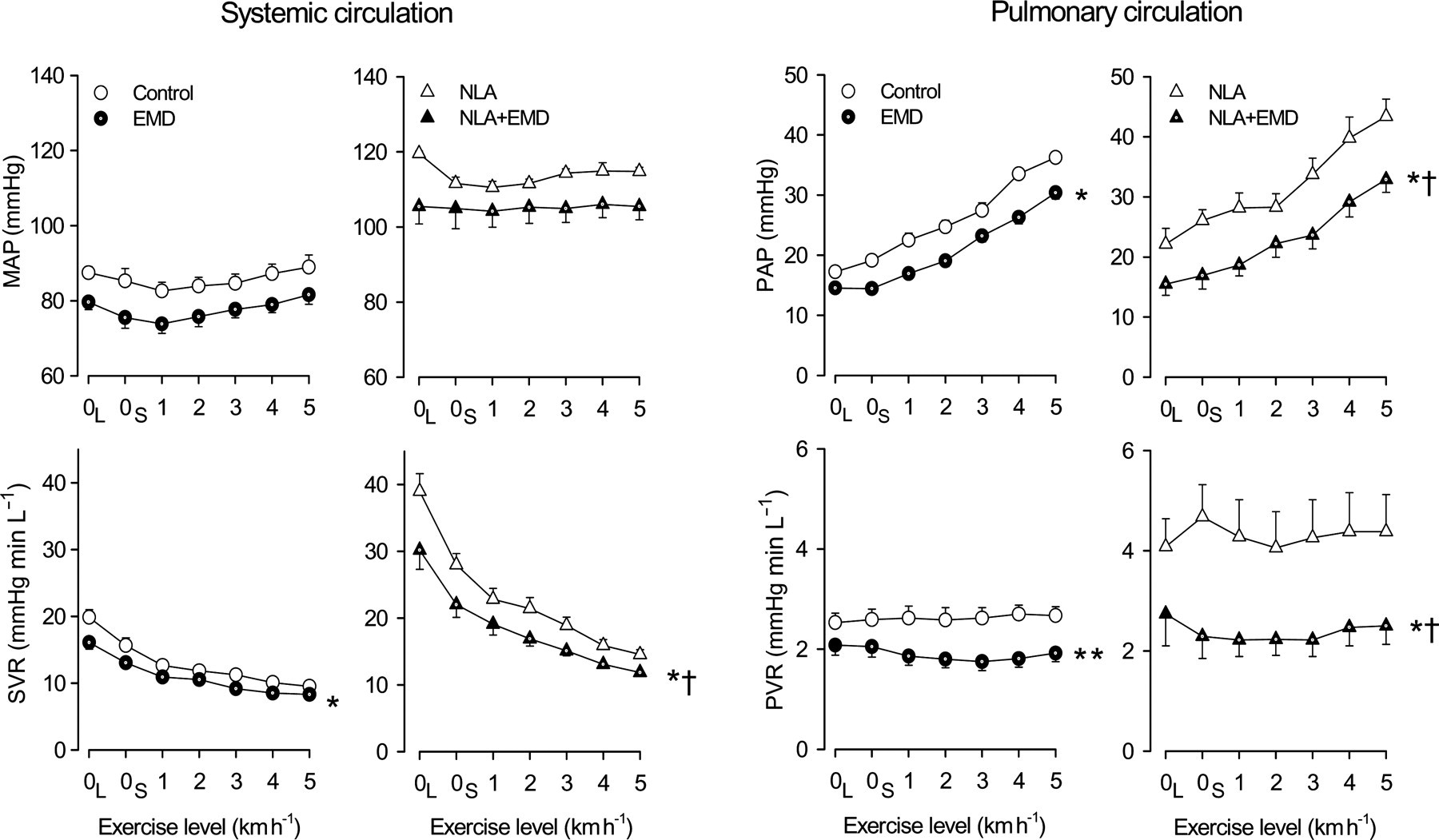
Effects of PDE5 inhibition during exercise in the presence of NOS inhibition
Administration of the NOS inhibitor NLA resulted in marked increases in aortic (37 ± 2%) and pulmonary arterial (41 ± 14%) blood pressures, which were the result of a 94 ± 9 and 89 ± 23% increase in SVR and PVR, respectively (Table 2). The vasoconstrictor responses to NLA were similar at rest and during exercise.
