
Abstract
Studies in vivo have shown that dietary copper (Cu) supplementation reverses pressure overload-induced cardiac hypertrophy in a mouse model, which is vascular endothelial growth factor (VEGF)-dependent and correlates with enhanced angiogenesis. Because Cu stimulation of endothelial cell growth and differentiation would play a critical role in angiogenesis, the present study was undertaken to examine the effect of Cu on growth of human umbilical vein endothelial cells (HUVECs) in cultures. The HUVECs were treated with CuSO4 at a final concentration of 5 μmol/L Cu element in cultures or with a Cu chelator, tetraethylenepentamine (TEPA), at a final concentration of 25 μmol/L in cultures. Cell growth and Cu effect on cell cycle were determined. In addition, the effect of Cu on VEGF and endothelial nitric oxide synthase (eNOS) mRNA levels was determined, and anti-VEGF antibody and siRNA targeting eNOS were applied to determine the role of VEGF or eNOS in the Cu effect on cell growth. Cu significantly stimulated and TEPA significantly inhibited cell growth, and the TEPA effect was blocked by excess Cu. Cu increased the number of cells in the S phase and correspondingly decreased the number in the G1 phase. Interestingly, Cu did not increase the level of VEGF mRNA, but significantly increased eNOS mRNA. Furthermore, neutralizing VEGF by anti-VEGF antibody did not suppress Cu stimulation of cell growth. However, siRNA targeting eNOS completely blocked Cu reversal of TEPA inhibition of cell growth. The data demonstrate that Cu stimulation of HUVEC cell growth is VEGF-independent, but eNOS-dependent.
Introduction
Previous studies in vivo have shown that dietary copper (Cu) supplementation reverses pressure overload-induced cardiac hypertrophy and prevents heart failure in a mouse model. 1 This effect is vascular endothelial growth factor (VEGF)-dependent and is associated with myocardial angiogenesis. 1 At cellular levels, there would be two different mechanisms that govern the Cu-induced regression process. First, the direct effect of Cu on hypertrophic cardiomyocytes to reduce their size would make a critical contribution to the regression of cardiac hypertrophy. Second, the effect of Cu on endothelial cells to promote angiogenesis or vasculogenesis would play another crucial role in the regression.
We have observed that Cu can indeed reduce the size of hypertrophic cardiac myocytes both in vivo 2 and in vitro. 3,4 Interestingly, we have observed that although VEGF is required for Cu-induced regression of cardiomyocyte hypertrophy, Cu itself at the effective and physiologically relevant level does not increase VEGF production. 4 However, Cu differentially changes the distribution of VEGF receptor-1 (VEGFR-1) versus VEGFR-2, increasing the ratio of VEGFR-1 to VEGFR-2 in the hypertrophic cardiomyocytes in cultures. 4 This increase plays a determinant role in Cu-induced regression of cardiomyocyte hypertrophy. Gene silencing of VEGFR-1 prevents the increase by Cu of the VEGFR-1/VEGFR-2 ratio and leads to suppression of both Cu- and VEGF-induced regression of cardiomyocyte hypertrophy. Conversely, gene silencing of VEGFR-2 increases the ratio of VEGFR-1/VEGFR-2 independent of Cu treatment and results in the regression of cardiomyocyte hypertrophy. 4 It appears that VEGFR-1 is linked to the c-GMP-dependent protein kinase (PKG)-1 pathway because inhibition of PKG-1 suppresses the VEGFR-1-mediated regression of cardiomyocyte hypertrophy. 4
The in vitro studies 3,4 have provided significant insights into Cu regression of cardiac hypertrophy. 5 In particular, the direct action of Cu on cardiomyocytes made a clear distinction between the effects of Cu on cardiomyocytes and on endothelial cells, which are essential for angiogenesis. The reversal by Cu of cardiomyocyte hypertrophy in vitro is VEGF-dependent, but angiogenesis-independent. However, it has been unequivocally shown that the regression of cardiac hypertrophy by Cu supplementation in vivo is angiogenesis-dependent. 1 Therefore, the role of Cu in endothelial cell growth and differentiation is equally important in the understanding of Cu regression of cardiac hypertrophy.
