
Abstract
We assessed the contribution of the paraventricular nucleus (PVN) in the heat stress-mediated changes in sympathetic nerve activity and blood flow redistribution from the core to the skin surface. Renal sympathetic nerve activity (RSNA), mean arterial pressure (MAP), heart rate (HR), and body and tail temperatures were recorded in anesthetized rats after bilateral microinjection of cerebrospinal fluid (CSF), lidocaine or NG-monomethyl-L-arginine (
Introduction
In order to maintain body temperature constant at ∼37°C, the balance between heat production and heat loss is accurately adjusted by coordinated autonomic outflow dictated by the central nervous system to produce the appropriate thermoregulatory responses. 1 As a result of increases in body temperature, heat loss mechanisms are initiated mainly through modulation of sympathetic activity. 2,3 During heat stress, the elevation of body temperature is proportionally followed by an increase in renal sympathetic nerve activity (RSNA), heart rate (HR) and mean arterial pressure (MAP), as well as sympathetic inhibition to the cutaneous circulation, resulting in vasodilation in the skin. 2,3 These autonomic adjustments are critical for heat dissipation through blood flow redistribution from the body core to the skin surface, as they are a consequence of sympathetically controlled vasoconstriction of the viscera, including the kidneys, and vasodilation of the skin. 4,5
The paraventricular nucleus (PVN) of the hypothalamus is a central site for the integration of sympathetic nerve activity, 6,7 and is shown to be involved in the control of body temperature. 8,9 The PVN contains thermosensitive neurons that are activated during heat stress. 10,11 In addition, there is evidence to suggest that projections arising from the PVN project to other thermoregulatory centers, like the rostral ventrolateral medulla, to influence the sympathetic activity to thermoregulatory effector organs like brown adipose tissue, vasculature of the rat's tail, as well as the kidneys and gut. 12,13 It has also been demonstrated that neuronal inhibition of the PVN with the gamma-aminobutyric acid (GABA) A agonist muscimol prevents the reduction in renal blood flow in response to increased body temperature. 4 Such evidence suggests an important regulatory role for the PVN in adjusting renal blood flow in situations of hyperthermia. However, there is no evidence suggesting a contributing role for the PVN in changes in autonomic outflow, such as RSNA and cardiovascular adjustments for heat dissipation through blood flow redistribution from the core to the skin surface in response to heat stress.
Nitric oxide, which is diffusely found in the PVN,
14
was implicated to exert thermoregulatory effects characterized by hypothermia.
15,16
Acting centrally, nitric oxide plays a tonic role in reducing body temperature due to increased heat loss through cutaneous vasodilation.
17
Under thermoneutral conditions, the injection of the nitric oxide synthase inhibitor, NG-monomethyl-L-arginine (
In order to evaluate the involvement of the PVN on the control of sympathetic activity that influences cardiovascular adjustments responsible for blood flow redistribution induced by changes in body temperature, we determined the effect of PVN blockade with lidocaine during heat stress on RSNA, MAP, HR, and body and tail temperatures. Modification in blood vessel resistance and, consequently, blood flow is mostly controlled by the sympathetic system. Therefore, RSNA was considered an indicator of sympathetic outflow to the kidneys and a reliable and consistent representation of renal vasomotor tonus and, consequently, renal blood flow. Tail temperature was measured simultaneously as an indirect index of cutaneous heat loss. The magnitude of vasodilation of the tail artery was reflected by skin temperature. 20,21 Since nitric oxide is thought to be a signaling molecule in thermoregulatory heat dissipation, as well as sympathoinhibitory, 14,19 we also investigated the contribution of endogenous nitric oxide in the PVN on autonomic adjustments induced by heat stress.
Materials and methods
Ethical approval
All procedures and experimental protocols used in this study were performed in accordance with approved protocols by the Institutional Animal Care and Use Committee of the University of Nebraska Medical Center and conformed to the guidelines for the care and use of laboratory animals of the National Institutes of Health and the American Physiological Society.