Subsequent infusion of EMD360527 caused vasodilator responses in the systemic and pulmonary circulations that were enhanced compared with the vasodilator responses to PDE5 inhibition in the absence of NLA (Figure 3). During exercise, the pulmonary vasodilation by EMD360527 was maintained, while the systemic vasodilation by EMD360527 decreased, compared with the vasodilation by EMD365027 during resting conditions.
The magnitude of the vasodilator responses of individual animals to PDE5 inhibition in the systemic and pulmonary vascular beds, both at rest and during exercise and both in the absence or presence of NLA, correlated well with the baseline values obtained just prior to administration of EMD360527. Conversely, the depressor responses (decreases in pulmonary and systemic blood pressure) correlated much less, reaching statistical significance only in the pulmonary circulation (Figure 4).
Effect of PDE5 inhibition with and without inhibition of NO synthase. Responses of mean arterial pressure (MAP), systemic vascular resistance (SVR), pulmonary arterial pressure (PAP) and pulmonary vascular resistance (PVR) to PDE5 inhibitor EMD360527 (300 μg kg−1 min−1) in the absence (open symbols) and presence (closed symbols) of inhibition of NO synthase plotted as a function of the basal (=pre-EMD360527) levels of each variable. Each data point represents a response of an individual animal at rest or during exercise. Linear regression lines are shown as solid lines. PDE5, phosphodiesterase-5; NO, nitric oxide
