
Abstract
We investigated differences in the pathogenesis of dextran sulfate sodium (DSS)-induced colitis between two inbred rat strains, Wistar King A Hokkaido (WKAH) and Dark Agouti (DA) rats, to determine the intrinsic factors responsible for the development of colitis. DSS exposure exacerbated the clinical symptoms such as body weight loss, stool consistency and rectal bleeding in DA rats rather than that in WKAH rats. Additionally, the average survival was shorter in DA rats than in WKAH rats. The expression levels of tumor necrosis factor-α, interleukin (IL)-12 p35 and IL-23 p19 increased prominently in the DA rats that were administered DSS, accompanied by severe infiltration of leukocytes into the colon. We also found that colonic permeability was greater in the DA rats than in the WKAH rats. In Ussing chambers, exposure of the isolated colon tissue to DSS enhanced the colonic permeability of both strains. Immunoblot analysis revealed that the expression levels of tight junction (TJ) proteins were modulated during DSS administration. Higher expression levels of claudin-4 and junctional adhesion molecule-A proteins were observed in DA rats than in WKAH rats, even in intact conditions. These results indicated that the expression pattern of TJ proteins determines the colonic permeability of the rats. In conclusion, the intrinsic colonic permeability is one of critical factors responsible for the susceptibility of rats to colitis.
Introduction
Many morphological and pathophysiological features in dextran sulfate sodium (DSS)-induced colitis are similar to those observed in human ulcerative colitis, such as ulceration, loss of barrier function and the production of inflammatory cytokines. 1,2 Barrier destruction is often observed during the development of inflammation in the intestines. 3 Likewise, DSS increases the mucosal permeability of rats 4 and reduces transepithelial resistance (TER) in the Caco-2 cell monolayer, 5 although the mechanism of DSS-induced colitis is not fully understood.
A well-known cellular component responsible for the mucosal barrier is the tight junction (TJ) among epithelial cells. The TJ is assembled from a variety of proteins, including some transmembrane proteins, such as occludin, 6 claudins, 7 junctional adhesion molecule (JAM) 8 and the intracellular plaque proteins, such as zonula occludens. 9,10 In addition, circumferential contraction and subsequent tension in the apical perijunctional actin–myosin ring regulate solute transport via the paracellular route. 11 The myosin regulatory light chain II (MLC2) and its phosphorylation trigger the contraction of the actin–myosin ring, resulting in the activation of paracellular transport. 12 Some reports suggest that dysfunctions of the intestinal barrier are associated with the pathogenesis of inflammatory bowel disease (IBD). 13–15 For example, a high level of MLC2 phosphorylation has been demonstrated in IBD patients. 13 The expression levels of claudin-1 and claudin-2 positively correlate with inflammatory activity in IBD. 14
The proliferation of epithelial cells is not only an important step in the formation of the barrier compartment but is also responsible for the restoration of epithelial tissue after inflammation. 16,17 The crypt epithelial cells (CECs) proliferate rapidly, and their turnover is completed within three days. 18,19 Such continuous epithelial proliferation is indispensable for the regeneration of tissue, even under conditions of damaged tissue, such as IBD. 16 Several cytokines produced by immune cells are involved in the restoration of epithelial tissue. For example, interleukin (IL)-1 and IL-2 have been demonstrated to enhance the migration of epithelial cells through a transforming growth factor (TGF)-β-dependent pathway. 20,21 Interferon (IFN)-γ inhibits enterocyte migration by preventing gap-junction communication among epithelial cells. 22
Our previous study indicated that the aberrant crypt foci that are induced by both chemical carcinogen and γ-irradiation in the Dark Agouti (DA) rats are significantly fewer in number than in the Wistar King A Hokkaido (WKAH) rats. 23 This result demonstrates that the DA rats are resistant to DNA damage-induced carcinogenesis and suggests that the mode of maintenance of intestinal homeostasis is different between the WKAH and DA rats. It is possible that comparing the responses to DSS of these strains is useful in evaluating the importance of the intrinsic intestinal barrier. Actually, among the inbred mouse strains, Balb/c, C3H/HeJ and C57BL/6J are relatively susceptible to DSS, whereas DBA/2J mice are mostly resistant to DSS. 24,25 Although alterations of the barrier function in intestinal inflammation have been widely investigated, the role of the intrinsic intestinal barrier in IBD development has not been thoroughly elucidated. Accordingly, we investigated the difference in pathogenesis, especially examining the intestinal barrier and immune response, between the WKAH and DA rats in the course of DSS-induced colitis.
Materials and methods
Rats and diet
Four-week-old male WKAH/HkmSlc (WKAH) and DA/Slc (DA) rats (Japan SLC, Tokyo, Japan) were housed individually in stainless-steel cages with wire-mesh bottoms. The cages were located in a room with a controlled temperature (22 ± 2°C), relative humidity (40–60%) and lighting (lights on from 08:00–20:00) throughout the day. The diet was as follows: 602.5 g/kg of sucrose, 250 g/kg of casein, 50 g/kg of microcrystalline cellulose (PH-102; Asahi Kasei Co, Tokyo, Japan), 50 g/kg of soybean oil, 2.5 g/kg of choline bitartrate, 35 g/kg of American Institute of Nutrition (AIN)-93G mineral mixture 26 and 10 g/kg of AIN-93G vitamin mixture. 26 The rats had free access to deionized water and the AIN-93G-based diet through the experimental period. All of the experiments were approved by the Hokkaido University Animal Committee, and the rats were maintained in accordance with the guidelines of Hokkaido University for the care and use of laboratory animals.