Animals
Eighteen male Sprague-Dawley rats (290–320 g body weight; Sasco Breeding Laboratories, Omaha, NE, USA) were allowed to acclimate to our animal care facility for one week before use. On the day of the experiment, each rat was anesthetized with urethane (0.75 g/kg intraperitoneally) and α-chloralose (70 mg/kg intraperitoneally), and the left femoral vein was cannulated with polyethylene (PE-50) tubing for injection of supplemental anesthesia, as required. Adequacy of anesthesia was monitored throughout the experimental procedures by absence of a withdrawal reflex, as well as by absence of a corneal reflex. The left femoral artery was also cannulated and connected to a computer-driven data-recording and -analyzing system (MacLab; AD Instruments, Mountain View, CA, USA) via a pressure transducer (model P231D; Gould, Cleveland, OH, USA) for recording of arterial blood pressure and HR. The trachea was intubated to facilitate spontaneous ventilation.
Placement of microinjection cannula in the PVN
The anesthetized rat was placed in a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA, USA). A longitudinal incision was made on the head and the bregma was exposed. The coordinates for the PVN (1.5 mm posterior to bregma, 0.4 mm lateral to the midline and 7.8 mm ventral to the dura) were determined from the atlas of Paxinos and Watson. 22 A small burr hole was made in the skull. For the microinjections, a thin needle (0.5 mm outer diameter, 0.1 mm inner diameter) connected to a microsyringe (0.5 μL; model 7000.5; Hamilton, Reno, NV, USA) was lowered into the PVN bilaterally.
Recording of renal sympathetic nerve activity
The left kidney was exposed through a retroperitoneal flank incision. A branch of the renal nerve was isolated from the fat and connective tissue. The nerve was placed on a pair of thin bipolar platinum electrodes. The nerve–electrode junction was insulated electrically from the surrounding tissue with silicone gel (Sil-Gel 604 AB; Wacker, St Louis, MO, USA). The electrical signal was amplified (10,000 times) with a Grass Technologies amplifier (model P55; Grass Technologies, West Warwick, RI, USA) with high- and low-frequency cut-offs of 1000 and 100 Hz, respectively. The output signal from the amplifier was rectified and integrated (20 ms time constant) and stored for later analysis using a computer-run data acquisition system (MacLab). The signal recorded at the end of the experiment (after the rat was killed with anesthesia overdose) was considered background noise. Nerve discharge was calculated by subtraction of the background noise from the actual recorded value. Basal nerve discharge was defined by subtraction of the background noise from the actual nerve discharge before administration of drugs into the PVN. The responses of RSNA to heat stress during the experiment were subsequently expressed as percent change from baseline.
Recording of colonic temperature and tail skin temperature
Colonic temperature was used as body temperature and was measured using a thermistor probe (model 401; Yellow Springs Instruments, Yellow Springs, OH, USA). The thermistor probe was inserted 4 cm past the anal sphincter after fecal pellets had been removed from the colon by gentle external massage. Tail temperature was measured using a probe (series 409-B; Yellow Springs Instruments) taped to the dorsal surface of the skin, 10 mm from the base of the tail. The tail skin temperature provides reliable measurement of heat transferred by the vasodilating blood vessels to the surface. The parameters mentioned above were used to determine body temperature threshold for tail skin vasodilation, i.e. the body temperature at the moment at which tail temperature begun to increase. 20,23
Experimental protocol
After the surgical procedures, the animals were allowed to stabilize for at least 30 min before being submitted to the experimental protocol. RSNA, MAP, HR, and body and tail temperatures of the rats were recorded continuously during heat stress. Heat stress was produced in rats by increasing the temperature of a heating pad (model 3PN 1010BV; Staco Energy Products Company, Dayton, OH, USA) from 37°C to 43°C at a rate of 1.2°C every six minutes in an interval of 30 min. Heat stress was applied to the ventral side of the animal. The rats were randomly assigned to groups receiving bilateral microinjection into the PVN of artificial cerebrospinal fluid (CSF; 100 nL/side; control group), lidocaine (1%; 100 nL/side) or
Brain histology
After the experiment, the rats were killed and the brains removed and fixed in 10% formalin for at least 24 h. The brains were then frozen, and serial transverse sections (30 µm) were cut with a cryostat (−18°C). The sections were mounted on microscope slides and stained using 1% neutral red. The location of the injection within the PVN was verified under a microscope with ×40 magnification (Figure 1). The microinjections that terminated in the boundaries of the PVN were considered effective. The 100 nL injection volumes targeting the PVN would be expected to distribute the drug in or within <0.5 mm from the rostrocaudal and mediolateral boundaries of the PVN. A total of six rats were excluded from the study due to misplacement of the injections (i.e. outside the boundaries of the PVN).