Cu promotion of angiogenesis has been known for a long time. 6 Cu stimulates endothelial cell proliferation in cultures, 7–12 and this stimulation is accompanied by enhanced production of VEGF. 13,14 However, it is worthy to note that most of these studies, if not all, have used Cu concentrations much more than physiologically relevant levels, ranging from 50 to 500 μmol/L. In our recent studies, we have used a Cu concentration of 5 μmol/L in cultures, 3,4 derived from what would be achieved in the blood of mice under the condition of dietary Cu supplementation at 20 mg Cu/kg diet. 1 It is thus critical to determine the effect of Cu on endothelial cell growth at its physiologically relevant levels, so that the mechanistic studies in vitro can be extrapolated to in vivo experimental conditions and provide valuable insights into the observation from animal studies.
There are several factors involved in the regulation of endothelial cell growth and function, and the most studied are VEGF 15–17 and endothelial nitric oxide synthase (eNOS). 18–20 VEGF has been shown to play a pivotal role in the initiation and formation of blood vessels, which involve endothelial cell growth and differentiation. 15–17 We have demonstrated that Cu is required for VEGF expression through its regulation of hypoxia-inducible factor (HIF) activity, 21 a transcription factor that is required for VEGF expression. This Cu requirement for VEGF expression does not explain the effect of high levels of Cu on VEGF over-production. High levels of Cu stabilize HIF-1α, leading to HIF-1α accumulation and enhanced HIF transcriptional activity. 22,23 As mentioned above, under the condition of cells exposed to physiologically relevant levels of Cu, VEGF levels were not increased in cardiomyocytes in cultures. 4 It is important to know if this is true in the endothelial cells.
The effect of eNOS on cell growth is less studied. The eNOS is responsible for the production of NO in blood vessels and the major effect of NO on cell growth appears to regulate cell cycle progression. It has been demonstrated that high NO concentrations promote the G1 to S phase transition, while low NO promotes the G2 to M transition, which is mediated by a cGMP-dependent pathway. 24 It is interesting to know if Cu stimulation of endothelial cell growth is related to eNOS.
The present study was undertaken to determine the effect of Cu at physiologically relevant levels on growth of human umbilical vein endothelial cells (HUVECs) in cultures. The effect of Cu on VEGF or eNOS expression and conversely the effect of suppression of VEGF or eNOS on Cu stimulation of cell growth were determined. The results obtained demonstrated that Cu at physiologically relevant levels stimulates growth of HUVECs in cultures and this stimulation is eNOS-dependent, but VEGF-independent.
Methods
Cell culture and treatments
A HUVEC line (American Type Culture Collection, Manassas, VA, USA) was cultured in MCDB 131 Medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Hyclone, Logan, UT, USA) without antibiotics. Cells were routinely maintained in a standard culture incubator with humidified air containing 5% CO2 at 37°C. For treatment with reagents, cells were seeded in 96-well plates and grown overnight. Thereafter, tetraethylenepentamine (TEPA; Sigma, St Louis, MO, USA) or CuSO4 (Kelong, Chengdu, China) was added to the cultures, followed by further incubation for a period of time indicated. Cell numbers in cultures were counted by a hemocytometer.
MTT assay
HUVECs were seeded onto 96-well plates at 5000 cells/well in 0.2 mL MCDB 131 media without serum or antibiotics. Cell growth was determined by a colorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Amresco, Solon, OH, USA) method. In brief, cultured cells were washed with phosphate-buffered saline (PBS), refreshed with fresh MCDB 131 media and then followed by addition of 20 μL MTT solution (5 mg/mL). After further incubation for 3.5 h, the MTT solution was carefully removed from each well, and 100 μL dimethyl sulfoxide (Kelong) was added to dissolve the MTT formazan crystals. The absorbance was measured at 490 nm using a spectrophotometer (Spectra Max190; Molecular Devices, Sunnyvale, CA, USA).
3H-radioactive thymidine incorporation assay
HUVECs treated under different conditions for 48 h were exposed to 1 μCi 3H-radioactive thymidine (TdR) in cultures for six hours. DNA was extracted from the cultures at the end of exposure and 3H-TdR-labeled DNA was measured by the radioactivity, determined by a liquid scintillation counter (Beckman LS 6500; Beckman Coulter, Brea, CA, USA).
