
Abstract
Morin is a flavonoid isolated from members of the Moraceae family. Morin has been reported to possess antioxidative and anticarcinogenic activities. However, the antiosteoarthritic properties of morin have not been investigated. In this study, we evaluate the antiarthritic properties of morin through in vitro and in vivo studies. We examined the effects of morin on the expression levels of matrix metalloproteinase (MMP)-3, MMP-13 and tissue inhibitors of metalloproteinase (TIMP)-1 in interleukin-1β (IL-1β)-induced rat chondrocytes by realtime polymerase chain reaction and Western blotting. The effects of morin on the phosphorylation of mitogen-activated protein kinases were also investigated. The in vivo antiosteoarthritic effects of morin were evaluated in the rat model of anterior cruciate ligament transection (ACLT)
Introduction
Osteoarthritis (OA) is a degenerative disease associated with painful joints, and is commonly a disease that occurs in older individuals. Cartilage degradation is the primary event in OA; this degradation is a result of an imbalance between matrix degradation and matrix synthesis. 1 Matrix metalloproteinases (MMPs) are considered to be the main proteases responsible for cartilage degradation. A body of evidence has demonstrated that among the MMPs, MMP-13 is particularly important in the progression of OA. 2–4
OA is widely treated with non-steroidal anti-inflammatory drugs, selective cyclooxygenase-2 inhibitors and corticosteroids, and these drugs can ameliorate the symptoms of OA. However, these agents are not ideal as they exert undesirable side-effects, 5,6 and true disease-modifying drugs do not yet exist. There is a great need to develop new agents for the treatment of OA.
Morin (3,5,7,2′,4′-pentahydroxyflavone) is a flavonoid isolated from members of the Moraceae family which have been used as dietary agents. Morin can be found in old fustic (Chlorophora tinctoria), osage orange (Maclura pomifera), almonds (P. guajava L.), mill (Prunus dulcis), fig (Chlorophora tinctoria), onion and apple and other Moraceae. It has been shown to possess antioxidative and anticarcinogenic activities. 7–9 It has also been shown to act as an anti-inflammatory agent in vitro and in vivo. 10,11 In addition, morin has been shown to inhibit MMP-2 and MMP-9 expression in the diethylnitrosamine-induced rat model of hepatocellular carcinoma. 12 However, the effects of morin on chondrocytes have not been investigated. We have undertaken this investigation in the current study.
Materials and methods
Reagents
Morin, interleukin (IL)-1β, collagenase II, dimethylsulfoxide (DMSO), 3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazoliumbromide (MTT) and Hoechst 33258 were obtained from Sigma-Aldrich (St Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM), penicillin and streptomycin, and fetal bovine serum (FBS) were obtained from Gibco BRL (Grand Island, NY, USA). Morin was dissolved in DMSO to prepare the stock solution.
Primary cell culture and treatment
The study was approved by the Institutional Animal Care and Use Committee of Zhejiang University (Hangzhou, China). Two-week-old Sprague-Dawley (SD) rats were sacrificed with an overdose of halothane. 10 Cartilage harvested from the knee joints of rats under sterile conditions was digested with 0.2% collagenase II at 37°C for four hours, as previously described, with minor modifications. 13 Cells were grown to confluence in DMEM supplemented with 10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C with 5% CO2. Cells from the third generation were used in this study. Experiments were performed after subconfluent cells were serum-starved overnight. Cells were seeded in six-well plates (1×105/well), and subconfluent cells were preincubated with various concentrations of morin for one hour, followed by stimulation with IL-1β (10 ng/mL) for 24 h. Cells were then harvested for the mRNA and protein analysis of MMP-3, MMP-13 and tissue inhibitors of metalloproteinase 1 (TIMP-1) expression levels. In another experiment, cells were preincubated with morin (1, 10 and 50 μmol/L) for 24 h, followed by stimulation with (IL)-1β (10 ng/mL) for 30 min and harvested for phosphorylation analysis.
MTT assays
Cells were cultured in a 96-well plate (6×103/well). After incubation with various concentrations of morin for 24 or 48 h in serum-free medium, MTT (5 mg/mL) was added (20 μL
Hoechst staining
To investigate the toxicity of morin on chondrocyts, we used Hoechst staining as described previously. 14 Chondrocytes were treated with morin for 24 or 48 h, stained with Hoechst 33258 (5–10 μg/mL) for 10 min and observed using a fluorescence BX51 microscope (Olympus, Tokyo, Japan). Nuclear DNA were stained as blue while apoptotic cells were identified by the presence of condensed or fragmented nuclei stained either blue or red.
