
Abstract
This study was designed to determine the therapeutic effect of Agaricus bisporus lectins (ABL) by the regeneration of β-cells in mice following 70% partial pancreatectomy (PPx), and to explore the mechanisms of ABL-induced β-cell proliferation. Adult C57BL/6J mice were subjected to a 70% PPx operation or a sham operation, and mice received 10 mg/kg body weight of ABL or saline immediately after surgery. Blood glucose concentrations and insulin secretion levels were measured. To determine the growth rates of β-cells and duct cells, immunohistological analysis of pancreatic tissues was performed. Key cell cycle proteins and β-cell specific genes were measured by realtime polymerase chain reaction, Western blotting and immunohistological staining. In this study, a significant decrease in blood glucose concentrations, increase in glucose tolerance and expanded β-cell mass were observed in the ABL-treated mice. At the same time, after ABL treatment, increased β-cell proliferation rates were observed. Further studies on the expression of cyclin D1, cyclin D2 and Cdk4 demonstrated that these genes were significantly up-regulated in the ABL-treated mice. Meanwhile, Cdk4 activity was also enhanced. Moreover, the expression of PDX-1 (pancreatic and duodenal homeobox 1), Ngn3 (neurogenin 3), insulin, GLUT-1 (glucose transporter 1) and glucokinase was also increased in the ABL-treated mice. These findings demonstrate that ABL administration could partially reverse the impaired β-cell growth potential by regulating cell cycle proteins. Induction of islet β-cell proliferation by ABL suggests the therapeutic potential in preventing and/or treating diabetes.
Introduction
Hyperglycemia in both type 1 and type 2 diabetes, which causes considerable morbidity, is due to insufficient insulin secretion to meet body requirement. Defective insulin secretion in both forms of diabetes is caused in part by a loss of β-cell mass. 1,2 Although transplantation with donor islets has achieved some success in clinical trials since the application of the Edmonton protocol, pancreas and islet transplantation has limited applicability in the treatment of diabetes owing to the shortage of organ donors. Due to the vital importance of β-cell mass in the pathogenesis of diabetes, an alternative and promising approach is the stimulation of endogenous β-cell regeneration to replace the deficit in β-cell mass in diabetic patients. It is widely acknowledged that β-cell mass is dynamic both in function and in mass to maintain the glycemic concentrations within a very narrow physiological range. 3 So in this study, we aimed to endogenously regenerate β-cells and expand β-cell mass. It has been proved that partial pancreatectomy (PPx), a classical model of regeneration in rodents, is followed by replication of pre-existing differentiated cells, hypertrophy of β-cells and the differentiation of whole new lobes of the pancreas. 4 Studies by Xu et al. 5 have demonstrated that partial duct ligation could induce neogenesis. Therefore we established partial pancreatectomized mice to study whether Agaricus bisporus lectin (ABL) therapy could induce islet regeneration.
ABL, two phytohemagglutinins with similar specificities for carbohydrate receptors, is originally extracted from A. bisporus. 6 Studies have demonstrated that A. bisporus stimulates insulin release in vitro, and that the stimulation is associated with increased conversion of proinsulin to insulin in the isolated islets of Langerhans of rats. 7,8 Further studies of lectins from A. bisporus and A. campestris have demonstrated their antihyperglycemic effect by stimulating insulin and glucagon release. 9 Recently, Jeong et al. 10 have reported that A. bisporus possesses both hypoglycemic and hypolipidemic activity in vivo. Herein, we reported for the first time the enhanced pancreatic β-cell proliferation capability induced by ABL in partial pancreatectomized mice, and we further studied the mechanisms of the ABL-stimulated β-cell proliferation.
Previous studies have demonstrated that replication is achieved by re-entry of the β-cell into the cell cycle and relies on proteins regulating the G1 phase. 11 Further studies have established that cyclin D1 and cyclin D2, G1/S cell-cycle regulators, are necessary for the postnatal expansion of β-cell mass. 12,13 Therefore, we focused our study on the expression of cell cycle proteins. Furthermore, Rane et al. 14 reported that loss of Cdk4 expression causes insulin-deficient diabetes and Cdk4 activation could induce β-cell hyperplasia in islets. Recently published data indicated that Cdk4 controls β-cell mass by recruiting quiescent cells to enter the cell cycle. 15 Thus we also assessed the abundance and activity of Cdk4. Meanwhile, β-cell-specific genes were also investigated in this study. Taken together, we explored the novel therapeutic approach of ABL by endogenously restoring functional β-cell mass in diabetes, and we focused on the mechanisms of the ABL-induced β-cell proliferation, hoping its therapeutic effects will be beneficial to the augmentation of β-cell mass and the enhancement of insulin secretion in patients with diabetes.
Materials and methods
Study design
Eight-week-old male C57BL/6J mice were randomly divided into three groups:
Sham group: Mice received sham operation;
PPx control group: Mice were administered saline after PPx operation; and
ABL group: Mice were administered 10 mg/kg body weight ABL after PPx operation.