Cell cycle analysis
To analyze the distribution of the HUVECs in the cell cycle under different treatment conditions, cells were trypsinized (Invitrogen), washed with PBS and fixed in 75% ethanol at 4°C for at least 18 h. The HUVECs were then incubated for 30 min in DNA-staining buffer, which contained 200 μg/mL ribonuclease A (Roche, Indianapolis, IN, USA) and 5 μg/mL propidium iodide (PI; Roche). The PI-stained cells were analyzed using a flow cytometer (BD FACSAria™ I Cell Sorter; Becton Dickinson, Franklin Lakes, NJ, USA). For each sample, 1 × 104 cells were captured and cells in different phases of the cell cycle were expressed as a percentage of the total number of cells counted with the BD FACS Diva software (Becton Dickinson).
Realtime quantitative reverse transcriptase polymerase chain reaction
Total RNA was extracted from HUVECs which were harvested after specified treatment times using RNAiso Plus (Takara, Shiga, Japan) according to the manufacturer's instruction. Complementary DNAs (cDNAs) were synthesized using a SYBR® PrimeScript™ reverse transcriptase polymerase chain reaction (RT-PCR) kit (Takara), under the following conditions: denaturation at 95°C (30 s); 50 cycles at 95°C (5 s) and 60°C (20 s) in a multicolor realtime PCR detection system (Bio-Rad, Hercules, CA, USA). The amount of cDNA corresponding to 100 ng of RNA was amplified using an SYBR® PrimeScript™ RT-PCR kit (Takara) with the primers for human β-actin, VEGF or eNOS (Takara). The primer sequences were designed through the software offered by Takara. Oligonucleotide primer sequences for each gene are presented in Table 1. Realtime PCR reactions were performed, recorded and analyzed by the iCycler (Bio-Rad), and β-actin was used as an internal reference in each reaction.
RT-PCR primer and siRNA sequences
RT-PCR, reverse transcriptase polymerase chain reaction; VEGF, vascular endothelial growth factor; eNOS, nitric oxide synthase
Neutralization of VEGF
HUVECs were cultured in 96-well plates in 0.2 mL MCDB 131 media without serum or antibiotics overnight. Then the cultures were refreshed by the same media containing anti-VEGF antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at a final concentration of 2 ng/mL for two days. Cell growth was assessed using the MTT assay as described above.
Gene silencing of eNOS
siRNA targeting human eNOS and negative mismatched control were designed and synthesized from RiboBio (Guangzhou, China). The sequences for siRNA targeting eNOS are presented in Table 1. The optimal transfection efficiency was determined from our preliminary study testing the range from 5 to 100 nmol/L, and we selected the condition that the siRNA caused an optimal silencing effect with minimal cytotoxicity. After HUVECs were transfected with 10 nmol/L annealed siRNA targeting human eNOS or negative mismatched control siRNA in serum- or antibiotics-free media, a Lipofectamine 2000 reagent (Invitrogen) was used as the transfection reagent according to the manufacturer's instruction. After 24-h transfection, cells were trypsinized and collected for further analysis as described in the Methods section.
Statistical analysis
Data were obtained from three separate experiments and expressed as means ± SEM, and analyzed by Student's t-test for the data presented in Figures 1a, c, Figures 2 and 3a and b. A parallel line analysis was applied to Figure 1b and d, and a 2 × 2 factorial design was applied for the data presented in Figure 3c. The level of significance was considered when P < 0.05.