(a) Schematic transverse sections of the rat hypothalamic PVN showing the center of the microinjection sites. (b) Representative photomicrograph of an injection into the PVN of a rat. PVN, paraventricular nucleus; AH, anterior hypothalamus. (A color version of this figure is available in the online journal)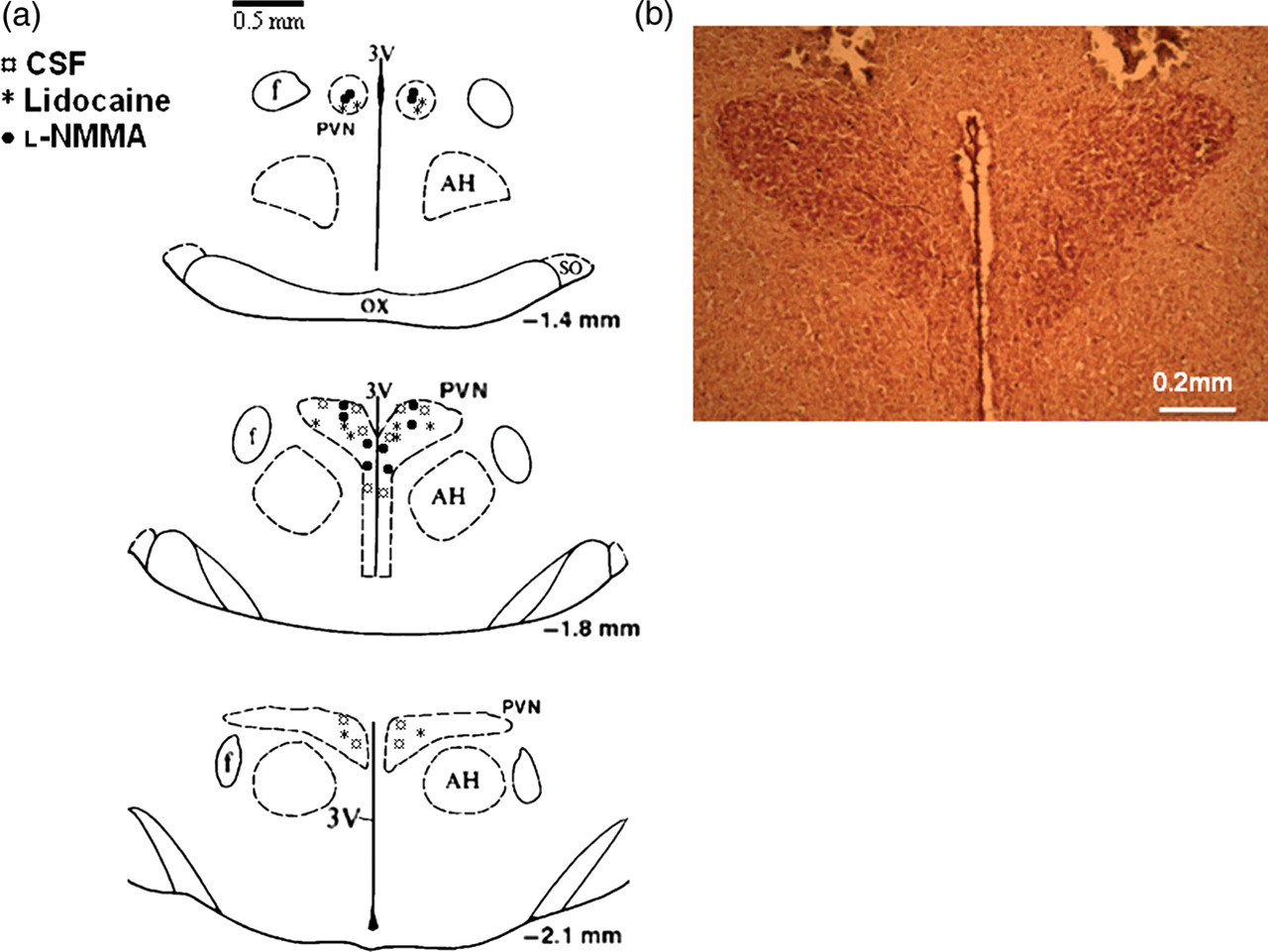
Data analysis
Responses of RSNA to heat stress are expressed as percent change from baseline. Responses of MAP, HR, and body and tail temperatures are expressed as the difference between the basal value and the value during heat stress. These data are reported as mean ± SEM and were subjected to a two-way analysis of variance, followed by the Newman–Keuls test. The body temperature threshold for tail skin vasodilation was compared using unpaired Student's t-test. The significance level was set at P < 0.05.