Effects of NOS inhibition on the vasodilator responses to SNP and 8-bromo-cGMP
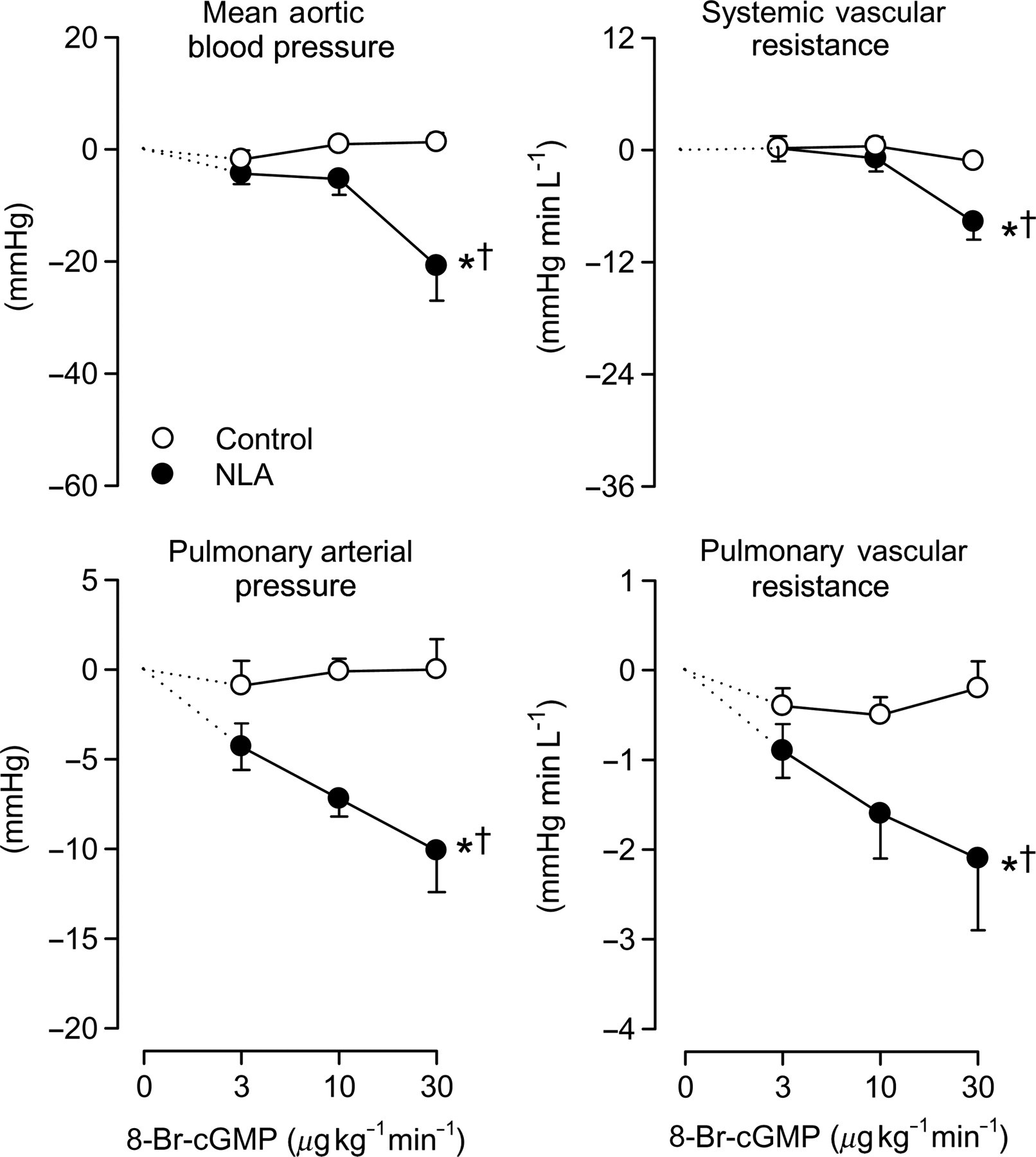
To investigate whether the vasculature becomes more sensitive to the vasodilator influence of NO/cGMP after inhibition of NOS, the NO donor SNP and the stable cGMP analog 8-bromo-cGMP were infused intravenously under control conditions and after NOS inhibition with NLA. SNP, in doses that elicited no vasodilator responses in systemic and pulmonary circulation under control conditions, did induce dose-dependent vasodilation in both the systemic and pulmonary vascular bed following NLA (Figure 5). Similarly, the systemic and particularly pulmonary vasodilator responses to 8-bromo-cGMP (which were negligible under control conditions) were enhanced after pretreatment with NLA (Figure 6). These data suggest that constriction of the vasculature renders it more sensitive to the vasodilator actions of NO and cGMP.
Effect of sodium nitroprusside with and without inhibition of NO synthase. Responses of mean arterial pressure, systemic vascular resistance, pulmonary arterial pressure and pulmonary vascular resistance to different doses of sodium nitroprusside (SNP) in the absence (open symbols) and presence (closed symbols) of inhibition of NO synthase (NLA). *P < 0.05 by ANOVA, effect of SNP; †
P < 0.05 by ANOVA, effect of SNP different after NLA. NO, nitric oxide; NLA, Nω