The effect of Cu on growth of HUVECs in cultures. Cells were cultured with 5 μmol/L CuSO4 as a function of time (a) or with different concentrations of CuSO4 for 48 h (b) before subjected to the MTT assay. For the cell cycle analysis, cells were cultured in the presence or absence of 5 μmol/L CuSO4 for 48 h before collection (c). The data expressed in ‘percentage’ refer to the treatment groups relative to the control that was designated as 100%. Each group of data was obtained from three independent experiments and each experiment contains triplicate samples for each treatment. Values are means ± SEM. *Significantly different from control group (P < 0.05). HUVEC, human umbilical vein endothelial cell; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
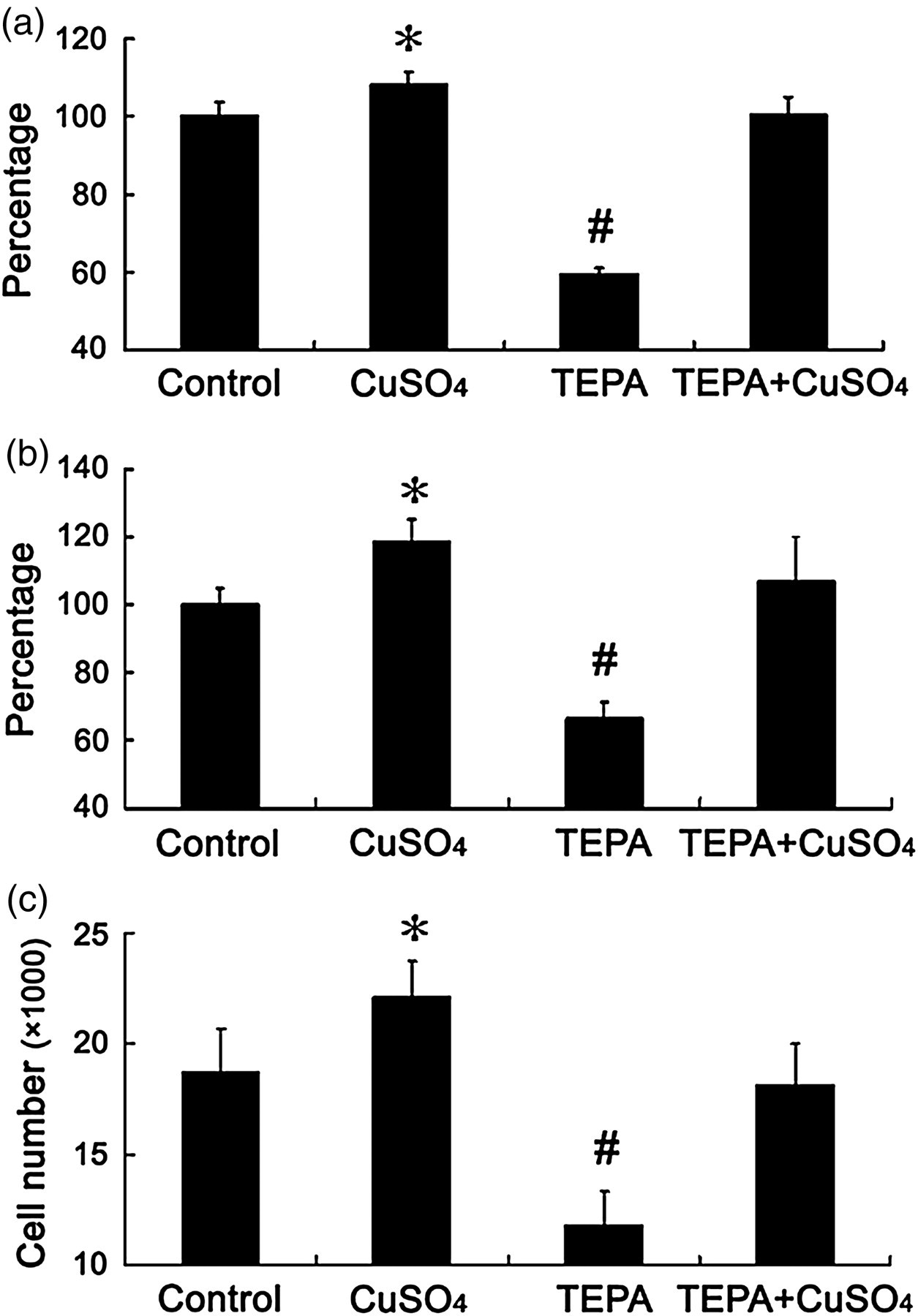
Effects of Cu chelator, TEPA, on cell growth. Each well was seeded with 5000 cells and treated with 25 μmol/L TEPA for 48 h with or without concomitant presence of 25 μmol/L CuSO4 for the MTT assay (a), 3H-TdR incorporation assay (b) and cell count (c). The data presented as ‘percentage’ are as defined in Figure 1. Each group of data was obtained from three independent experiments and each experiment contains triplicate samples for each treatment. Values are means ± SEM. *,#Significantly different from control and other treatment groups, respectively (P < 0.05). TEPA, tetraethylenepentamine; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; TdR, radioactive thymidine
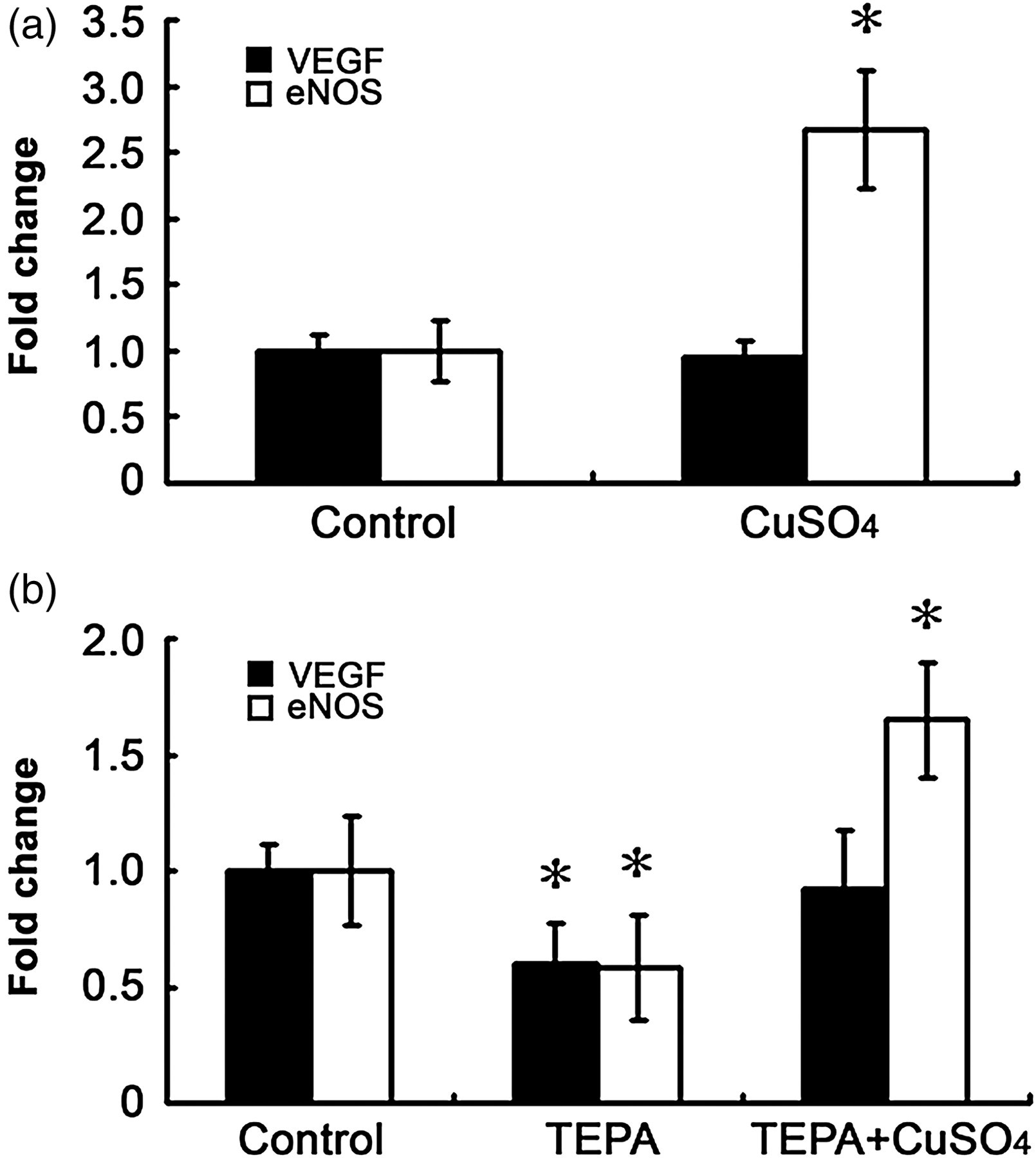
Effects of Cu on the levels of mRNA for VEGF or eNOS in cultured HUVECs. The cells were treated for 48 h with or without 5 μmol/L CuSO4 (a), or treated with 25 μmol/L TEPA for 48 h with or without a concomitant presence of 25 μmol/L CuSO4 (b) before subjected to realtime RT-PCR analysis. The data presented as ‘percentage’ are as defined in Figure 1. Each experiment was repeated three times and