Survival test in DSS-induced colitis
To induce colitis, rats were given 4% DSS (wt/vol; MW 36,000–50,000; MP Biomedicals, Tokyo, Japan) in deionized water ad libitum after 10 days for adaptation. 1 The rats were weighed daily and were visually inspected for rectal bleeding and diarrhea. The number of surviving rats was scored every eight hours.
Myeloperoxidase activity in colonic mucosa
In a separate experiment, WKAH and DA rats (n = 5) were supplied deionized water or 4% DSS ad libitum for four days after a 10-day adaptation. The myeloperoxidase (MPO) activity was determined, as previously described. 27,28 The absorbance of the reaction mixture was measured at 655 nm. The MPO activity was expressed as the initial velocity of increasing absorbance at 655 nm [ΔA 655/(min × g mucosa)].
5-Bromo-2′-deoxyuridine incorporation
5-Bromo-2′-deoxyuridine (BrdU) immunostaining was performed according to the previously described method. 28 We scored the total number of CECs and the BrdU-incorporating CECs in a half-crypt section and analyzed in 50 different crypts for each sample, as previously described. 29 The peak position of BrdU-incorporating CECs was calculated.
Expression of mRNA in the colonic mucosa
Total RNA was isolated from the scraped colonic mucosa by RNeasy mini kit (QIAGEN, Tokyo, Japan) according to the manufacturer's instructions and converted to cDNA using random hexamer primer (Toyobo Biologics, Inc, Osaka, Japan) and Reverscript I (Wako Pure Chemical Industries, Ltd, Osaka, Japan). The cDNA templates were filtered with MicroSpin S-300 (GE Healthcare Co, Tokyo, Japan) before quantitative realtime polymerase chain reaction (PCR) was conducted. The expression of cytokine mRNA was quantified in triplicate by the ABI PRISM 7000 system (Life Technologies Japan Co, Tokyo, Japan) using TaqMan Gene Expression Assays as follows: tumor necrosis factor-α (TNF-α) (assay ID: Rn00562055_m1), IL-12 p35 (assay ID: Rn00584538_m1), IL-12 p40 (assay ID: Rn00575112_m1), IL-23 p19 (assay ID: Rn00590334_g1), IL-1β (assay ID: Rn00580432_m1), IL-2 (assay ID: Rn99999181_m1), IL-4 (assay ID: Rn99999010_m1), IL-6 (assay ID: Rn01410330_m1), IL-10 (assay ID: Rn99999012_m1), IL-18 (assay ID: Rn00824548_m1), CX3CL1 (assay ID: Rn00593186_m1) and IFN-γ (assay ID: Rn00594078_m1). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (assay ID: Rn99999916_s1) was employed as an endogenous control. The PCR consisted of one 10-min cycle at 95°C, followed by 45 cycles of 15 s each at 95°C and one minute each at 60°C (annealing and extension). For each sample, the relative expression of each mRNA was compared with that of GAPDH according to the standard-curve method. 28
Cytokine production from isolated mesenteric leukocytes
Leukocytes were isolated from mesenteric lymph nodes, as previously described.
30
The viability of mesenteric leukocytes (MLs) was assessed immediately after their isolation by trypan blue exclusion. The MLs (1.0 × 106 cells/well) were stimulated with various concentrations of DSS (0.04, 0.4, 2 or 4% wt/vol as final concentrations) in a 48-well plate for 24 h, after which the media, including the cells, were collected. After centrifugation at 400 ×
Permeability of colonic tissues
The colonic flat sheets for Ussing chambers were prepared as previously described. 28 After 10 min of preincubation at 37°C, the Hank's balanced salt solution (HBSS) in the mucosal space was exchanged for lucifer yellow (LY; 100 μmol/L; Sigma-Aldrich Japan Co, Tokyo, Japan)-containing HBSS with or without DSS (4% wt/vol as final concentration). The colonic tissues were further incubated for 120 min at 37°C. After the incubation period, the LY concentration in the serosal solution was measured using a fluorescent spectrophotometer (CAF-110; JASCO International Co, Ltd, Tokyo, Japan) at 430 nm for excitation and 530 nm for emission. The level of LY flux was calculated as nmol/cm2 surface area per one hour. The TER in each colonic segment was also measured using the Millicell electrical resistance system (Millipore, Bedford, MA, USA).
Isolated colonic sack assay
Rats were killed under pentobarbital anesthesia, and their colons were immediately excised. The colons were washed with HBSS to remove the contents and were tied at both ends. Next, 10 mmol/L of fluorescein isothiocyanate (FITC)-dextran 40,000 (MP Biomedicals) in HBSS, with or without 4% DSS (Sigma-Aldrich), was introduced into the isolated colonic sack (approximately 2 cm in length). After incubation for 30 min at 37°C in a 20-mL conical flask with 5 mL HBSS, the colon was immediately washed with HBSS, embedded into an optimal cutting temperature compound and frozen in liquid nitrogen. The frozen colonic sections (7 μm) were dried in air and fixed in cold acetone at −20°C for 24 h. Next, the sections were washed three times with PBS (pH 7.4) for five minutes. Finally, the sections were immersed in a mounting medium containing 4′,6′-diamidino-2-phenylindole (DAPI; ProLong®Gold Antifade Reagent, Invitrogen), and the fluorescence images were recorded using a fluorescence microscope (DMI6000B; Leica Microsystems, Wetzlar, Germany) and Leica FW4000 software (Leica Microsystems). Images are depicted at ×200 magnification.