-nitro- Effect of 8-bromo-cGMP with and without inhibition of NO synthase. Responses of mean arterial pressure, systemic vascular resistance, pulmonary arterial pressure and pulmonary vascular resistance to different doses of 8-bromo-cGMP (8-br-cGMP) in the absence (open symbols) and presence (closed symbols) of inhibition of NO synthase (NLA). *P < 0.05 by ANOVA, effect of 8-br-cGMP; †
P < 0.05 by ANOVA, effect of 8-br-cGMP different after NLA. PDE5, phosphodiesterase-5; NLA, Nω
-nitro-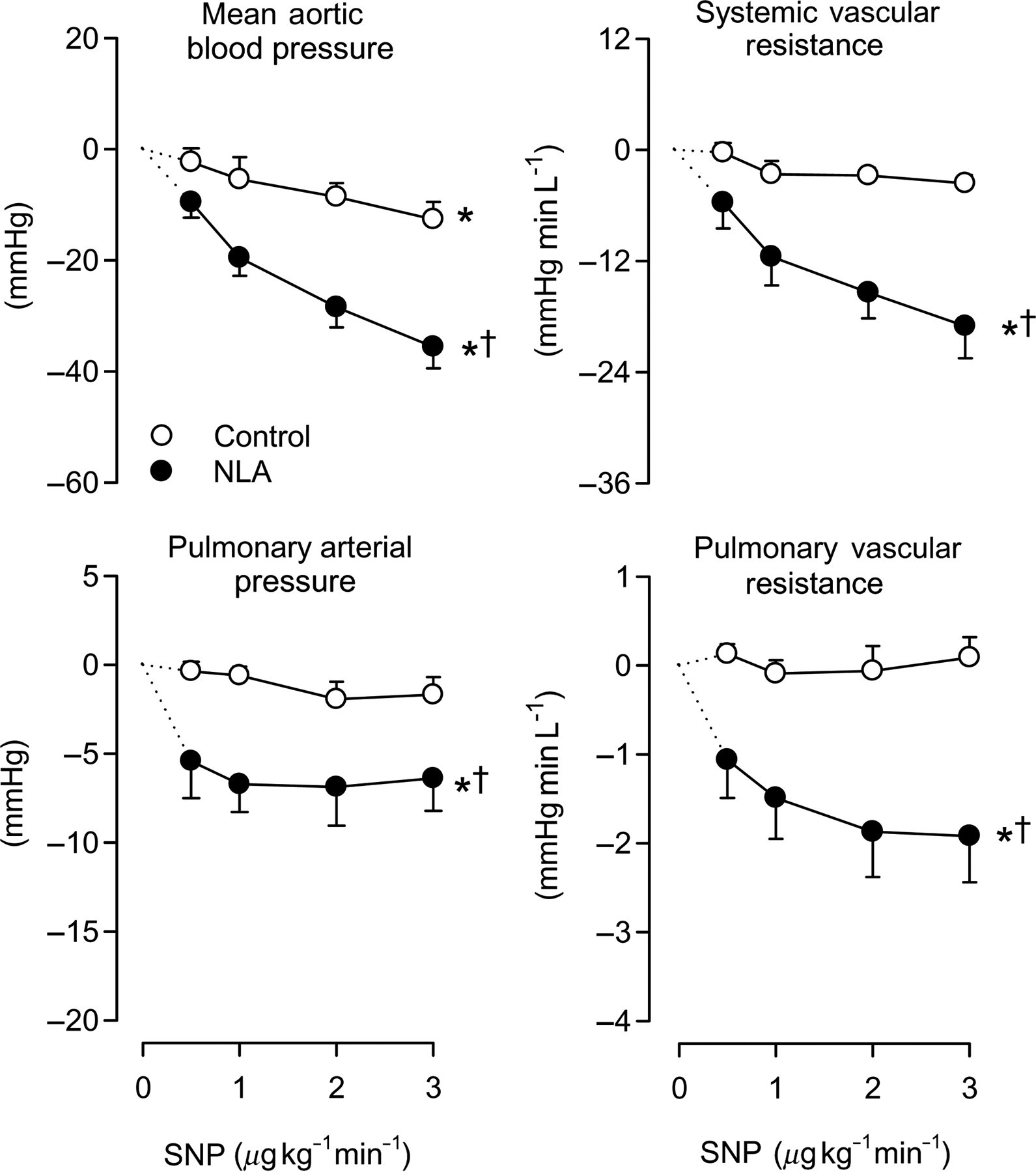
Discussion
The present study investigated the systemic and pulmonary vasodilator effects of PDE5 inhibition in a model of endothelial dysfunction at rest and during exercise. The main findings of the present study are that (i) the PDE5 inhibitor EMD360527 resulted in dose-dependent vasodilation in the pulmonary and systemic circulation of awake swine under resting conditions; (ii) the pulmonary circulation displayed slightly higher sensitivity to the vasodilator influence of EMD360527 than the systemic circulation; (iii) the vasodilator effect of PDE5 inhibition on PVR was well maintained during exercise, while its effect on SVR decreased slightly; (iv) inhibition of NOS increased the sensitivity to vasodilation via the NO–cGMP pathway; and (v) the vasodilator effect of PDE5 inhibition by EMD360527, in a dose of 300 μg kg−1 min−1, was enhanced after inhibition of eNOS at rest and during exercise in both pulmonary and systemic circulation. The implications of these findings will be discussed.