Results
Effect of bilateral injection within the PVN of CSF, lidocaine or
PVN, paraventricular nucleus; CSF, cerebrospinal fluid
Values are expressed as mean ± SEM
Heat stress induced an increase in RSNA response after PVN injection of CSF, starting at 6 min (Δ1.2°C) and sustained until the end of the stimulus (Figures 2 and 3a) (P < 0.05). As seen in Figure 3a, lidocaine injection resulted in blunted RSNA response throughout heat stress. Such response was significantly lower than the CSF group from 12 min (Δ2.4°C) (34.9 ± 4.8% versus 10.5 ± 6.3%, P < 0.05) of heat stress until the end of the protocol. Segments of original recordings from individual rats demonstrating the effect of heat stress on changes in heart rate (HR), mean arterial pressure (MAP) and renal sympathetic nerve activity (RSNA) after bilateral paraventricular nucleus (PVN) injection of cerebrospinal fluid (CSF) (a), lidocaine (b) and Effect of heat stress over time on changes in renal sympathetic nerve activity (ΔRSNA; a), mean arterial pressure variation (ΔMAP; b) and heart rate variation (ΔHR; b) after bilateral PVN injection of CSF, lidocaine or 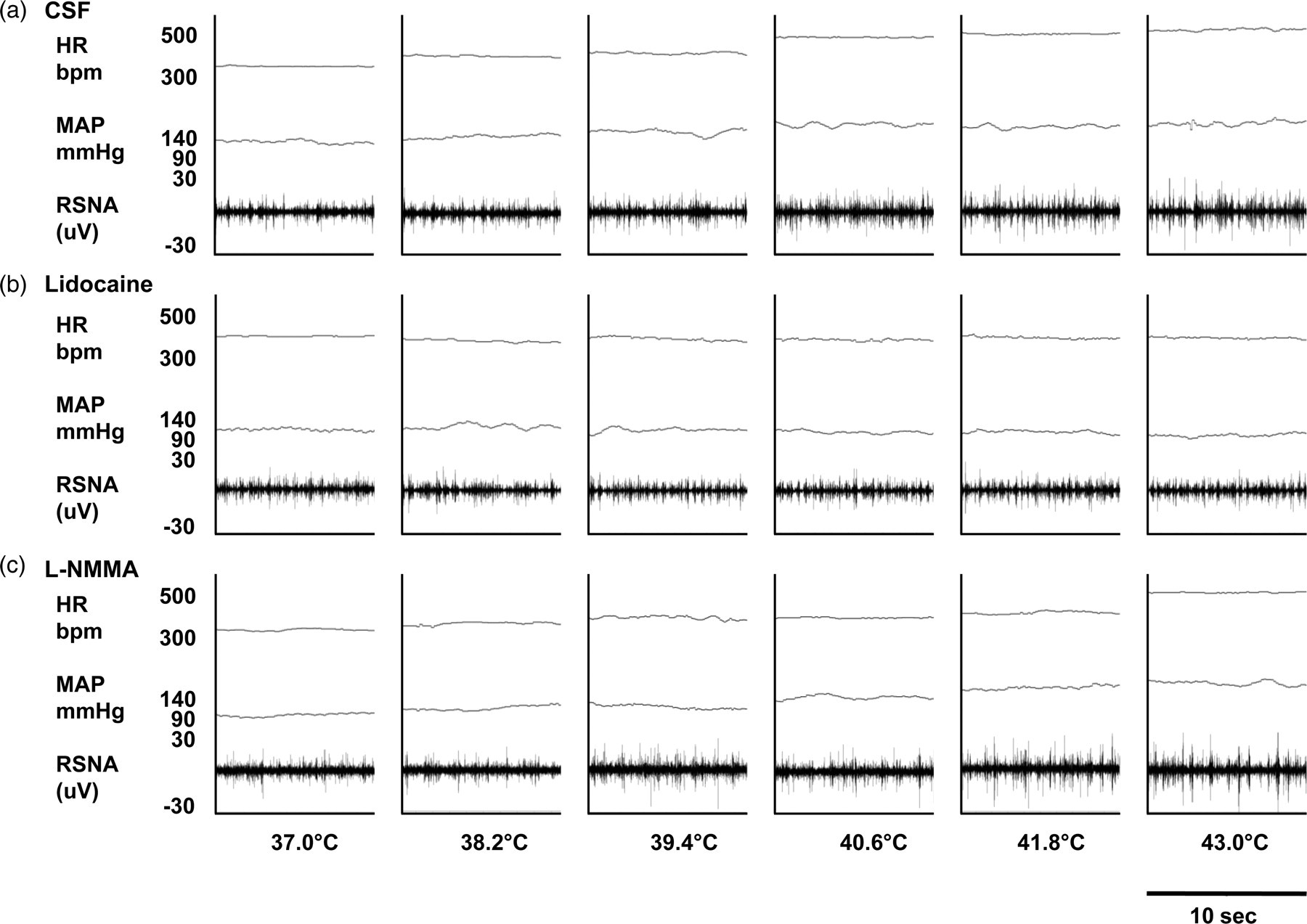
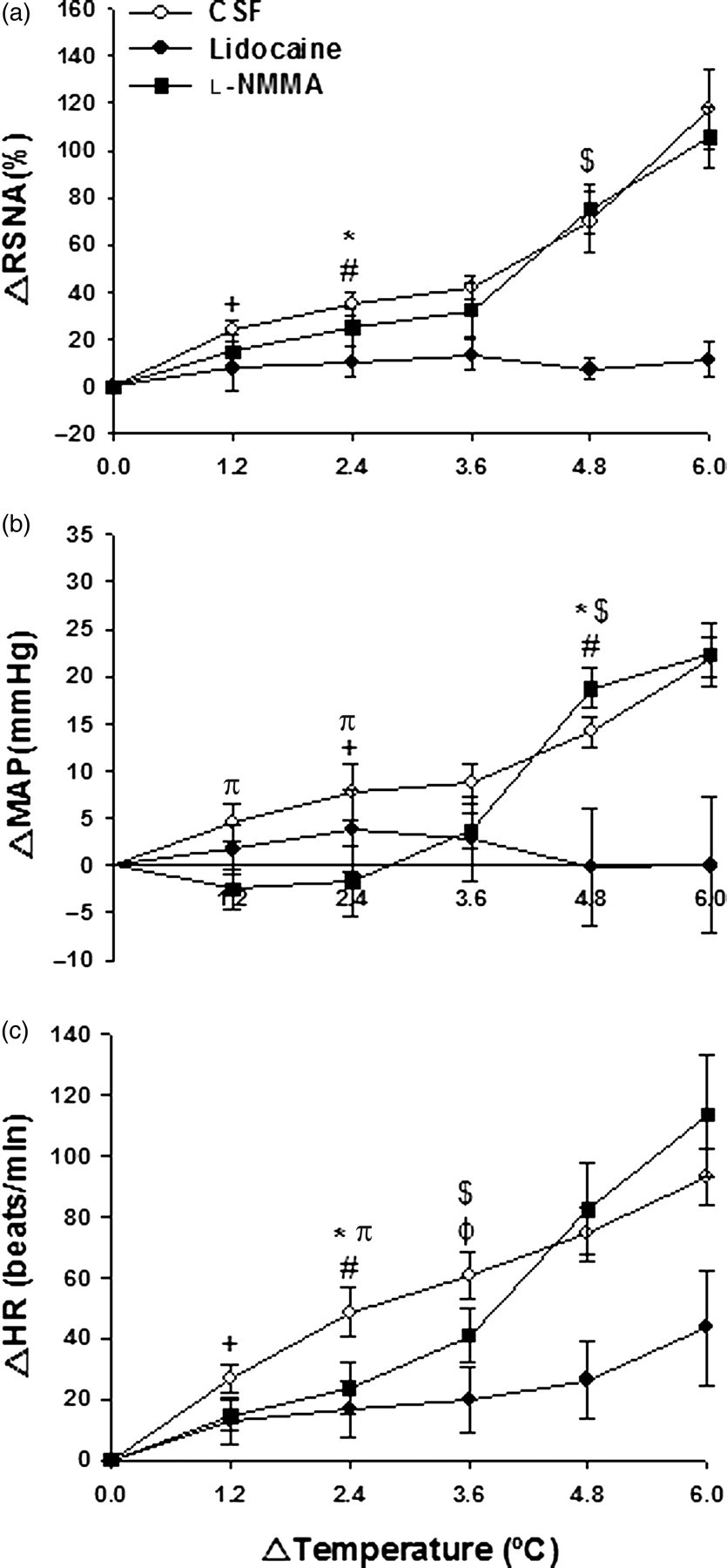
MAP, following PVN injection of CSF, increased after 12 min (Δ2.4°C) of heat stress (Figures 2 and 3b) (P < 0.05). Again, lidocaine injection resulted in a blunted MAP response throughout heat stress (Figures 3b). MAP was only significantly different between the CSF and lidocaine groups after 24 min (Δ4.8°C) (14.9 ± 1.6 mmHg CSF versus −0.2 ± 1.6 mmHg lidocaine, P < 0.05) until the end of heat stress. MAP responses in the
HR values increased progressively with elevations in temperature in all groups of animals (Figures 2 and 3c). HR was already elevated after six minutes (Δ1.2°C) of heat stress in the CSF group (Figure 3c) (P < 0.05). Although lidocaine injection induced an increase in HR after body heating for 18 min (Δ3.6°C) until the