Gene expression analysis
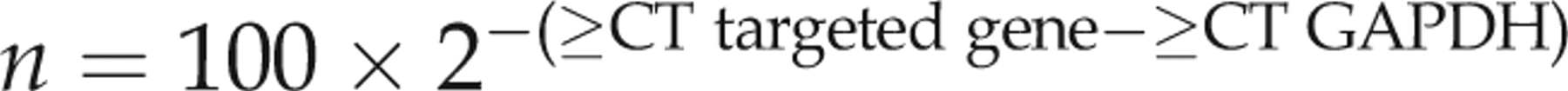
Primers of targeted genes
S, sense; A, antisense; TIMP, tissue inhibitors of metalloproteinase; MMP, matrix metalloproteinase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
Western blot analysis
Cells were rinsed with phosphate-buffered saline (PBS) and lysed with lysis buffer (50 mmol/L Tris-Cl [pH 7.4], 150 mmol/L NaCl, 1 mmol/L ethylenediaminetetraacetate [EDTA], 1 mmol/L ethylene glycol tetraacetic acid, 10 μg/mL aprotinin, 10 μg/mL leupeptin, 5 mmol/L phenylmethylsulfonyl fluoride [PMSF], 1 mmol/L dithiothreitol and 1% Triton X-100). Homogenates were centrifuged at 13,000 rpm for 15 min at 4°C and the supernatants were collected.
Proteins were resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes. After blocking for one hour with 5% milk in Tris-buffered saline-Tween, membranes were incubated with antibodies for MMP-3, MMP-13, TIMP-1, phosphorylated extracellular signal-regulated kinase-1/2 (p-ERK1/2), phosphorylated p38 (p-p38), phosphorylated c-jun amino-terminal kinase (p-JNK) and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. Membranes were incubated with horseradish peroxidase-conjugated secondary antibody at room temperature for one hour and detected using an Enhanced Chemiluminescence (ECL) kit (GE Healthcare, Shanghai, China) and exposed to X-ray films (Kodak, Hanzhou, China).
Induction of OA in rats
Fifteen eight-week-old SD rats were used in this study. OA was induced in 10 rats in the left knee joint by anterior cruciate ligament transection (ACLT) as described previously. 15 Briefly, the rat was anesthetized by 10% chloral hydrate. A medial parapatellar incision was then made, the patella was dislocated and the ACL was transected. Five rats were used as sham-operated controls; the patella was dislocated and the incisions were closed after reduction of the patella.
Rats were divided into three groups: group 1, sham-operated group (n = 5); group 2, vehicle group, rats underwent ACLT and were treated with vehicle (n = 5); and group 3, morin-treated group, rats were treated with morin (n = 5). Morin was suspended in water, and morin was administered at 50 mg/kg/d in 2 mL water after surgery as in a previous study. 10 All rats were allowed to move freely and were sacrificed 28 days after the surgery.
Gross morphology
The gross morphological changes of femur condyles were assessed in a blind manner using a grading method as follows: grade 1, intact surface; grade 2, minimal surface fibrillation; grade 3, overt surface fibrillation; and grade 4, erosion. 16
Histopathological evaluations
The left knee joints were collected for histological evaluation. Specimens were fixed in 10% neutral buffered formalin, and decalcified in EDTA. After dehydration, specimens were embedded in paraffin and cut into sections (5 μm). To assess the degree of cartilage damage, the sections were stained with Safranin O-fast green and graded according to the modified Mankin score system which was a combined score of structure, cellular abnormalities and matrix staining. In detail, the structure was defined as follows: 0 – normal; 1 – surface irregularities; 2 – pannus and surface irregularities; 3 – clefts to transitional zone; 4 – clefts to radial zone; 5 – clefts to calcified zone; and 6 – complete disorganization. Cellular abnormalities were defined as follows: 0 – normal; 1 – diffuse hypercellularity; 2 – cloning; and 3 – hypocellularity. Matrix staining was defined as follows: 0 – normal; 1 – slight reduction; 2 – moderate reduction; 3 – severe reduction; and 4 – no dye noted. 17–19
Statistical analysis
All data are expressed as mean ± standard deviation. The gross morphology data were analyzed by the non-parametric Mann-Whitney U test. MTT assay data were analyzed by an unpaired t-test while histology and gene expression data were analyzed by a paired t-test. Differences were considered significant when P was less than 0.05.