ABL was purchased from the Sigma-Aldrich company (St Louis, MO, USA). For the ABL group, all the mice were treated with ABL dissolved in 0.9% NaCl+50 mmol/L Na2CO3 for consecutive 14 d. Then all mice were sacrificed, and the remnant pancreases were harvested for the study of drug efficacy. All animals were handled according to guidelines of the Sichuan University Medical Center Institutional Animal Care and Use Committee.
Animal surgery
Animals were fasted for 12 h before operation. Five hours after the operation, animals were allowed free access to standard diet and water. Mice were anesthetized with 10% chloral hydrate (Sigma, Bellefont, PA, USA) and the abdomen was opened through an upper midline incision. The spleen and the entire splenic portion of the pancreas were surgically removed, but the mesenteric pancreas between the portal vein and duodenum was left intact. This resulted in a 70% pancreatectomy, which was confirmed by weighing the removed and remnant portions. The sham operation was performed by removing the spleen while leaving the pancreas intact. The incision was closed using 5-0 silk sutures. The proliferating cells were labeled with bromodeoxyuridine (BrdU) (0.8 mg/mL; Sigma) in drinking water, and this labeling continued for 14 d after operation.
Biochemical measurements
The fasting blood glucose concentrations were measured by a Surestep Blood Glucose meter (Lifescan, Milpitas, CA, USA), and any blood glucose concentrations higher than 500 mg/dL were not included. Intravenous glucose tolerance tests (IVGTTs) were performed by tail vein injection of
Immunoblotting
The pancreatic duct was injected with 1 mL collagenase P (Roche, Indianapolis, IN, USA). Islets were isolated as described, 17 handpicked, washed once in phosphate-buffered saline and frozen at −80°C. For Western blots, islets were lysed and the protein lysates were centrifuged at 12,000 rpm for 30 min. Supernatants were collected and loaded in a sodium dodecylsulfate polyacrylamide gel electrophoresis (PAGE) gel (Bio-Rad, Hercules, CA, USA) for electrophoresis. After electrophoresis, proteins were transferred onto polyvinylidene fluoride membrane (Bio-Rad) in transfer buffer. Membranes were blocked with 5% non-fat dry milk in Tris-buffered saline for one hour. Blots were incubated overnight at 4°C with primary antibodies at the dilutions recommended by the manufacturer, followed by incubating with horseradish peroxidase-conjugated secondary antibody (Zymed, San Francisco, CA, USA), and then visualized by enhanced chemiluminescence reagents (Amersham, Piscataway, MA, USA). Antibodies for cyclin D1, cyclin D2, cyclin D3, Cdk4 and actin were obtained from Abcam (Cambridge, MA, USA). Polyclonal antibody for phospho-RB (S780) was obtained from Cell Signaling (Beverly, MA, USA), and polyclonal antibody for RB was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Quantitative reverse transcription polymerase chain reaction analysis
RNA was extracted using the RNeasy micro kit (Qiagen, Valencia, CA, USA) and reverse transcribed (1 μg total RNA, 1 μL random primer [50 μmol/L; Applied Biosystems, Foster City, CA, USA], 1 × reverse transcriptase buffer and 10 units reverse transcriptase [Promega, Madison WI, USA] in a total volume of 20 μL). The RNA and primer were heated to 72°C and slowly cooled before reverse transcription at 42°C for one hour. The room temperature reaction was then diluted to 100 μL with RNase-free water. For realtime polymerase chain reaction (PCR) analysis, 2.5% of the total room temperature reaction was used as input for PCR using SYBR Green Master Mix (Applied Biosystems) on an ABI 7900 Real-Time PCR System. Primers for cyclin D1, cyclin D2, cyclin D3, Cdk4, insulin, glucagon, GLUT-2 (glucose transporter 2), glucokinase, PDX-1 (pancreatic and duodenal homeobox 1) and Ngn3 (neurogenin 3) were previously described. 18–20 For analysis of Ngn3 mRNA abundance, RNA was extracted from the remnant pancreas, and for analysis of other genes, RNA was extracted from islets.
Cdk4 kinase activity
Cdk4 kinase assays were carried out based on protocols described previously. 21,22 Pooled islets from remnant pancreas of 30 mice were lysed in lysis buffer, and protein concentration was determined using a BCA Protein Assay kit (Beyotime, Suzhou, China). For each sample, 300 μg of total protein was immunoprecipitated using 1 μg anti-Cdk4 (Abcam) and 50 μL protein G Sepharose beads (Sigma). The final kinase reaction was carried out in 50 mmol/L HEPES, pH 7.5, 10 mmol/L MgCl2, 1 mmol/L dithiothreitol, 2.5 mmol/L EGTA, 10 mmol/L glycerophosphate, 0.1 mmol/L Na2VO4, 1 mmol/L NaF, 5 μmol/L ATP, 6 mCi per reaction of [γ-32P]ATP (Amersham) and GST-Rb 769-921 (Santa Cruz Biotechnology). Samples were incubated at 30°C for 30 min and separated by PAGE. The amount of 32P-labeled GST-Rb was evaluated by autoradiography and quantified using PhosphorImager and ImageQuant (Molecular Dynamics, Sunnyvale, CA, USA) analysis.