Immunoblot analysis
Whole colonic mucosal extracts were prepared, as previously described. 31 In brief, the scraped colonic mucosa (50 mg) was lysed with 500 μL of lysis buffer (0.3% [wt/vol] sodium dodecyl sulfate [SDS] in 10 mmol/L Tris [pH 7.4]) containing protease inhibitors (5 μg/mL aprotinin from bovine lung, 3 μg/mL leupeptin hemisulfate and 5 mmol/L benzamidine hydrochloride) and phosphatase inhibitors (5 mmol/L imidazole, 10 mmol/L sodium fluoride, 2.5 mmol/L sodium molybdate dehydrate, 5 mmol/L sodium tartrate dihydrate and 2 mmol/L sodium orthovanadate). The protein concentrations in the lysates were measured using the bicinchoninic acid method (Pierce Biotechnology, Inc, Rockford, IL, USA). The lysates were mixed with Laemmli sample buffer (6% [wt/vol] SDS, 30% [vol/vol] glycerol, 15% [vol/vol] 2-β-mercaptoethanol and 0.02% [wt/vol] bromophenol blue in 187.5 mmol/L Tris, pH 6.8) 32 and heated at 100°C for five minutes. The samples, consisting of 50 μg of protein, were separated by SDS-polyacrylamide gel electrophoresis (12%) and transferred to polyvinylidene difluoride membranes (Bio-Rad Laboratories, Inc, Hercules, CA, USA). The membranes were blotted for various TJ proteins, such as claudin-1 (Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA), claudin-2 (Invitrogen), claudin-4 (Invitrogen), claudin-7 (Santa Cruz Biotechnology), occludin (Invitrogen), JAM-A (Invitrogen), villin (Santa Cruz Biotechnology), β-actin (Sigma-Aldrich), GAPDH (Abcam Inc, Cambridge, MA, USA), MLC2 (Cell Signaling Technology, Inc, Danvers, MA, USA) and phosphorylated MLC2 (pMLC; Cell Signaling Technology) using specific antibodies in combination with horseradish peroxidase-conjugated anti-mouse, anti-rabbit or anti-goat IgG antibodies depending on the species of the primary antibody. GAPDH was employed as a loading control. The blots were developed using the enhanced chemiluminescence method (GE Healthcare). Quantification was achieved by densitometric analysis of specific bands on the immunoblots using Image J software (National Institutes of Health, Bethesda, MD, USA).
Claudin-4 staining
Colonic frozen sections were fixed in cold acetone (−20°C) for 30 min, and washed three times with Tris-buffered saline Tween-20 (TBST; 20 mmol/L Tris, 150 mmol/L NaCl, 0.5% Tween 20, pH 8.0) for five minutes. The sections were immersed in 3% bovine serum albumin for one hour, and incubated for 1.5 h with mouse monoclonal anti-claudin-4, followed by incubation for one hour with AlexaFluor 546-conjugated anti-mouse IgG antibodies (Invitrogen). Finally, the sections were immersed in a mounting medium containing DAPI. The image was visualized using a fluorescence microscope (IX81; Olympus, Tokyo, Japan) and DP Controller software (Olympus).
Statistics
Statistical analysis was performed with JMP software (version 5.0; SAS Institute, Inc, Tokyo, Japan). The DSS survival rates were analyzed by the Mann-Whitney U test. The Student's t-test was employed for the comparison of cytokine mRNA expression levels. Two-way analysis of variance (ANOVA) was performed to analyze body weight gain, food intake, colon length, MPO activity, the number of CECs in a half crypt section, the BrdU-incorporating ratio, the peak cell position of BrdU-incorporating CECs, immunoblots, the differences in cytokine production, LY permeability and the changes in TER. Differences were considered significant at P < 0.05. All values are represented as the mean ± SEM.
Results
Changes in survival during DSS-induced colitis
The average length of survival in the DA rats was significantly shorter than that observed in the WKAH rats (P < 0.01, Figure 1a). The levels of body weight and food intake were, on average, lower in DA rats than in WKAH rats (Figures 1b and c), regardless of DSS treatment. On day 4 during the DSS treatment period, the level of food intake in the DA rats decreased to 55.1% compared with that observed on day 0 (Figure 1c). In contrast, the reduction of food intake was not as pronounced (30%) in WKAH rats. During the DSS treatment period, water consumption in the DA rats did not differ from that observed in the WKAH rats (data not shown). Additionally, no difference in occult bleeding and stool consistency was observed between the strains during the experimental period. The administration of DSS shortened the colon length in both strains, and the colon length was significantly shorter in DA rats than in WKAH rats (Figure 1d). An increase in MPO activity was observed in the DSS administration period (Figure 1e).