Results
Effects of morin on cell viability
Morin showed no adverse effects on cell viability (Figure 1a). Therefore, 50 μmol/L of morin was used for subsequent studies.
Effect of morin on cell viability. Cells were treated with various concentrations of morin for 24 or 48 h. The effect of morin on cell viability was evaluated by MTT assay (a) and Hoechst staining (b). Cells incubated with culture media were used to represent 100% viability. #
P > 0.05 as compared with normal chondrocytes. MTT, 3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazoliumbromide. (A color version of this figure is available in the online journal)
Hoechst 33258 staining showed that chondrocyte exposure to morin exhibited no significant morphological features of apoptosis such as bright nuclear condensation or fragments (Figure 1b).
Effects of morin on the expression of MMP-3, MMP-13 and TIMP-1 in rat chondrocytes
Realtime PCR was performed to determine the expression levels of MMPs in chondrocytes. Our results reveal that stimulation by IL-1β resulted in the induction of MMP-3 and MMP-13 gene expression, but caused a down-regulation of TIMP-1 expression. To investigate the effects of morin on gene expression, chondrocytes were preincubated with morin for one hour prior to stimulation with IL-1β for 24 h. Morin inhibited the IL-1β-mediated induction of MMP-3 and MMP-13 gene expression, and induced the expression of TIMP-1 (Figure 2a).
Effects of morin on the expression of matrix metalloproteinase (MMP)-3, MMP-13 and tissue inhibitors of metalloproteinase (TIMP)-1 in chondrocytes. Cells were pretreated with various concentrations of morin for one hour, followed by stimulation with interleukin (IL)-1β (10 ng/mL) for 24 h. Quantitative realtime polymerase chain reaction was performed to determine the expression levels of MMPs in chondrocytes (a). The protein concentrations of MMP-3, MMP-13 and TIMP-1 were detected by Western blot analysis (b). The protein concentrations of MMP-3, MMP-13 and TIMP-1 were quantitated (c). Data are expressed as mean ± standard deviation. *P < 0.05 compared with cells stimulated with IL-1β alone
We next examined the effects of IL-1β and morin on protein expression of MMP-3, MMP-13 and TIMP-1 in chondrocytes. As observed with the mRNA, treatment with IL-1β resulted in the up-regulation of MMP-3 and MMP-13, as well as the down-regulation of TIMP-1 at the protein concentration. These effects were blocked by morin (Figures 2b and c).
Morin blocks the IL-1β-mediated induction of the mitogen-activated protein kinases signaling pathway
To investigate the mechanism by which morin inhibits MMPs, we examined the effects of morin on mitogen-activated protein kinase (MAPK) phosphorylation, as MAPKs have been implicated in the regulation of MMPs in arthritis. As shown in Figure 3, IL-1β significantly induced the phosphorylation of ERK-1/2, p38 and JNK in chondrocytes. Phosphorylation of p38 and p-ERK were blocked by treatment with morin, while the level of p-JNK was not affected (Figure 3).