Histology staining
Tissues were fixed in 10% formalin for 24 h at room temperature, dehydrated, embedded in paraffin and sectioned. For immunohistochemical staining, rat anti-BrdU antibody (Abcam), guinea-pig anti-insulin antibody (Dako, Glostrup, Denmark) and rabbit anti-cyclin D2 antibody (Abcam) were used. Primary antibodies were detected with corresponding secondary antibodies (Jackson Immunoresearch, West Grove, PA, USA). For transferase-mediated dUTP nick-end labeling (TUNEL) staining, the Promega fluorescence Dead End Kit was used following the manufacturer's instructions.
Islets were photographed at ×400 by a Nikon 80i microscope (Tokyo, Japan). All photograph and statistical works were performed by two blinded technicians. The number of β-cells, BrdU(+) β-cells and TUNEL(+) β-cells was manually counted, and double-checked with Image Pro Plus 6.3 software (Media Cybernetics, Silver Spring, MD, USA). For statistics, at least 10 islets with at least 1000 β-cells were counted per mouse. In addition, at least 10 consecutive sections from six mice per group were stained. The first section and the 10th section were selected to ensure a 50-μm distance. Islet diameters were calculated with an intraocular calibrated grid. Islet size was also measured at ×400 image of insulin-stained islets converted to grayscale.
Analysis of relative β-cell volume was performed by point counting morphometry, using a 56-point grid. An average of 10,000 points/mouse was counted. β-Cell mass was calculated by multiplying the relative β-cell volume by the total weight of the remnant pancreas.
Statistical analysis
All data were expressed as mean values ± SEM from at least three independent experiments. Statistical significance was determined by one-way analysis of variance with Bonferroni correction and Student's t-test.
Results
Increased insulin release induced by ABL to restore euglycemia
Based on the results of the pilot dose-finder study, we used a slightly higher dose of ABL (10 mg/kg body weight) delivered continuously for 14 d, a window during which regeneration is known to be most active in rodents,
23–25
for the pancreatic regeneration studies. Daily blood samples from the PPx control mice, the sham-operated mice and the ABL-treated mice were analyzed for the fasting glucose concentrations over a period of 14 d postsurgery. Measurement of the fasting blood glucose concentrations revealed that the PPx control mice displayed fasting blood glucose concentrations consistently >300 mg/dL, indicating that these mice were unable to maintain glucose homeostasis (Figure 1a). In contrast, the ABL-treated mice did not show fasting blood glucose concentrations that were significantly >300 mg/dL, since seven days after operation. Glucose tolerance tests showed that blood glucose concentrations of the PPx control mice, after seven days of recovery, remained >350 mg/dL by the end of the 120-min testing period. In contrast, the ABL-treated mice, after seven days of therapy, reached ≤300 mg/dL by the end of the 120-min testing period (Figure 1b). Glucose tolerance tests performed on day 14 revealed significant differences between the PPx control mice and the ABL-treated mice by the end of the 120-min testing period (Figure 1c, P < 0.05).
Agaricus bisporus lectin (ABL) induces metabolic changes in mice after 70% partial pancreatectomy (PPx). (a) Comparisons of fasting blood glucose concentrations. (b) Intravenous glucose tolerance test (IVGTT) on day 7. (c) IVGTT on day 14. n = 8 in each group, *P < 0.05 and **P < 0.01 versus all other groups at the same time point by one-way analysis of variance, #
P < 0.05 versus PPx control group at the same time point by Student's t-test. Sham group (black triangles), PPx control (black squares) and PPx+10 mg/kg ABL (black circles). Data are means ± SEM
Analysis of proliferation
To investigate whether ABL could induce β-cell proliferation, proliferated cells were detected by continuous BrdU incorporation. Pancreas sections stained for BrdU and insulin showed a marked increase in the number of BrdU-positive β-cells in the ABL-treated mice (Figure 2a). Quantification revealed a robust and significant increase in β-cell proliferation in mice treated with ABL (Figure 2b). Statistically, a 2.7-fold increase in BrdU-positive β-cells was observed in the mice treated with ABL compared with the control group (14.4 ± 4.8% for the ABL-treated mice versus 5.3 ± 3.6% for the PPx control mice, P < 0.05). To determine whether β-cell proliferation in response to ABL varied with islet size, islets were stratified by the number of β-cells in islet cross-section (Figure 2c). Interestingly, in the ABL-treated mice, small islets (<20 β-cells) had higher rates of proliferation than larger islets, albeit ABL treatment increased β-cell proliferation in all islet sizes (Figure 2c, P < 0.05). Meanwhile, TUNEL staining was performed as a measure of apoptosis. Statistically, no significant difference in apoptosis percentages was observed in the ABL-treated group and the PPx control group (data not shown). Because after PPx, the proliferation/neogenesis from both β-cells and duct cells contributes to the β-cell hyperplasia, the BrdU-labeled duct cells were observed in different groups (Figure 2d). By manually counting and calculating the ratio of the number of BrdU-positive duct cells to the number of total duct cells, we found out that there were more proliferated duct cells in pancreas of the ABL-treated mice than those of controls (Figure 2e). Therefore, ABL contributes to the proliferation of both β-cells and duct cells.