The development of colitis in inbred rat strains in response to DSS. (a) Survival rates in the WKAH (dashed line) and DA (solid line) rats in DSS-induced colitis. The average lengths of survival were significantly different between the strains (Mann-Whitney U test, P < 0.05, n = 10). (b) Body weights. (c) Food intake. (d) Colon length during DSS treatment period (n = 5). (e) MPO activity in the distal colon during the DSS treatment period (n = 5). Values are expressed as the means ± SEM. Two-way ANOVA P values are shown for the body weight, food intake, colon length and MPO activity (day and strain). WKAH, Wistar King A Hokkaido; DA, Dark Agouti; DSS, dextran sulfate sodium; MPO, myeloperoxidase; ANOVA, analysis of variance; NS, non-significant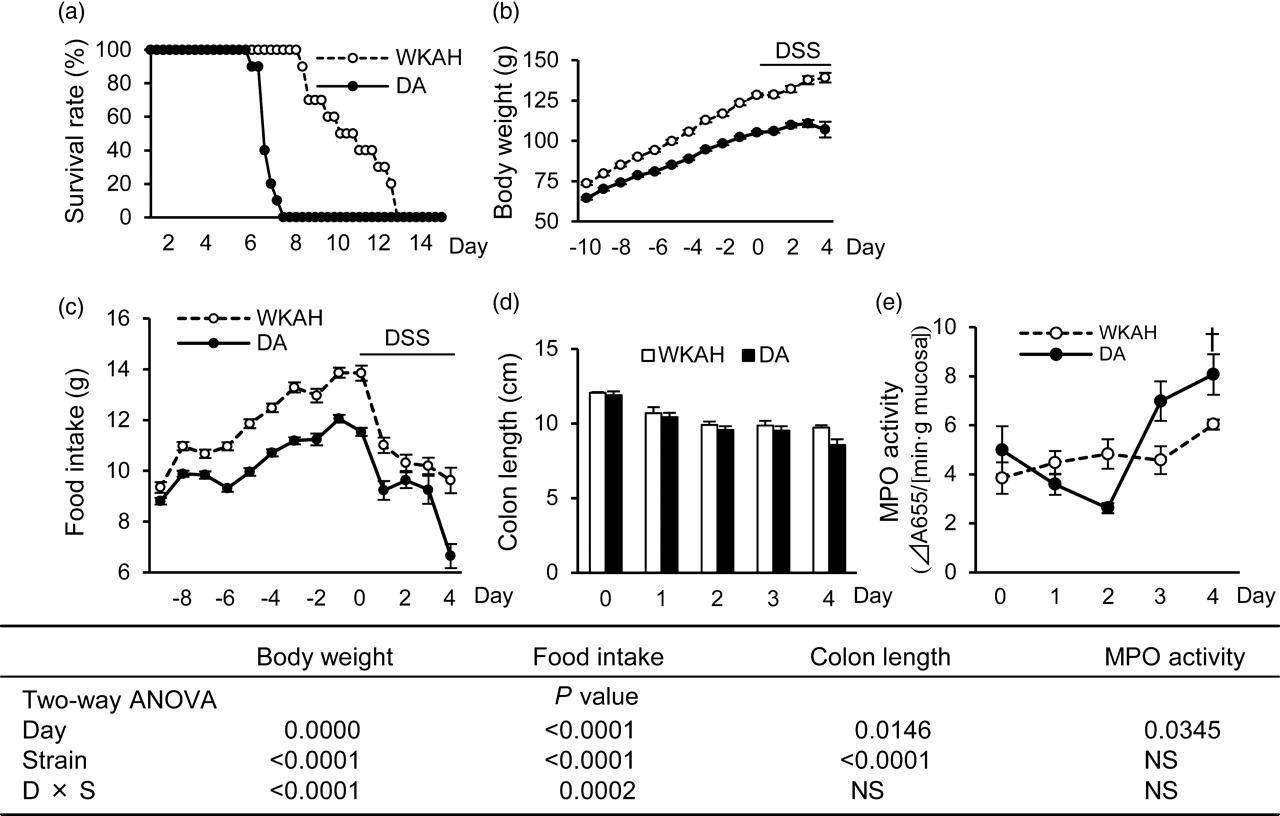
Changes in CECs during DSS-induced colitis
Peak cell position of BrdU-incorporating CECs in colon*
BrdU, bromodeoxyuridine; CEC, crypt epithelial cell; ANOVA, analysis of variance; ND, not detected
*Values are calculated from the data of BrdU distributions in CECs (Supplemental Figure 1) and expressed as the means ± SEM (n = 4–5). Two-way ANOVA P values for the peak cell position of BrdU-incorporating CECs except for that of day 4 (day and strain) are 0.0004 for day, and 0.0216 for the interaction between day and strain. No significant difference is observed for strain
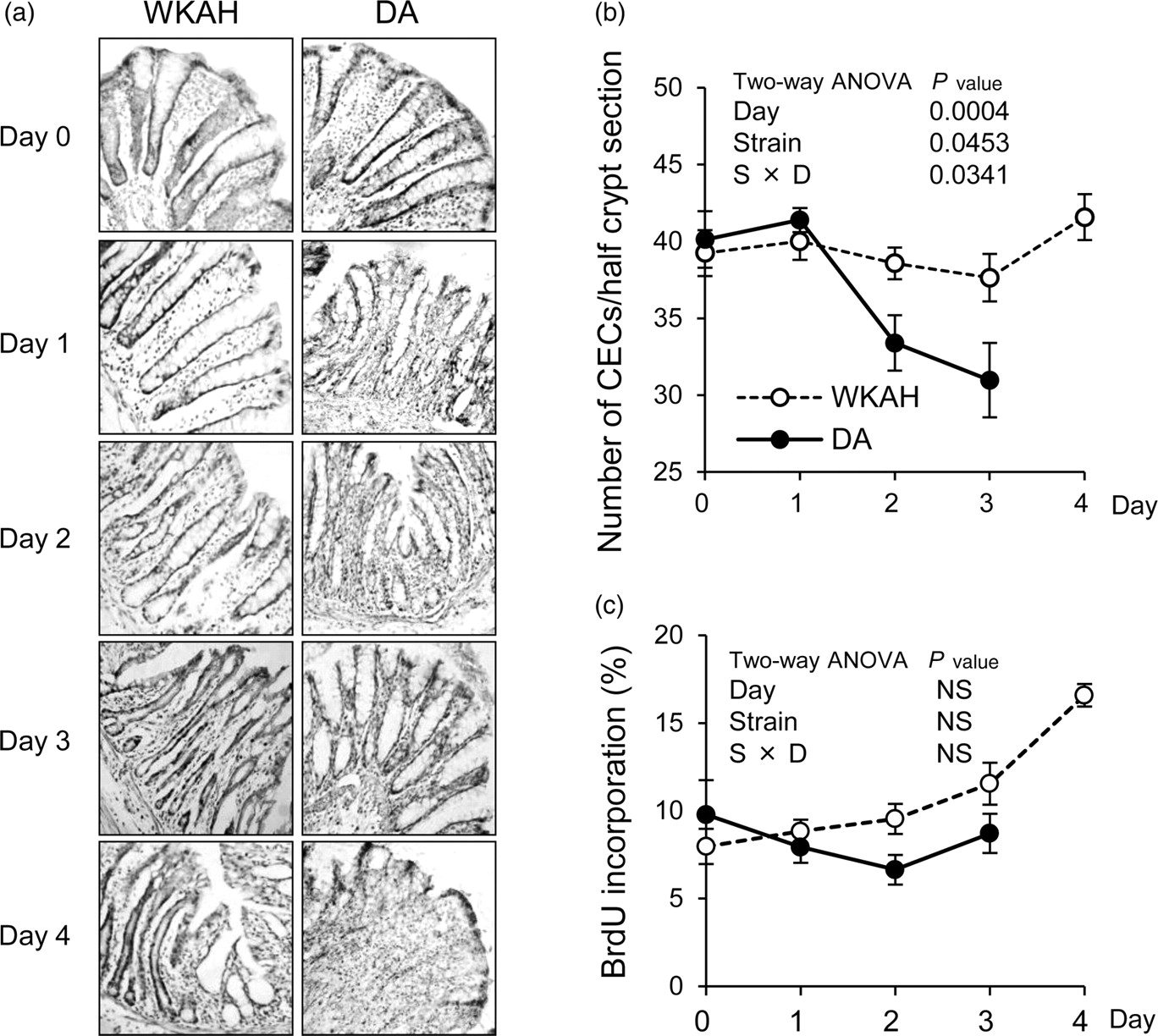
Histological observations in DSS-induced colitis. (a) Frozen section of colon stained with anti-BrdU antibody and hematoxylin. Images are depicted at ×200 magnification. (b) Changes in the number of CECs in both strains. (c) Changes in the percentages of BrdU-incorporating CECs. On day 4, we could not count the number of CECs due to the severe damage only in DA. Values are expressed as the means ± SEM (n = 5). Two-way ANOVA P values are shown for the number of CECs per a half crypt section, and the percentage of BrdU-incorporating CECs from day 0 to day 3 (day and strain). DSS, dextran sulfate sodium; BrdU, bromodeoxyuridine; CEC, crypt epithelial cell; ANOVA, analysis of variance; NS, non-significant; WKAH, Wistar King A Hokkaido; DA, Dark Agouti