each experiment contains triplicate samples. Values are means ± SEM. *Significantly different from the control group (P < 0.05). TEPA, tetraethylenepentamine; HUVEC, human umbilical vein endothelial cell; eNOS, nitric oxide synthase; RT-PCR, reverse transcriptase polymerase chain reaction; VEGF, vascular endothelial growth factor
Results
Effect of Cu on growth of HUVECs in cultures
The HUVECs in cultures were first treated with CuSO4 at a final concentration of 5 μmol/L in cultures to examine the effect of Cu on cell growth as a function of time. As presented in Figure 1a, the results obtained from the MTT assay showed that Cu stimulated cell growth after the treatment for 48 h. To determine the physiological range of Cu under the experimental condition, these cells were exposed to different concentrations of CuSO4 for a total of 48 h. Cu stimulated cell growth within the concentration range below 100 μmol/L but caused a cytotoxic effect beyond 200 μmol/L in cultures (data not shown). The data presented in Figure 1b showed the effect of Cu concentrations at 0, 5, 50 and 500 μmol/L; there was no significant difference between the concentrations 5 and 50 μmol/L, but a cytotoxic effect at 500 μmol/L. The cell cycle analysis shows that Cu significantly increased the number of cells in the S phase, and decreased the number in the G1 phase (Figure 1c).
Further analysis revealed that addition of TEPA to the cultures at a final concentration of 25 μmol/L for 48 h significantly inhibited cell growth, but this inhibition can be reversed by the addition of an equal molar concentration of CuSO4 in the cultures, as determined by the MTT assay (Figure 2a). The 3H-TdR incorporation assay (Figure 2b) and cell counts (Figure 2c) confirmed these results of the effect of TEPA on Cu promotion of cell growth.
Effect of Cu on VEGF or eNOS expression