Agaricus bisporus lectin (ABL) induces islet regeneration. (a) Representative pictures of immunohistochemistry staining of insulin-positive cells (red) and bromodeoxyuridine (BrdU)-labeled cells (brown) of different groups. Arrows point to the BrdU-labeled cells. Scale bar represents 100 μm. (b) BrdU-positive percentages of insulin-positive β-cells of different groups on day 14. At least 10 islets with at least 1000 β-cells were counted per mouse (n = 6). *P < 0.05 versus sham group by Student's t-test; **P < 0.01 versus all other groups by one-way analysis of variance (ANOVA). (c) Assessment of the distribution of proliferated β-cells in different islet size on day 14 (*P < 0.05 by Student's t-test). (d) Representative pictures of immunohistochemistry staining of insulin-positive cells (red) and BrdU-labeled cells (brown). Arrows point to the BrdU-labeled duct cells. Scale bar represents 100 μm. (e) BrdU-positive percentage of duct cells. *P < 0.05 versus sham group by Student's t-test; **P < 0.01 versus all other groups by one-way ANOVA. Data are means ± SEM. PPx, partial pancreatectomy. (A color version of this figure is available in the online journal)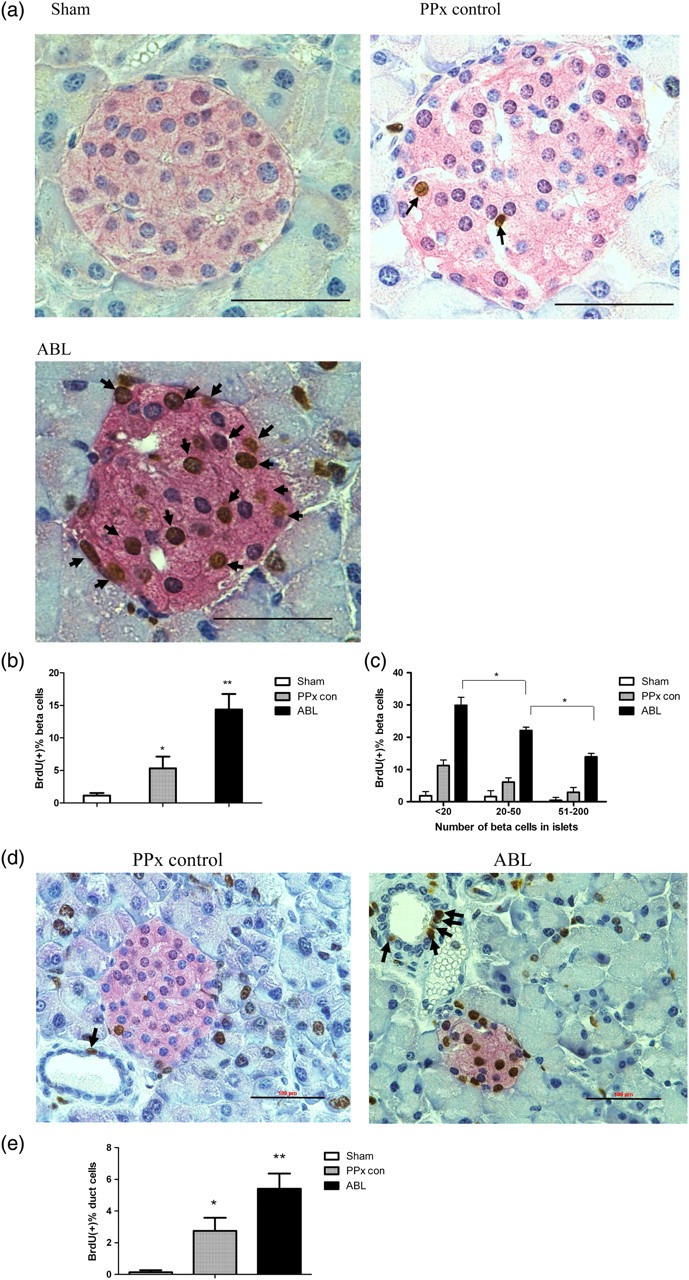
Analysis of β-cell mass
Because the reduced blood glucose concentrations were caused by the increased serum insulin concentration, we next assessed the serum insulin concentrations. The ABL-treated mice had elevated serum insulin concentrations compared with those of the PPx control mice (Figure 3a, P < 0.05). To investigate whether ABL therapy would induce islet hyperplasia, the relative β-cell volume in the remnant pancreas and β-cell mass was subsequently assessed. The ABL-treated group got a significant increase in relative β-cell volume (Figure 3b, P < 0.05) compared with that of the PPx control group and the sham-operated group. Thus, absolute β-cell mass in the remnant pancreas was significantly elevated in the ABL-treated mice (Figure 3c, P < 0.05).