Effects of PDE5 inhibition on pulmonary and SVR in resting swine
Many vasoactive substances influence vascular resistance by increasing the production of the second messengers cyclic adenosine monophosphate (cAMP) and/or cGMP within vascular smooth muscle cells. These second messengers are short-lived because they are degraded by PDEs, allowing tight control of vascular tone. At least 11 different gene families of PDEs are currently known to exist in mammalian tissues but the tissue distribution of the PDE isoforms and their specificity for cAMP and cGMP varies. 28 Inhibition of PDE in vascular smooth muscle can provide a powerful tool to reduce vascular resistance, by prolonging the half-life of cAMP and/or cGMP. The PDE isoforms that are predominantly present in vascular smooth muscle are PDE1, 3, 4, 5, 7 and 9. 29,30 PDE1, PDE4 and PDE7 are cAMP-specific, while PDE3 has a 4–10 times higher affinity for cAMP than for cGMP; PDE5 and PDE9 are cGMP-specific. 28 However, PDE9 is abundantly expressed throughout the body, 28 whereas PDE5 expression is 10 times more abundant in the pulmonary vasculature as compared to the heart. 8 Thus, PDE5 inhibition may provide a tool to preferentially reduce PVR, while leaving SVR principally unchanged. 3 In addition to its direct vasodilator effect, PDE5 inhibition may also affect vascular tone by increasing sympathetic nerve activity. 31 However, we have previously shown that activation of the sympathetic nervous system (i.e. during exercise) has no net effect on the pulmonary and systemic vasculature as α-adrenergic constriction and β-adrenergic vasodilation are similar in magnitude 25 and neutralize each other. It is therefore unlikely that the effects of PDE5 inhibition in the present study can be explained through an altered activation of the sympathetic nervous system.
In the present study we found that the PDE5 inhibitor EMD360527 decreased both SVR and PVR. However, the pulmonary circulation appeared to be slightly more sensitive to EMD360527, as the relative decreases in PAP and PVR were greater than the relative decreases in MAP and SVR. These findings are in agreement with several other experimental studies in lambs with normal or acutely elevated PAP, 32,33 and clinical studies in patients with chronic pulmonary hypertension, which all showed that pulmonary pressure decreased more in response to PDE5 inhibition as compared with systemic arterial pressure. 4,9,10,17 The degree of pulmonary over systemic selectivity may depend on the particular PDE5 inhibitor and/or dosage that is used. 11,12 However, it cannot be excluded that the selectivity for the pulmonary bed reported in studies performed in animal models and patients with pulmonary hypertension is in part due to the elevated baseline levels of PAP and vascular tone (see below).
Effects of PDE5 inhibition on PVR during treadmill exercise
In agreement with previous observations from our laboratory, 34 we found that exercise had minimal effects on PVR so that, in parallel to the increase in CO, PAP almost doubled during exercise. The maintained PVR during exercise appears to be the result of a balanced increase in endothelin vasoconstrictor influence 24 and NO vasodilator influence, 19,34 demonstrating that pulmonary vasomotor tone is the result of an interplay between vasodilator and vasoconstrictor influences. 14 The pulmonary vasodilator effect of PDE5 inhibition observed under resting conditions was well maintained during exercise and actually tended to increase. Similarly, a recent study showed that the effect of PDE5 inhibition was enhanced during exercise in patients with pulmonary hypertension secondary to left ventricular dysfunction, although such an effect was not observed in those patients that had normal PAP. 17 Since the exercise capacity of patients with pulmonary hypertension is inversely correlated with PVR, 18 these findings may explain the PDE5-inhibitor-induced improvement in exercise capacity that is observed in patients with pulmonary hypertension. 12,35 Indeed, both short- 17 and long-term 36 improvement in exercise capacity in response to PDE5 inhibition correlated well with the reduction in PVR.
Effects of PDE5 inhibition on SVR during treadmill exercise
In the present study, we observed that PDE5 inhibition reduced aortic blood pressure and SVR both at rest and during exercise. However, the vasodilator response was not enhanced during exercise compared with resting conditions (the absolute decrease in SVR was actually slightly less during exercise compared with rest). These observations are in good agreement with findings in humans with left ventricular dysfunction that the modest systemic vasodilation induced by PDE5 inhibition under resting conditions was blunted during exercise. 17 Since the exercise-induced systemic vasodilation is principally accounted for by skeletal muscle vasodilation, these observations suggest that PDE5 does not limit exercise-induced vasodilation in skeletal muscle. Indeed, a recent study found that PDE5 inhibition had no effect on forearm blood flow of healthy humans, either at rest or immediately following handgrip exercise. 37 Taken together, these findings support the current concept that exercise-induced vasodilation in the systemic vascular bed is not critically dependent on NO/cGMP-mediated vasodilator influence 19,38 and is not limited by PDE5 activity (see refs 17,37 and the present study).