Cytokine expression in the colonic mucosa
Interestingly, the mRNA expression levels of proinflammatory cytokines observed in rats in the absence of DSS treatment, such as TNF-α (P = 0.009), IL-12 p40 (P = 0.0372), IL-1β (P = 0.0495), IL-2 (P = 0.031) and IL-6 (P = 0.0462), were significantly greater in the WKAH rats than in the DA rats (Supplemental Figure 2; please see Cytokine expressions in an early phase of DSS-induced colitis. Messenger RNA expression levels were analyzed by quantitative reverse transcriptase polymerase chain reaction of TNF-α, IL-12p35, IL-12p40, IL-23p19, IL-1β, IL-2, IL-4, IL-6, IL-10, IL-18, CX3CL1 and IFN-γ. The relative expression levels were calculated after normalization against GAPDH. Values are expressed as the means ± SEM (n = 4–8). *Significantly different from the value in the WKAH rats on day 2, as determined by Student's t-test (P < 0.05). DSS, dextran sulfate sodium; TNF, tumor necrosis factor; IL, interleukin; IFN, interferon; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; WKAH, Wistar King A Hokkaido; DA, Dark Agouti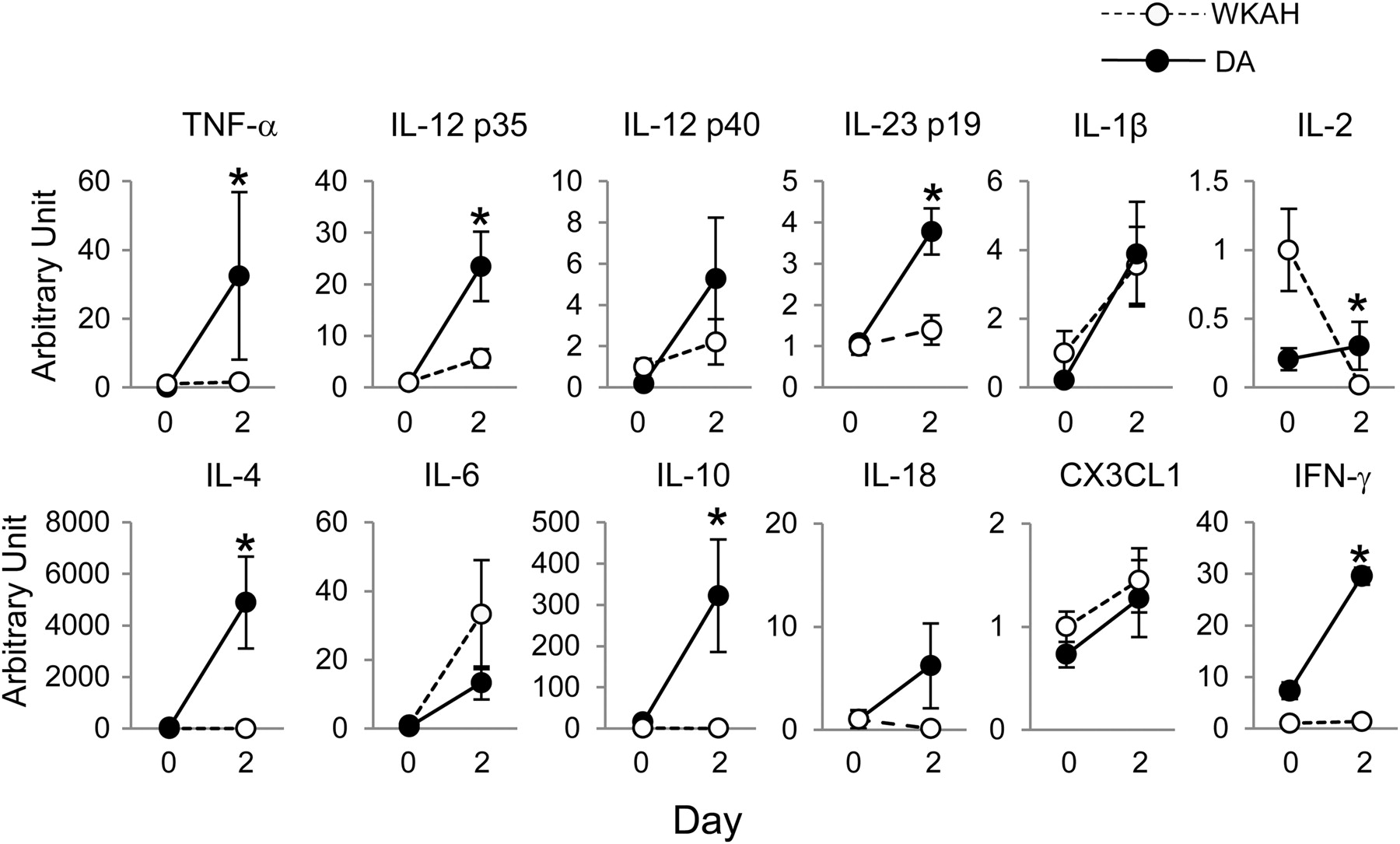
TER and colonic permeability in response to DSS
The level of LY permeability in intact colonic tissue was significantly higher in the DA rats than that in the WKAH rats, and DSS treatment in Ussing chambers promoted LY permeability in both strains (Figure 4a). Lactate dehydrogenase activity in the mucosal site of the chamber was not different between HBSS and 4% DSS treatment (data not shown). Interestingly, in colons of rats treated with DSS for two days, no increase in LY permeability was observed in response to DSS in the Ussing chambers (Figure 4b). TER gradually decreased over 120 min and also decreased in response to DSS exposure in the chambers (Supplemental Figure 3; please