Agaricus bisporus lectin (ABL) induces β-cell hyperplasia. (a) Comparisons of fasting serum insulin concentrations. (b) Relative β-cell volume by point counting. (c) β-Cell mass calculated by relative β-cell volume by the total weight of remnant pancreas. Two or three slides (200 μm apart) from the broadest pancreatic sections were analyzed for β-cell mass measurement (n = 6 for each group). All graphs show means ± SEM from three independent experiments (#
P < 0.05 versus PPx control group by Student's t-test, *P < 0.05 and **P < 0.01 compared with all other groups by analysis of variance.). Data are means ± SEM. PPx, partial pancreatectomy
Analysis of the cyclin D-Cdk4 complex
To further investigate the mechanisms by which ABL-induced β-cell proliferation, key components in cell cycle regulation such as cyclin D1, cyclin D2, cyclin D3 and cyclin-dependent kinase 4 (Cdk4) were studied.
14
Cyclin D–Cdk4 complex plays an essential role in β-cell cycle progression.
26
Although cyclin D3 mRNA abundance and protein concentrations were unchanged after ABL treatment (Figure 4c), a marked increase in Cdk4 mRNA and protein concentrations was observed in the ABL-treated mice compared with those of the PPx control mice by realtime PCR and immunoblotting (Figure 4d). Meanwhile, cyclin D1 (Figure 4a) and cyclin D2 (Figure 4b) mRNA and protein concentrations were also significantly increased after ABL treatment. Furthermore, elevated nuclei expression of cyclin D2 after ABL treatment was observed by co-immunostaining of insulin (green), BrdU (blue) and cyclin D2 (red) (Figure 4e). Although the ratios of cyclin D2 and BrdU double-positive β-cells to BrdU-positive β-cells were the same between the PPx group and the ABL group, more β-cells were found to express cyclin D2 in ABL-treated mice, suggesting that the upregulation of cyclin D2 allows β-cells to rapidly re-enter the proliferative pool.
Cyclin D1, D2 and D3 concentrations and Cdk4 levels were assessed in islets from Agaricus bisporus lectin (ABL)-treated and -untreated mice. Immunoblotting and realtime polymerase chain reaction analyses of cyclin D1 (a), cyclin D2 (b), cyclin D3 (c) and Cdk4 (d) (*P < 0.05 and **P < 0.01 versus all other groups by one-way analysis of variance). (e) Representative pictures of immunohistochemistry staining of cyclin D2 (red), BrdU (blue) and insulin (green). Scale bar represents 100 μm. All graphs show means ± SEM from at least three independent experiments. PPx, partial pancreatectomy. (A color version of this figure is available in the online journal)
Analysis of Cdk4 activity
To determine whether the effect of ABL-induced islet β-cell proliferation was associated with Cdk4 activity, in vitro kinase activity assays were conducted using recombinant GST-Rb (amino acids 769–921) as substrate as previously described.