Effects of morin on the phosphorylation of mitogen-activated protein kinases. Cells were preincubated with various concentrations of morin for 24 h, followed by stimulation with interleukin (IL)-1β (10 ng/mL) for 30 min. Phosphorylation of extracellular signal-regulated kinase (ERK), p38 and c-jun amino-terminal kinase (JNK) were assessed by Western blot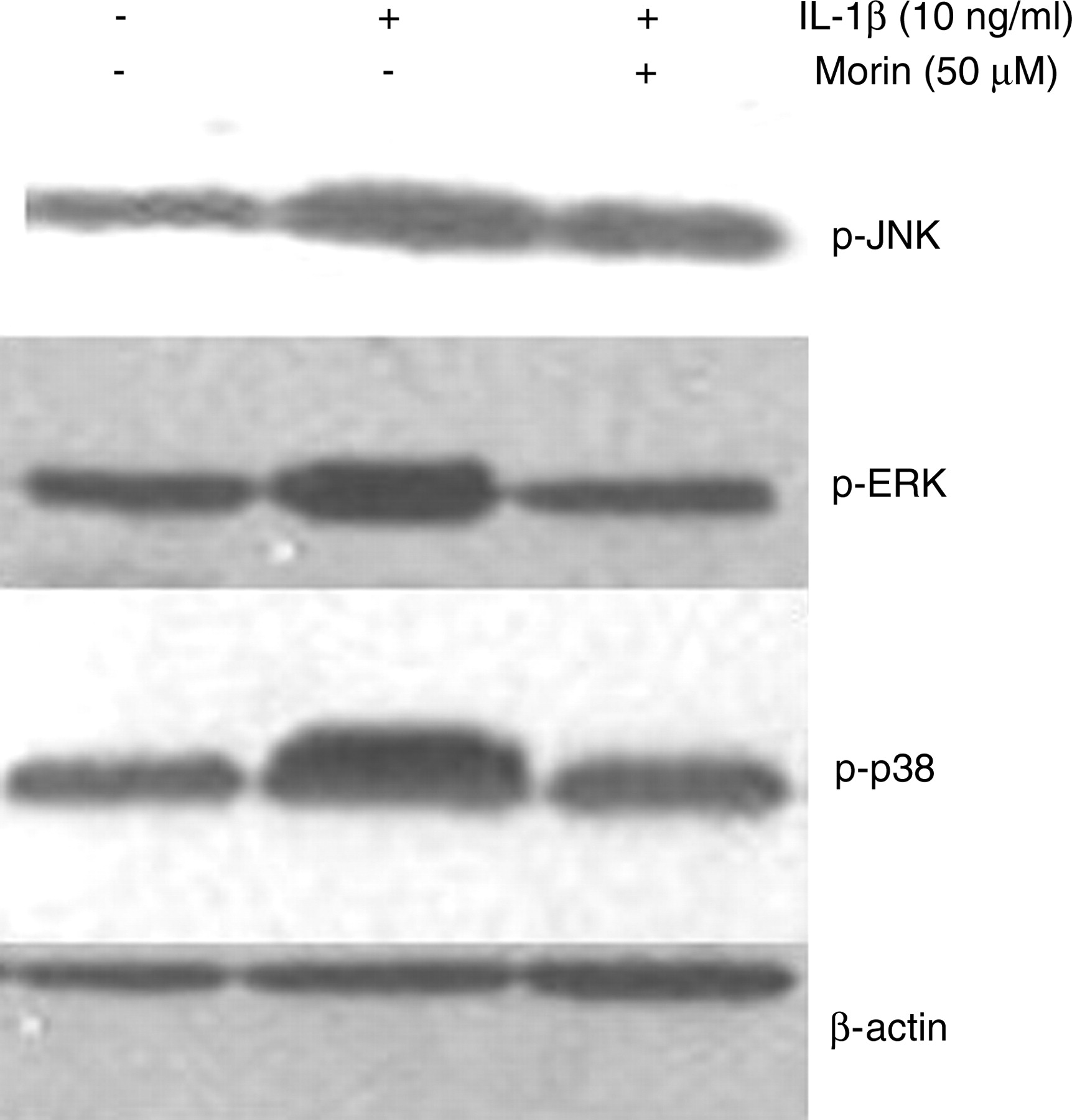
Gross morphology of articular cartilage
There were no macroscopic changes observed in cartilage from the sham-operated group. In contrast, ACLT was observed to induce the degradation of articular cartilage, and morin inhibited these degenerative changes; however, the difference between the vehicle group and the morin-treated group was not significant (Figure 4a).
The in vivo effects of morin on cartilage. After the anterior cruciate ligament transection operation, morin was orally administrated to rats daily for 28 days. Morin inhibited cartilage degradation; however, statistically significant differences were not observed (P > 0.05) (a). Representative pictures of Safranin O staining are shown (original magnification is ×50) (b). The Mankin scores are also presented. *P < 0.05 as compared with vehicle group (c). (A color version of this figure is available in the online journal)
Histopathological changes in articular cartilage
ACLT led to histopathological changes such as surface depletion and reduced Safranin O-fast green-staining in the cartilage, and morin inhibited the cartilage degradation, which developed in the progression of OA (Figure 4b). Consistent with these findings, the modified Mankin score was reduced in the morin-treated group as compared with the vehicle group (Figure 4c).