see Permeability in the colonic mucosa isolated from WKAH and DA rats in response to DSS in ex vivo conditions. (a) LY permeability of isolated colonic tissue from the intact WKAH and DA rats with or without exposure to DSS in Ussing chambers. (b) Representative images of FITC-dextran (MW 40,000) incorporation in colonic tissue from the intact WKAH and DA rats after incubation for 30 min with or without DSS in the sack of colon tissue. (c) LY permeability of isolated colonic tissue from the DSS-administered WKAH and DA rats with or without exposure to DSS in Ussing chambers. (d) Representative images of FITC-dextran incorporation in colonic tissue isolated from DSS-administered WKAH and DA rats after incubation for 30 min with or without DSS in the sack of colon tissue. Values are expressed as the means ± SEM (n = 6). Two-way ANOVA P values are shown for the LY permeability (strain and DSS treatment in chamber). The images are depicted at ×200 magnification. Representative images from four experiments are presented. DSS, dextran sulfate sodium; FITC, fluorescein isothiocyanate; LY, lucifer yellow; ANOVA, analysis of variance; NS, non-significant; WKAH, Wistar King A Hokkaido; DA, Dark Agouti
Incorporation of FITC-dextran in colonic mucosal cells
Using isolated colons, we found that DSS treatment enhanced the incorporation of FITC-dextran in colon mucosal cells only in the intact DA rats (Figure 4c). The DSS-induced incorporation of FITC-dextran in colons was not found in either strain of rat administered DSS beforehand (Figure 4d).
MLC2 phosphorylation in the colonic mucosa
MLC2 phosphorylation increased significantly in both stains in response to DSS treatment. No differences in the levels of MLC2 phosphorylation (Figure 5) and total MLC2 (data not shown) were observed between the strains. Although total MLC levels seem to decrease in response to DSS, the levels were not significantly different between control group and DSS group regardless of the strains.