21,22
Rb plays a fundamental role in cell cycle progression through its association with the E2F family of transcription factors, and it is an important substrate of Cdk4. Recently published data revealed that Rb associates with and stabilizes PDX-1, which is essential for embryonic pancreas development and adult β-cell function.
27
In this study, the incorporation of radioactive phosphate to this substrate is proportional to Cdk4 activity in the immunoprecipitate. Phosphorylation of recombinant GST-Rb was significantly increased in islet lysates from the ABL-treated mice compared with that of the PPx control mice, suggesting that Cdk4 was activated after ABL treatment (Figure 5a). Because phosphorylation of Rb protein is mediated by the cyclin D1/Cdk4 complex,
28
immunoblotting of pRb (Ser780) was performed, and an increase in Rb phosphorylation was observed in islets isolated from the ABL-treated mice, which indicated that the Cdk4 activity was increased (Figure 5b). Taken together, we may conclude that ABL initiates β-cell proliferation through regulation of cell cycle protein.
Cdk4 activity was assessed in islets from Agaricus bisporus lectin (ABL)-treated and -untreated mice. (a) In vitro Cdk4 kinase activity in islets from ABL-treated and -untreated control mice. Islets from remnant pancreas of 30 mice were pooled by recombinant GST-Rb and lysated. (b) Immunoblotting for hyperphosphorylated Rb at Ser780 (upper panel), total Rb (middel panel) and β-actin (bottom panel) in islet lysates. The data represent one out of three independent experiments that gave similar results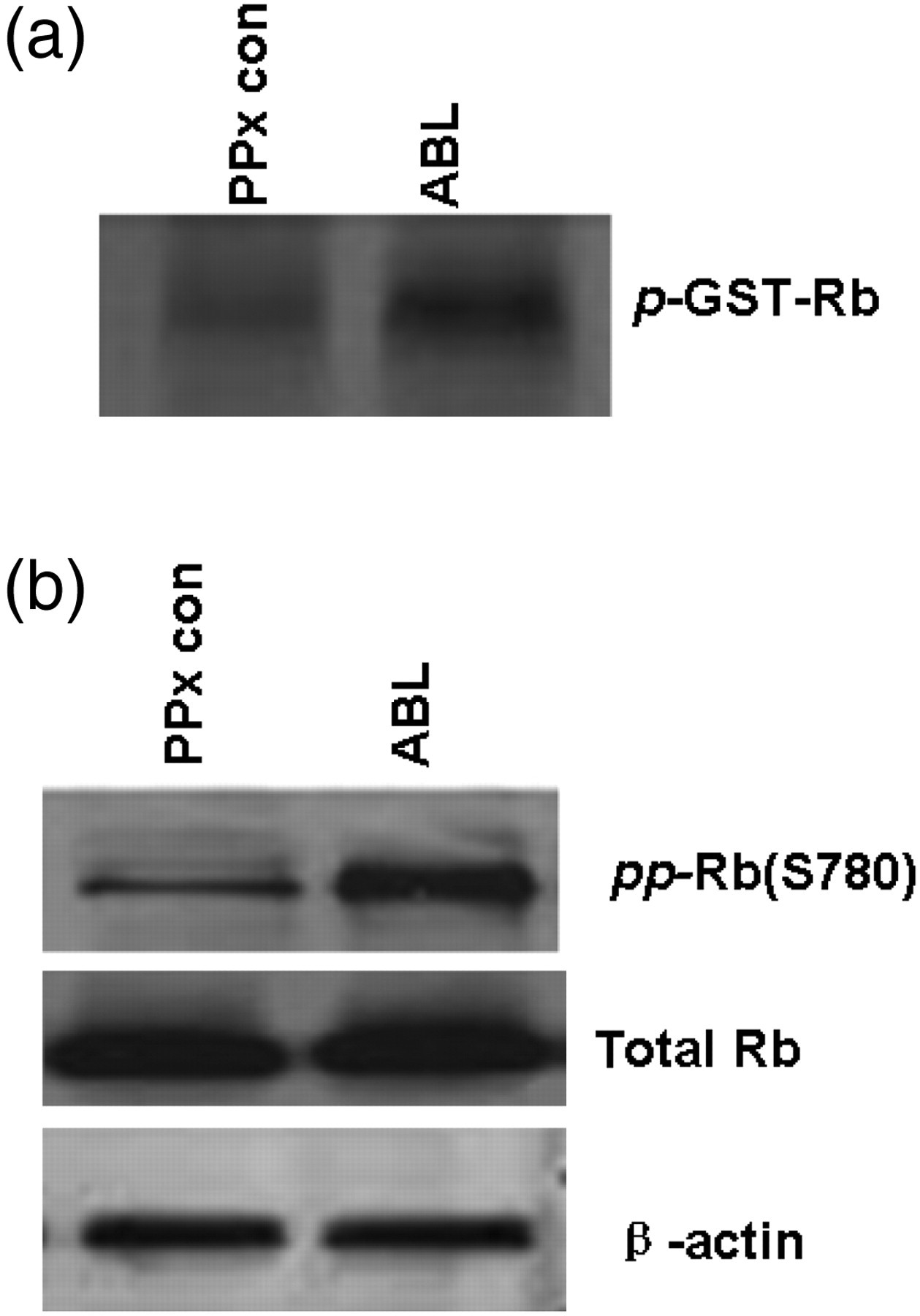
Analysis of the abundance of β-cell-specific genes
To further investigate whether the reduction of fasting blood glucose concentrations of the ABL-treated mice could be attributed to the increased expression of β-cell-specific genes (insulin, GLUT-2, glucokinase) and glucagon mRNA, realtime PCR was performed and a significant increase in insulin, GLUT-2 and glucokinase (GCK) mRNA levels was observed in the islets of the ABL-treated mice (Figure 6, P < 0.05). Meanwhile, no significant difference was observed in the expression of glucagon gene between the ABL-treated mice and the PPx control mice. Recent studies on the Cdk4 pathway have demonstrated that Cdk4 promotes β-cell replication by activating PDX-1 and Ngn3 genes,
27,29
and Ngn3 is involved in β-cell neogenesis, which also contributes to β-cell hyperplasia.
5
Immunohistochemistry staining of pancreas sections revealed that ABL treatment could induce the proliferation of both β-cells and duct cells. Thus the abundance of these two genes was studied as well, and these two genes were found out to be upregulated in ABL-treated mice (Figure 6, P < 0.05).