end of heat stress (P < 0.05), this increase was blunted compared with the CSF group after 12 min (Δ2.4°C) (48.5 ± 8.2 beats/min CSF versus 16.5 ± 9.4 beats/min lidocaine, P < 0.05) until the end of heat stress. Figure 3c shows that
The effect of heat stress on body temperature is shown in Figure 4a. Heat stress induced a rapid increase in body temperature in all groups, which was observed nine minutes after injection of CSF within the PVN (Figure 4a) (P < 0.05). After eight minutes of heat stress, body temperature was higher than baseline values in the lidocaine group (Figure 4a) (P < 0.05). Similar to the response observed with CSF injection, Effect of heat stress over time on changes in body temperature (a) and skin tail temperature (b) and body temperature threshold for tail skin vasodilation (c) after bilateral PVN injection of CSF, lidocaine or 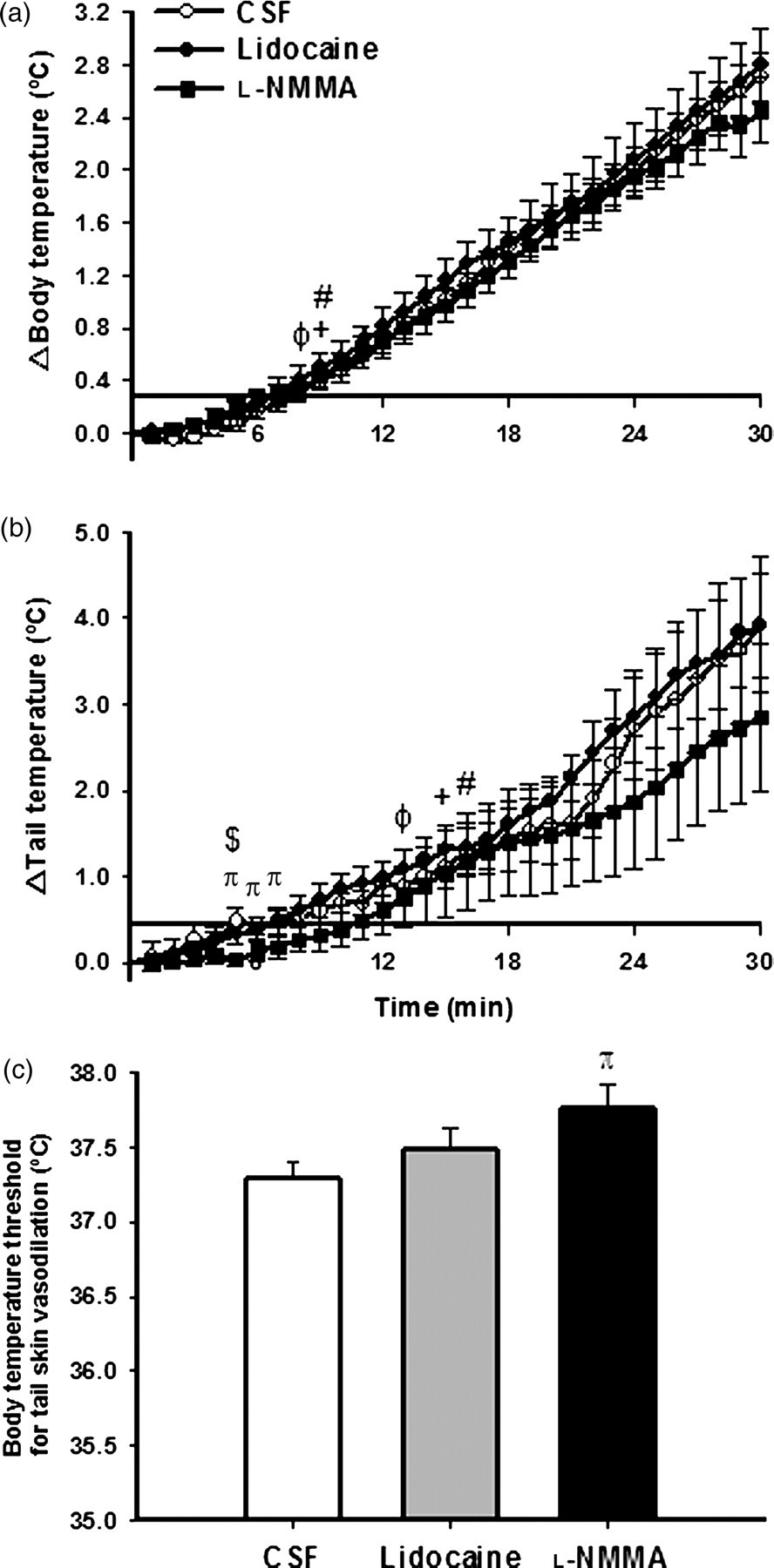
As illustrated in Figure 4b, tail temperature increased within 15, 13 and 16 min of heat stress in the CSF, lidocaine and
Discussion
In the present study, blockade of the PVN with lidocaine resulted in blunted RSNA, MAP and HR responses to heat stress, suggesting a contributing role for the PVN in autonomic responses to heat stress. However, such blockade of the PVN did not interfere with body temperature threshold for tail vasodilation. On the other hand, blockade of nitric oxide synthase within the PVN, led to higher body temperature threshold for tail vasodilation, without affecting the increase in RSNA, MAP and HR. Taken together, these data indicate that the PVN contributes to the responsiveness of sympathetic pathways that mediate autonomic adaptations related with body temperature control during heat stress, specifically by modulating RSNA that enables renal blood flow redistribution. This effect is not mediated by a nitric oxide mechanism. However, heat loss through skin vasodilation depends on a nitric oxide mechanism within the PVN.