To determine if VEGF or eNOS is involved in the Cu stimulation of cell growth, we measured the levels of VEGF or eNOS mRNA in cultures treated with Cu at a final concentration of 5 μmol/L for 48 h. The results presented in Figure 3a show that Cu did not increase the level of mRNA for VEGF, agreeing with what has been observed in primary cultures of neonatal rat cardiomyocytes. 4 In addition, the measurement of VEGF levels in culture media using an enzyme-linked immunosorbent assay did not show any significant changes induced by 5 μmol/L Cu (data not shown). However, Cu significantly increased the level of mRNA for eNOS. To further probe the effect of Cu on VEGF or eNOS expression, the cells were treated with TEPA at a final concentration of 25 μmol/L for 48 h. As shown in Figure 3b, TEPA significantly depressed both VEGF and eNOS expression, demonstrating the requirement of Cu for the gene expression. Importantly, when an equal molar concentration of Cu was added to the TEPA-treated cultures, TEPA-inhibited VEGF expression was relieved to normal, but TEPA-inhibited eNOS expression was not only relieved but also significantly rebounded (Figure 3b).
Effect of VEGF or eNOS on Cu stimulation of cell growth
To determine whether or not VEGF is involved in the Cu stimulation of cell growth, the cells were neutralized by an anti-VEGF antibody at a final concentration of 2 ng/mL in cultures. As shown in Figure 4a, this neutralization did not block the stimulating effect of Cu on cell growth, which was validated by the result that the same antibody blocked the stimulating effect of exogenously added VEGF (10 ng/mL in cultures). To probe the role of eNOS in Cu stimulation of cell growth, a validated siRNA targeting eNOS was applied and it effectively suppressed eNOS expression, as shown in Figure 4b. Under the condition of eNOS depression, normal cell growth was not affected, but the Cu reversal of TEPA-inhibited cell growth was completely blocked (Figure 4c).
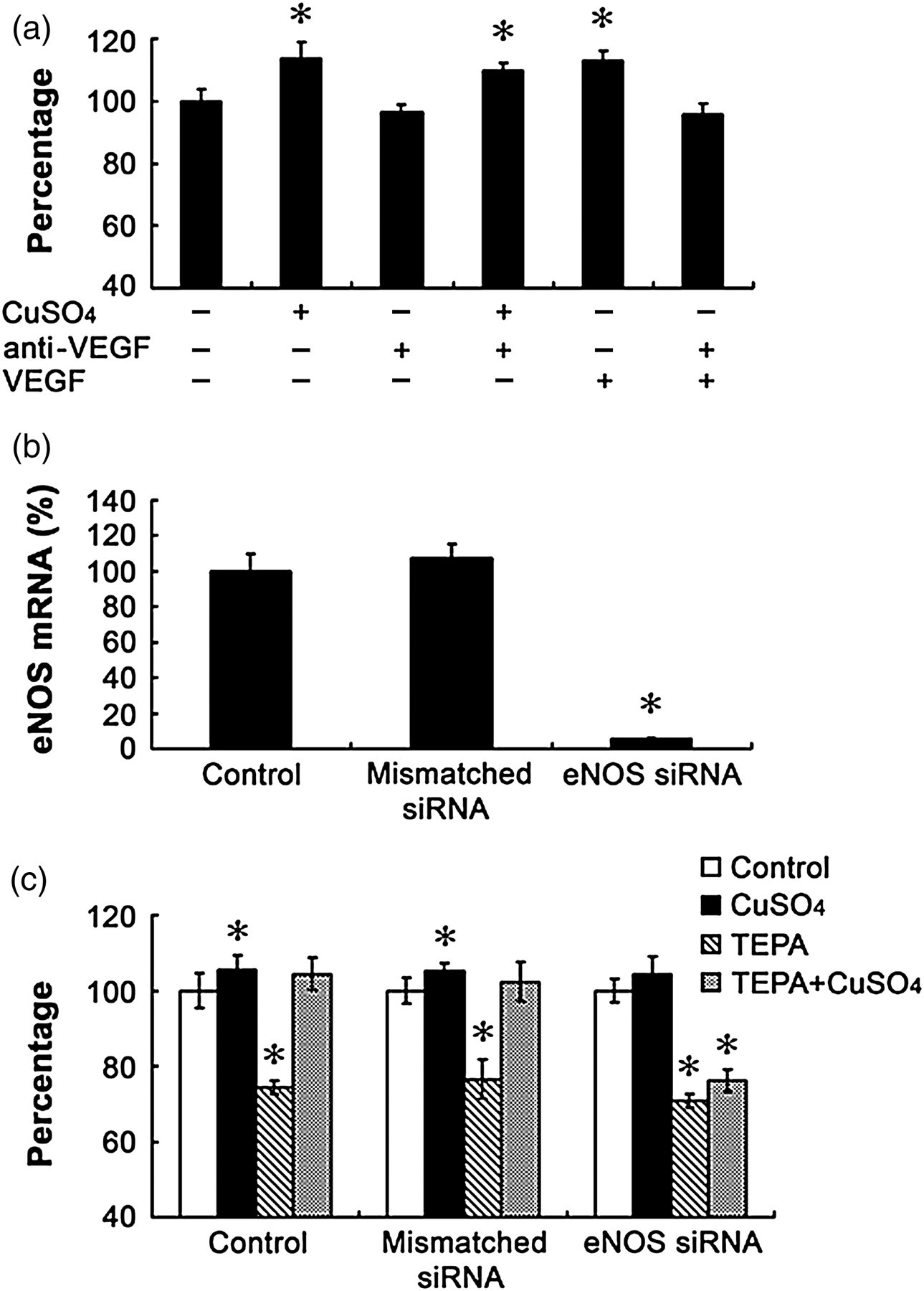
Effects of anti-VEGF antibody or siNRA targeting eNOS on growth of HUVECs in cultures. (a) Cells were treated for 48 h with anti-VEGF antibody at a final concentration of 2 ng/mL in cultures in the presence or absence of 5 μmol/L CuSO4. In a separate experiment, VEGF was added to the cultures at a final concentration of 10 μmol/L in the presence or absence of anti-VEGF antibody for 48 h as a positive control for the validation of the efficacy of the anti-VEGF antibody. (b) Effect of a validated siRNA targeting eNOS on the mRNA levels of eNOS after treatment for 48 h in comparison with the mismatched siRNA controls. (c) Effects of eNOS silencing by siRNA targeting eNOS on cell growth and its interaction with TEPA effects on cell growth. The data presented as ‘percentage’ are as defined in Figure 1. Data were obtained from three independent experiments and each experiment contains triplicate samples for each treatment. Values are means ± SEM. *Significantly different from the control group (P < 0.05). HUVEC, human umbilical vein endothelial cell; eNOS, nitric oxide synthase; VEGF, vascular endothelial growth factor
Discussion
Cu stimulation of cell growth has been known for a long time and the underlying mechanism has been linked to Cu stimulation of VEGF expression. 13,14 However, the data obtained from the present study demonstrate that Cu stimulation of HUVEC cell growth at the physiologically relevant level is not VEGF-dependent, but is eNOS-dependent. This provides a novel insight into the understanding of Cu promotion of angiogenesis and Cu-induced regression of cardiac hypertrophy. It appears that VEGF is required for the growth of endothelial cells, but eNOS is more responsible for the stimulation of cell growth above normal.
Cu is required for VEGF expression 21 and is also able to increase VEGF production. 22,23 However, the mechanisms underlying these two actions are different. It has been known that HIF-1α is the critical transcription factor for VEGF expression. The activity of HIF-1α is mainly regulated by its intracellular stability and accumulation. 25–27 Metals such as cobalt, 23,28,29 nickel 23,29 and Cu 22,23 can inhibit the process of HIF-1α degradation, thus leading to its accumulation and activation. Therefore, cells exposed to high levels of cobalt, nickel or Cu leads to over-production of VEGF. 22,23,28,29 On the other hand, the activation of HIF-1α transcription activity requires Cu. 21 Cu depletion results in suppression of HIF-1α activation as well as inhibition of VEGF expression. 1,21
Most studies have used high levels of Cu to examine its effect on VEGF production and cell growth, 7–12 leading to a conclusion that Cu stimulation of cell growth is mediated by VEGF production. However, the present study made a clear distinction between stimulation of cell growth by physiologically relevant levels of Cu and that by Cu overload. Under the present experimental condition, Cu did not increase VEGF expression, but Cu was clearly required for VEGF expression because Cu chelation by TEPA inhibited VEGF expression. Corresponding to this effect was the inhibition by TEPA of normal cell growth, an effect that could be reversed by excess Cu. Therefore, under physiological condition, Cu is required for VEGF expression to maintain normal cell growth.
At the level of Cu that did not increase VEGF production, eNOS production was increased. This increase was likely to be responsible for Cu stimulation of cell growth under the present experimental condition. Gene silencing of eNOS did not affect normal cell growth, but suppressed the reversal by Cu of TEPA-inhibited cell growth. This is an interesting observation. It suggests that change in Cu concentration is related to eNOS production. Addition of physiologically relevant levels of Cu increased eNOS production and eNOS production was suppressed in cultures treated with TEPA. The addition of excess Cu not only reversed the suppression but also further increased eNOS production in the presence of TEPA. Therefore, this effect is Cu-dependent.
It is important to note that both genes for eNOS and VEGF are under the regulation of HIF-1; Cu is required for HIF-1 transcriptional activation 21 and high levels of Cu induce HIF-1α accumulation in the cell. 22,23 Therefore, the data obtained here would suggest that the increase in eNOS transcription results from the effect of Cu on mechanisms other than the regulation under HIF-1. In future studies, how Cu increases eNOS production in the endothelial cells, will be determined.
In conclusion, Cu at a physiologically relevant level stimulated growth of HUVECs in cultures. This stimulation was not accompanied by increases in VEGF production and anti-VEGF antibody neutralization did not inhibit Cu stimulation of cell growth; thus, the cell growth stimulating effect of Cu was not VEGF-dependent. On the other hand, Cu at the same level increased eNOS production and gene silencing of eNOS inhibited Cu recovery of TEPA inhibition of cell growth, indicating that the Cu effect is eNOS-dependent. This observation is in contrast to current belief that Cu stimulation of cell growth is VEGF-dependent, and provides novel insights into Cu promotion of angiogenesis.
Footnotes
ACKNOWLEDGEMENTS
The authors thank Xiaohe Chen for technical support. This work was supported by Science Foundation of Sichuan Province (2009HH0016; 09ZQ026-081) and by US National Institutes of Health (HL63760 to YJK).