The expression of β-cell specific gene levels was analyzed. Insulin, GLUT-2, glucokinase, PDX-1, glucagon and Ngn3 mRNA levels were assessed by realtime PCR. All data are means ± SEM. *P < 0.05 and **P < 0.01 versus all other groups by one-way analysis of variance. The data represent one out of three independent experiments that gave similar results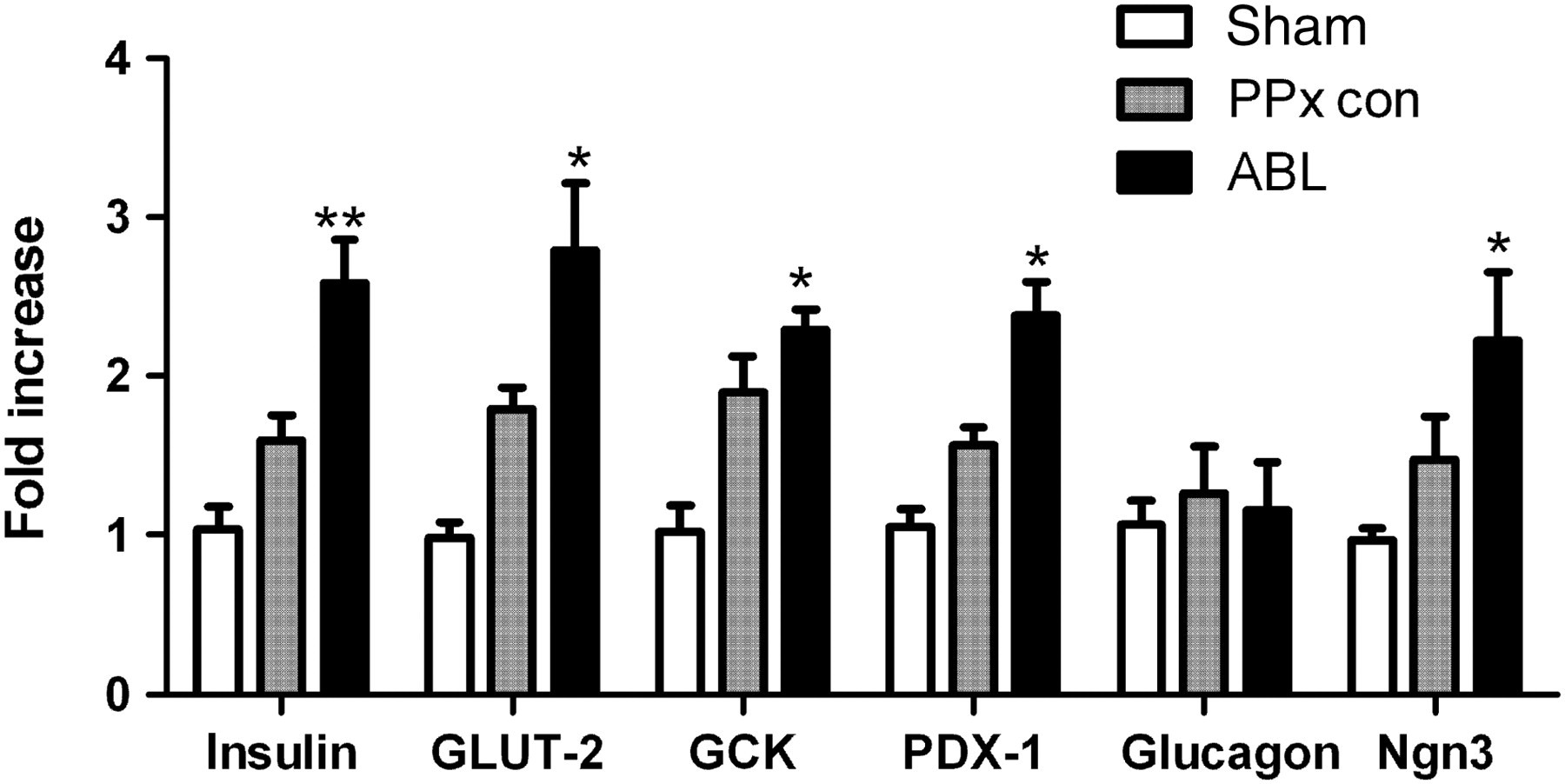
Discussion
Our main interest in studying mice islet regeneration is to pursue a therapeutic way in diabetes. Previous studies have demonstrated that during pregnancy or high-fat feeding, healthy β-cells maintain euglycemia, by increasing insulin secretion through an elevated β-cell mass. Herein, our studies reported for the first time that the regeneration ability of mice pancreas after PPx was enhanced after the treatment of ABL. In this study, we observed that accompanied with increased serum insulin, β-cell mass was adaptively elevated. Although previous reports have been focused on the insulin-stimulation effects of ABL, 7,8 studies on its mechanism are scarce. Ewart et al. 9 proposed that the interaction between ABL and its receptors contributes to the insulin-stimulating effects of ABL, and that this interaction leads to conformational changes in the structure of the membranes of the islet α- and β-cells that facilitate exocytosis. In the present study, we explored the insulin-stimulation effects of ABL from β-cell proliferation perspective, and found β-cell hyperplasia in the ABL-treated mice. Observations on the BrdU-positive percentage in islets demonstrated that the number of proliferated β-cells increased with time, which means that not all β-cells proliferate at the same time. A previous study indicated that the newly formed β-cells do not have glucose-responsive insulin secretion characteristics. 30 Because not all β-cells proliferate at the same time, and not all proliferated β-cells are functionally effective at the same time, blood glucose concentrations of ABL-treated mice could hardly return to control values, but nearly reach to control values on day 14. However, measurement of blood glucose concentrations and glucose tolerance tests indicated that mice could maintain glycemia, to some degree, after receiving ABL treatment. So the discrepancy between the reduced glucose concentrations and expanded β-cell mass is due to the immaturity of some newly formed β-cells on the sacrificed time. Taken together, ABL treatment facilitates β-cell proliferation and most proliferated β-cells are functionally effective. These studies demonstrated that ABL could play a significant role in the adaptive expansion and regeneration of β-cell mass after PPx.
Defining the mechanisms by which ABL regulates the proliferation of pancreatic β-cells is an essential prerequisite for the study of ABL pharmacological function. Studies focused on genomic analyses of type 2 diabetic patients have reported polymorphisms in genes that are close to those coding for key cell-cycle regulators, 31,32 which suggested that expansion of β-cell mass was linked to proteins in cell-cycle progression. Moreover, recent studies have demonstrated that human β-cell replication ability was enhanced under the circumstances of cyclin D3 and Cdk6 overexpression, 33 which shed light on the study of β-cell regeneration. Many reports have proved that cyclin D2 was crucial for controlling postnatal β-cell expansion. 12 In the present study, we observed significantly increased cyclin D1 mRNA and protein expression in mice treated with ABL. Additionally, cyclin D2 was also upregulated in islets of ABL-treated mice. Because cyclin D2 is more highly expressed during development and early postnatal life than during adulthood, ABL could be the intriguing possible β-cell growth stimuli which shift differentiated adult β-cells toward a more developmental phenotype. Interestingly, examination of β-cell differentiation markers in the proliferating islets showed that ABL could not possibly dedifferentiate. This could be explained by the activation of the insulin signaling pathway induced by ABL. Furthermore, because cyclin D2 is essential for the physiological remodeling of β-cell mass in the postnatal period, the ABL-treated mice had significantly expanded β-cell mass compared with that of the controls, suggesting that ABL-induced β-cell mass expansion via upregulation of cyclin D2. By co-immunostaining of BrdU, insulin and cyclin D2, we observed that both the controls and the ABL-treated mice had a similar rate of cyclin D2+BrdU+ cells/BrdU+ cells, indicating that ABL controls the upregulation of cyclin D2, which further allows β-cells to re-enter the proliferative pool. However, further evaluation of the ABL-treated mice lacking cyclin D2 will determine whether this cell cycle regulator is required for ABL-induced β-cell replication in adult mice.
Importantly, despite the upregulation of cyclin D1 and cyclin D2, Cdk4 was also upregulated in mRNA and protein concentrations. Several reports suggested that Cdk4 was necessary to maintain postnatal β-cell proliferation. 14,34 Recently published data revealed that Cdk4 plays an important role in promoting β-cell development by directing E2f1-mediated activation of Ngn3 and increasing the pool of endocrine precursors. 29 Furthermore, Lee et al. 15 performed PPx in Cdk4 mutant mice, and their study demonstrated that Cdk4 controls β-cell mass by recruiting quiescent cells to enter the cell cycle. Moreover, because Cdk4 partners with the D-type cyclins and hyperphosphorylate Rb, leading to the inactivation of hypophosphorylate-Rb and thereby cell cycle progression, studies on Rb also indicated that Rb regulates pancreas development by stabilizing PDX-1. 27 Therefore, in the present study, we explored Cdk4 in vitro activity after ABL treatment, and we also observed increased Rb hyperphosphorylation at Ser 780 in islet extracts of the ABL-treated mice. These observations suggested that ABL directly or indirectly induced expression of cyclin D-cdk4 complex, and enhance cdk4 activity.
Moreover, upstream signaling pathways such as PDX-1 are crucial in the regulation of β-cell regeneration, 35,36 and may be correlated with increased expression and activity of cyclin D–Cdk4 complex. Our study on PDX-1 by realtime PCR indicated that ABL could induce the increased expression of this gene. Besides, Xu et al. 5 have reported that in the partial duct ligation model, activation of Ngn3 gene contributes to the differentiation of the adult progenitors, and the differentiated β-cells are glucose responsive. Furthermore, studies on Cdk4 by Kim et al. 29 also showed the proliferation of Ngn3+ endocrine precursors by the expression of activated Cdk4. In our study, we observed increased proliferation of duct cells after ABL treatment. Therefore, we further analyzed Ngn3 mRNA abundance and found upregulation of Ngn3 genes in the ABL-treated mice. Despite the report that cell cycle activators lead to dedifferentiation, 37 the increased expression of β-cell-specific markers such as insulin, GLUT-2 and glucokinase was observed in ABL-treated mice, suggesting that ABL treatment could not induce dedifferentiation of islet β-cells. Taken together, consistent with a role in modulating β-cell proliferation and stimulating insulin secretion, ABL could also induce islet hyperplasia and euglycemia in diabetic mice within 14 days post-PPx.
Although it is not clear to what extent β-cell replication will be in adult humans if ABL was administered, this study supports the notion that ABL-induced β-cell replication may be a key for reversal of type 1 and 2 diabetes. Moreover, at present, we are working on whether ABL treatment has a protective effect against streptozotocin-induced diabetic mice. However, observations of its regeneration ability raised issues with respect to its antiproliferative effect on epithelial cells. 38,39 This could be explained by different cell surface receptors for ABL. Because ABL-induced islet regeneration function within two weeks was proved to be prominent, short-time application of ABL in diabetic patients is still promising. In summary, considering the overall evidence of ABL treatment on β-cell proliferation, new therapeutic application of ABL for diabetic patients may be envisaged in the future.
Footnotes
ACKNOWLEDGEMENTS
This paper is supported by grants from the National Natural Science Foundation of China (General Programs: No. 81173093, No. J1103518 and No. 30970643).