Discussion
Irreversible cartilage damage represents the pathological changes that cause OA. IL-1β, a proinflammatory cytokine, is considered to play a pivotal role in inducing these changes due to its ability to induce the production of MMPs, which lead to the breakdown of the cartilage matrix. In this study, we evaluated the effect of morin on MMP induction in rat chondrocytes, as well as its effect on ACLT-induced OA in a rat model. We demonstrated that morin inhibited the IL-1β-mediated induction of MMP-3 and MMP-13, and induced the expression of TIMP-1, a metalloproteinase inhibitor, in rat articular chondrocytes; these effects were manifested at both the mRNA and protein concentrations. The inhibitory effects of morin on MMP-3 and MMP-13 were associated with the phosphorylation of MAPK. In addition, morin administered orally at 50 mg/kg/d inhibited cartilage degradation in vivo in a rat model of OA.
MMPs are a family of proteinases that have been shown to be involved in the degradation of extracellular matrix (ECM). MMP-13, in particular, is a predominant proteinase because of its ability to cleave collagen 2, a main component of ECM. Targeting MMPs represents a potentially promising approach to the treatment of OA. MMPs can be induced by IL-1β, and the increased expression of MMPs has been observed in osteoarthritic cartilage. This is supported by the work of Bau et al., 20 who reported that the expression of MMP-13 was significantly increased in late-stage human OA cartilage, as compared with normal cartilage. Similarly, serum concentrations of MMP-3 were elevated in patients with OA, as compared with those from normal subjects. 21 In the present study, pretreatment with 50 μmol/L morin resulted in a significantly reduced expression level of MMP3 and MMP-13 in IL-1β-induced chondrocytes. Morin also induced the expression of TIMP-1 without causing cytotoxicity. Since TIMPs are the endogenous inhibitors of MMPs, we speculate that morin may modulate the balance between MMP and TIMP expression to exert its antiarthritic effects in IL-1β-treated rat articular chondrocytes.
IL-1β is known to play a pivotal role in cartilage degradation via the induction of MMPs by chondrocytes. Chondrocytes stimulated with IL-1β in vitro have been used to mimic the microenvironment that occurs in OA. 22,23 There are several signaling pathways involved in the induction of MMPs by IL-1β in chondrocytes. Among them, the MAPK signaling pathway involving ERK, JNK and p38 kinase are believed to play an important role in the induction of MMP expression. 24,25 Given this, agents that interfere with the MAPK signaling pathway may consequently affect MMP expression. 26 In the present study, we investigated whether morin affects the activation of the MAPK signaling pathway. Stimulation of cells with IL-1β (10 ng/mL) for 30 min led to significant activation of ERK, p38 and JNK, and pre-treatment with morin resulted in the inhibition of phosphorylation of ERK-1/2 and p38, while the phosphorylation of JNK was not affected. This study is the first to report the effects of morin on MAPK in chondrocytes. These findings are consistent with previous studies that described the effects of morin on MAPK in other cell systems; Kim et al. 27 reported that morin inhibited the activating phosphorylation of ERK, p38 and JNK in rat basophilic leukemia (RBL)-2H3 cells, while Hsiang et al. 28 demonstrated that morin inhibited the activation of p38 and exhibited no effects on the phosphorylation of JNK in 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced hepatocytes. The discrepancies between our findings and these reports may be related to differences in cell type and stimulation conditions.
Here we also investigated the in vivo effects of morin on cartilage degradation in an ACLT-based experimental model of OA in rats. This model has been widely used to evaluate the efficacy of agents in the treatment of OA due to the model's mechanical instability, which leads to cartilage degradation. 19,29 As shown in this study, ACLT in the rat resulted in cartilage degradation, and oral administration of morin at 50 mg/kg/d for four weeks inhibited cartilage degradation as assessed by histological evaluation. These results are consistent with the in vitro findings. These observations provide evidence that morin possesses chondroprotective activity both in vitro and in vivo.
In conclusion, the present study has demonstrated that morin possesses chondroprotective effects in IL-1β-induced rat chondrocytes and an experimental model of OA. The inhibition of MMPs by morin is partly associated with the inhibition of phosphorylation of p38 and ERK. Our results indicate that morin may show promise as a possible therapeutic agent for the treatment of OA. However, further studies are needed to confirm and extend these preliminary results.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by the National Natural Science Foundation of China (81071492).