Effects of PDE5 inhibition on pulmonary and systemic resistance in the presence of NO inhibition
Both systemic 39,40 and pulmonary 21–23 hypertension are often accompanied by endothelial dysfunction, resulting in decreased eNOS activity, and consequently cGMP production may be decreased. To simulate this condition we administered NLA to block NOS and compared the effect of PDE5 inhibition before and after NLA. We found that the vasodilator effect of EMD360527 was larger at rest and during exercise after prior inhibition of NOS in both the systemic and the pulmonary circulation. These findings in awake resting swine, appear at odds with several studies in anesthetized and awake (resting) animals which reported that inhibition of NOS blunted the systemic and pulmonary vasodilator responses to PDE5 inhibition. Thus, NOS inhibition blunted the vasodilator effects of various PDE5 inhibitors in rat aorta 41 as well as in the systemic circulation of mice 42 and lambs. 32,33 Moreover, studies in the pulmonary circulation, in either anesthetized animals 43 or awake animals at rest, 32,33 also suggest that the effect of PDE5 inhibition with either zaprinast or sildenafil is blunted by prior inhibition of NOS. However, these studies compared pulmonary vasodilation induced by PDE5 inhibition after NOS inhibition with its effect at high PVR, obtained with another pulmonary vasoconstrictor (i.e. the thromboxane analog U46619). Since we showed in the present study that the initial level of PVR is a strong determinant of the degree of vasodilation produced by PDE5 inhibition (Figure 4), we propose that these disparate results may be due, at least in part, to differences in basal vascular tone. This concept is further supported by the observation that the effect of PDE5 inhibition is larger in lambs with acute pulmonary hypertension as compared with its effect at baseline. 32,43 Similarly, the pulmonary vasodilator response to PDE5 inhibition was larger in patients with pulmonary hypertension and higher PVR. 17 However, to assess if the magnitude of the vasodilator response to PDE5 inhibition is indeed dependent on basal tone, additional experiments are required in which the vasodilator response to PDE5 inhibition is compared at different levels of basal tone (e.g. in the presence of a vasoconstrictor such as endothelin).
The unperturbed vasodilator response to EMD360527 in the presence of NLA may be explained, at least in part, by alternative sources of NO and/or cGMP. Thus, it has been shown that NO, released from erythrocytes in response to a decrease in their O2 content, can contribute to regulation of both pulmonary and systemic vascular tone. 44 Moreover, a large number of other mediators cause vasodilation via stimulation of soluble guanylyl cyclase, including carbon monoxide 45 and hydrogen peroxide. 46 There is evidence that blockade of NO production may result in increased carbon monoxide 47 and hydrogen peroxide 46 production, which could act to compensate for the loss of NO-induced cGMP production. In addition, the natriuretic peptide family of proteins (consisting of atrial, brain and C-terminal natriuretic peptide [ANP, BNP and CNP]) cause increases in cGMP by activating particulate GC 48 and may exert an increased vasodilator influence after NOS inhibition. 49 Moreover, ANP-induced vasodilation is enhanced by PDE5 inhibition in the pulmonary but not systemic vasculature. 50 Finally, it has been suggested that, while over-expression of NOS results in desensitization of the NO/cGMP pathway, 51 inhibition of NOS results in supersensitization of the NO/cGMP pathway, thereby resulting in enhanced response to vasodilators acting via the NO/cGMP pathway. 52 This concept is supported by our observation that the pulmonary and systemic vasodilator responses to the NO donor nitroprusside, as well as the cGMP analog 8-bromo-cGMP, were enhanced by pretreatment with NLA and could, in part, explain the enhanced vasodilator response to PDE5 inhibition in the presence of NLA.
Conclusions
The PDE5 inhibitor EMD360527 produced pulmonary vasodilation in awake resting swine that was maintained during treadmill exercise. Inhibition of NOS enhanced the vasodilator response not only to nitroprusside and 8-bromo-cGMP, but also to EMD360527. Patients with severe pulmonary hypertension often have endothelial dysfunction and decreased NOS activity. 21–23 The observation that inhibition of PDE5 was enhanced following NOS inhibition and correlated well with the basal level of pulmonary resistance vessel tone, supports clinical reports that PDE5 inhibition is most effective in patients with the severest degree of pulmonary hypertension.
Footnotes
Acknowledgements
This study was further supported by a grant from Merck KGaA, Darmstadt, Germany, and grant 2000T042 (to DM) from the Netherlands Heart Foundation. Technical assistance of Joris Fintelman and Rowena Utberg is gratefully acknowledged.