Enhanced MLC2 phosphorylation in the colonic mucosa of the WKAH and DA rats in response to DSS exposure (upper). Representative image of the blots (lower). Quantitative analyses of the immunoblots for pMLC2 were normalized with total MLC2. Total MLC2 was not different between the strains after normalized with GAPDH (data not shown). Values are expressed as the means ± SEM (n = 6). Two-way ANOVA P values are depicted in the quantification of MLC2 phosphorylation (strain and DSS treatment). MLC2, myosin light chain 2; DSS, dextran sulfate sodium; ANOVA, analysis of variance; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NS, non-significant; p MLC2, phosphorylated MLC2; WKAH, Wistar King A Hokkaido; DA, Dark Agouti
TJ protein in the colonic mucosa
In immunoblot analysis, claudin-4 and JAM-A protein expression levels were significantly greater in the DA rats than that in the WKAH rats. In contrast, the protein expression levels of β-actin and occludin were higher in the WKAH rats than that in the DA rats (Figures 6a and b). The expression levels of the claudin-1 and claudin-7 proteins increased significantly after two days of DSS administration. The protein level of β-actin decreased in response to DSS treatment. Claudin-4 was strongly expressed in the tip of crypt in DA rats. In WKAH rats, claudin-4 was weakly expressed in the epithelium of near crypt base (Supplemental Figure 4; please see Composition of TJ proteins in the colonic mucosa of the WKAH and DA rats with or without DSS administration. (a) Representative image of the blots. (b) Quantitative analyses of the immunoblots for TJ proteins. Values of each TJ protein were normalized against that of GAPDH. Values are expressed as the means ± SEM (n = 6). Two-way ANOVA P values in the quantification in TJ proteins are presented (strain and DSS treatment). TJ, tight junction; DSS, dextran sulfate sodium; ANOVA, analysis of variance; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NS, non-significant; JAM, junctional adhesion molecule; WKAH, Wistar King A Hokkaido; DA, Dark Agouti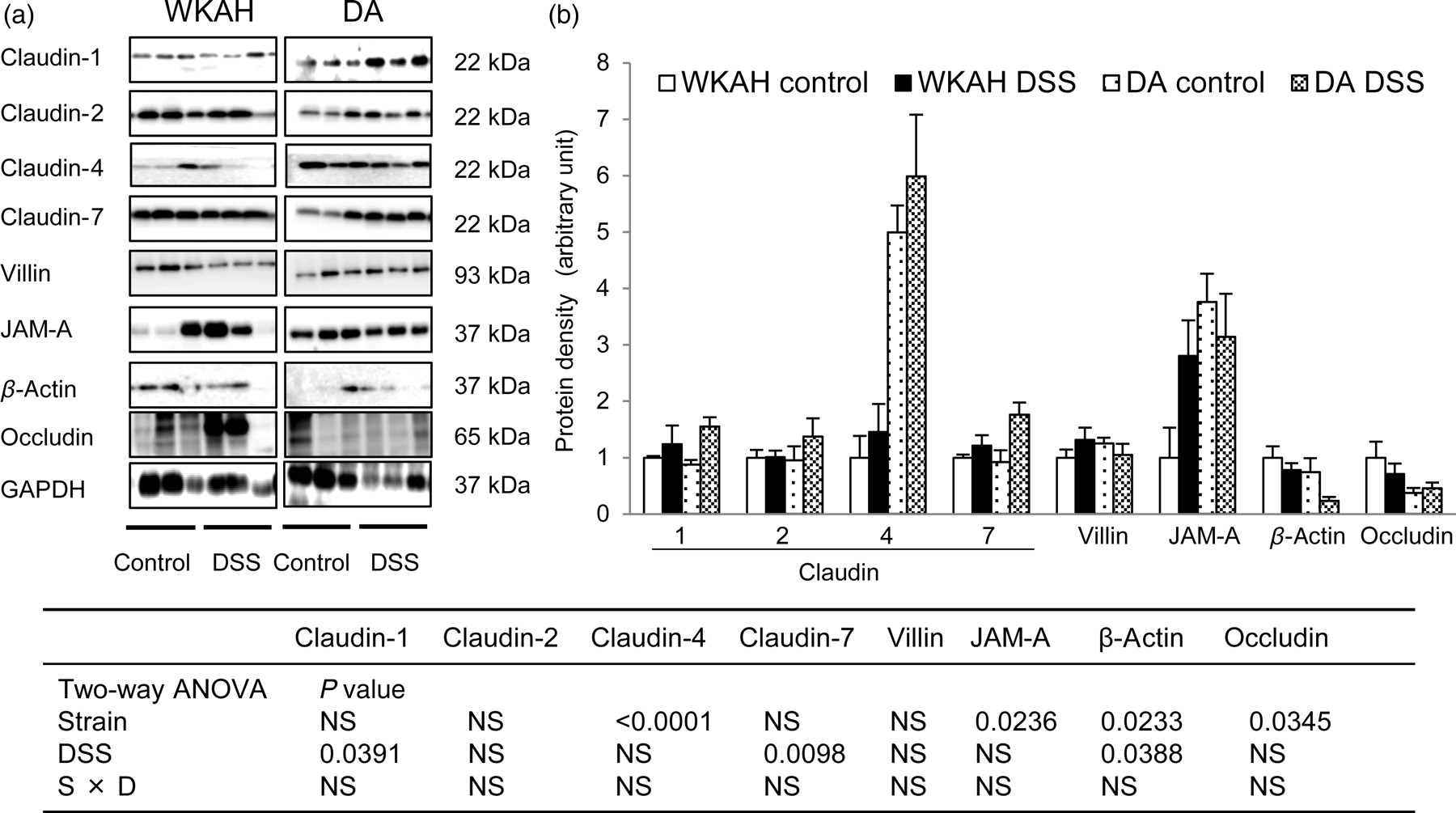
Discussion
In this study, the average survival in the WKAH rats was significantly greater than that of the DA rats after DSS administration. This result suggests differences in intrinsic factors responsible for the development of DSS-induced colitis. Previous in vivo studies have indicated that oral DSS administration increases the permeability of the colonic mucosa. 4,33 Additionally, DSS treatment in a culture study decreases the TER of Caco-2 cells. 34 In particular, 3% DSS at both 5000 and 500,000 Da exhibited a strong decrease in TER, regardless of the osmolality of the DSS solution in a culture experiment. 34 Our present study also indicates that the permeability of colonic tissue is an indispensable factor in understanding the susceptibility of rats to colitis.
Histological analyses suggest that the regeneration of CECs was enhanced in the WKAH rats but was reduced in the DA rats in response to DSS administration. It is well known that rapid epithelial cell proliferation maintains and repairs the intestinal barrier. 35 Therefore, increased CEC proliferation in the WKAH rats would contribute to the repair of the colonic injury in response to DSS, resulting in prolonged survival. However, increased epithelial cell proliferation is involved not only in a regenerative process but also in the development of neoplastic lesions. 36 In fact, our previous study demonstrated that the formation of aberrant crypt foci, a biomarker for colon carcinogenesis induced by chemical carcinogen or γ-rays, was upregulated in the WKAH rats relative to the DA rats. 29
In active IBD, a disturbed balance between regulatory and effector cells has been described, which mainly involves effector T-cells (Th1 and Th2) and Th17 cells. The Th1 cytokine profile includes IFN-γ, TNF-α and IL-12, 37 whereas the Th2-type response cytokine profile includes IL-4 and IL-10. In addition, the IL-23 is part of the immunological response in effector T-cells, and seems to be involved in IBD. Levels of expression of IL-23 and IL-17 increased in patients with active IBD. 38 A recent report demonstrated that IL-23 also plays an essential role in driving intestinal pathology in experimental models of both T-cell-dependent and innate colitis. 39 Therefore, we selected these cytokines associated with Th1, Th2 and Th17 to investigate the cause of difference in the development of colitis between WKAH and DA rats. Although Th1 and Th2 cytokine expressions were prominently increased by DSS administration in DA rats, most cytokine expression levels did not increase in WKAH rats. These results suggest that a disturbed balance between regulatory and effector cells occurred in DA rats unlike in WKAH rats. In addition, CX3CR1 plays a pivotal role for the formation of transepithelial dendrites on lamina propria dendritic cells which enable the cells to directly sample gut luminal antigens for the efficient clearance of entero-invasive pathogens. 40 Thus, CX3CL1 expression in colonic mucosa may regulate the interaction with epithelial cell and dendritic cells. Although the CX3CL1–CX3CR1 pathway also participates in the gastrointestinal mucosal immunity, we could not find any differences in CX3CL1 expression between WKAH and DA rats.
Recent reports showed that the gut-associated lymphoid tissue and mesenteric lymph nodes are a key pathogenic location of Crohn's disease.
41,42
Therefore, we focused on the differences in cytokine production in MLs. In fact, FITC-dextran, which has a similar molecular weight to DSS, was detected in the lamina propria, indicating that DSS directly stimulates MLs. Although a higher expression level of TNF-α was observed in the WKAH rats than in the DA rats after exposure to DSS in a culture study (Supplemental Figure 5; please see
Next, we investigated the factors responsible for the intrinsic permeability of epithelial tissue. MLC2 is reported to have a substantial influence on epithelial permeability, and increased MLC2 expression was reported in IBD patients. 13 However, this study indicates that MLC2 phosphorylation was not the factor responsible for determining the differences in permeability between the strains. Interestingly, an enhancement of permeability by DSS treatment in Ussing chambers was clearly suppressed when the rats were administered DSS beforehand. It is possible that DSS could not promote the MLC2 phosphorylation further.
In IBD, an increase in the expression of claudin-1 and claudin-2 is accompanied by an exacerbation of inflammation. 14 Similarly, DSS administration enhanced the expression of the claudin-1 protein, both in the WKAH and DA rats. Some cytokines have been demonstrated to modify the expression and distribution of TJ proteins. 43 For example, the combined treatment with TNF-α and IFN-γ results in the mislocalization into the cytoplasm of TJ proteins, such as JAM-A and claudin-4. 44,45 Thus, the increased levels of TNF-α, IL-1β and IFN-γ expression in response to DSS probably modified the expression levels of claudin-1, claudin-4 or JAM-A in this experiment. A prior study suggested that JAM-A knockout mice have reduced neutrophil infiltration and increased monocyte infiltration of tissues after ischemic injury. 46 A high level of JAM-A expression may promote neutrophil infiltration in the DA rats during colitis development.
Furthermore, a previous study suggests that claudin-4 expression is actually associated with loosening of intercellular junctions. 47 The tips of intestinal villi and the follicle-associated epithelium (FAE) are sites where external molecules may be incorporated. Since the FAE is an important antigen-sampling site for the initiation of immune responses, 48 looseness of its intercellular junctions may help incorporating antigens. A relatively high level of claudin-4 expression might contribute to the high permeability observed in the colons of the DA rats. In particular, claudin-4 is frequently expressed in the tip of crypt in DA rats unlike the localization in WKAH rats, indicating that the loose intracellular junction raises a chance for antigen penetration in DA rats. Hence, the varied expression and localization patterns of TJ proteins, such as JAM-A and claudin-4, are possibly responsible for the intrinsic differences in epithelial permeability observed between the WKAH and DA rats.
In summary, using Ussing chambers, we demonstrated that DSS induces an increase in colonic permeability over a short period of time. Our findings indicate that epithelial permeability in the colon is a key factor for colitis development in early phase, even in animals without any genetic modification. The expression of claudin-4 might be responsible for the leaky epithelial permeability observed in DA rats. Taken together, these results indicate that the intrinsic permeability of mucosal tissue is a factor in determining the susceptibility of rats to colitis. In other words, the mucosal barrier plays a substantial role in the prevention of colitis.