Regulation of sympathetic outflow is necessary to facilitate heat dissipation in response to heat stress responses and to maintain body temperature within safe limits. 1,3 During heat stress, the sympathetic nervous system is capable of altering blood flow to multiple organs by selectively altering the pattern of sympathetic outflow, non-uniformly and independently, according to specific vascular beds. 2,3 It is well known that redistribution of blood flow from the viscera to the cutaneous vasculature is the major cardiovascular response for restoration of thermal homeostasis after an elevation in body temperature. This is a consequence of sympathetically controlled vasoconstriction of the viscera and vasodilation of the skin. 3,4 During heat stress, up to 60% of cardiac output is redistributed to the surface of the skin to improve heat loss, this effect being primarily dependent on decreased vascular resistance to the blood vessels in the skin. 24,25 Such vasodilation in the vasculature of the skin is initiated when the body temperature threshold for skin vasodilation is reached, and corresponds to the moment when sympathetic outflow is overcome by active vasodilation, leading to increased heat dissipation. 20,21
It has been shown that the PVN influences sympathetic outflow that controls cardiovascular responses elicited by heat stress.
4,9
These studies proposed that efferent projections from the PVN could influence sympathetic tone via connections with spinally projecting neurons, as well as with the rostral ventrolateral medulla.
26,27
This region contains populations of premotor neurons influencing sympathetic activation, including to the kidneys and the skin.
26,27
In context of this background, in the present study, lidocaine was bilaterally injected to block traffic, both neuronally generated and that carried by fibers of passage in the PVN during heat stress. This design was expected to elucidate the contribution of the PVN in processing thermal adjustments. Blockade of the PVN blunted the RSNA response to heat stress, consistent with decreased renal blood flow. Although the blood flow to other viscera, such as the gut, liver etc, are also important for blood redistribution to the skin during heat stress, the current findings demonstrate that one visceral bed, the kidney, is affected. Therefore, renal vasoconstriction during heat stress could be induced by the PVN directly through spinally projecting neurons and indirectly through the rostral ventrolateral medulla. The fact that injection of
The same pattern of response observed in RSNA was observed with MAP and HR responses after lidocaine and
In the case of rodents, tail skin vasodilation is the primary route of heat loss from the body, being responsible for the dissipation of an equivalent of 25% of resting heat production.
29,30
Heat stress-induced skin vasodilation is a consequence of tail sympathetic activity withdrawal, which is primarily activated by body heating stimulation over heat loss thermoregulatory centers.
31
Most of the thermoregulatory studies suggest that brain nitric oxide enhances heat dissipation by diminishing such sympathetic outflow to cutaneous vascular beds.
15–17
The present data demonstrated that
When analyzing the current results, we cannot disregard the possible interference of the anesthesia on the data, even though this method is widely used in the study of sympathetic activity and cardiovascular adjustments during heat stress. 2,34 It is possible that anesthesia may influence sympathetic nerve discharge responses and, consequently, thermoregulatory effectors during heat stress. Although this cannot be disregarded, the fact that heat stress results in increased sympathetic discharge in anesthetized rats suggest that anesthesia has only a slight affect, if any, on sympathetic activity during heat stress. 2,34 Additionally, it is recognized that anesthesia eliminates behavioral modifications that could also alter sympathetic nerve discharge during heat stress.
In summary, our results show that the PVN participates in increasing renal sympathetic activity in response to heat stress, contributing to cardiovascular adjustments that influence core blood flow redistribution from the visceral organs like the kidneys to the periphery. Furthermore, the PVN modulates heat loss through cutaneous vasodilation induced by body heating through nitric oxide mechanisms.
Footnotes
ACKNOWLEDGEMENTS
The technical assistance of Lirong Xu and Xuefei Liu is greatly appreciated. This work was supported by National Institutes of Health Grants HL6222 and DK082956, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG).