
Abstract
High-dose methotrexate (HDMTX) chemotherapy is generally accepted as an effective method for the treatment and prevention of extramedullary leukemia in children. However, it is unknown whether HDMTX chemotherapy kills intestinal bacteria on a large scale, thus causing dysbacteriosis, which may in turn influence the progress or prognosis of leukemia. The aim of this study was to examine changes in intestinal flora in children with acute lymphoblastic leukemia (ALL) treated with HDMTX chemotherapy. Bacterial DNA in stool from 36 healthy children and 36 ALL children were tested at A 260 with a spectrophotometer before and after HDMTX chemotherapy. The primers of Bifidobacteria, Lactobacillus and Escherichia coli were designed according to the 16SrRNA/DNA bacterial sequences. Bacteria were qualitatively and quantitatively confirmed by routine polymerase chain reaction (PCR) and fluorescent quantitative PCR, respectively. Our data showed that the total amount of flora in the stools of children with ALL was decreased by 29.6% compared with healthy children (P < 0.01). The total amount of flora in the stools of children with ALL on the third and seventh days after chemotherapy were 1496.5 ± 577.1 and 1966.6 ± 598.3 ng/μL, respectively, which was notably less than before chemotherapy (2436.3 ± 768.6 ng/μL). The amount of Bifidobacteria, Lactobacillus and E. coli in the intestinal tract in the ALL group after chemotherapy had an apparent change, which decreased most clearly on the third day, and partially recovered on the seventh day after chemotherapy. HDMTX chemotherapy can cause intestinal dysbacteriosis in children with ALL. The amount of Bifidobacteria, Lactobacillus and E. coli decreased significantly compared with the control group.
Introduction
Pediatric leukemia is a type of malignant tumor with the highest incidence in children. Tumor cells frequently metastasize to the central nervous system and the testes, resulting in extramedullary leukemia. Tumor cells that infiltrate to the central nervous system and testes can avoid being killed by drugs because most chemotherapeutic drugs do not easily pass the blood–brain and blood–testis barriers. This is the key reason that acute lymphoblastic leukemia (ALL) frequently re-occurs. Recently, with the continuous improvement in combined chemotherapy, the life of children with leukemia has been prolonged. High-dose methotrexate (HDMTX) chemotherapy is generally accepted as an effective method for the treatment and prevention of extramedullary leukemia. 1,2 Owing to the fast proliferation and rapid replacement of human intestinal mucosal cells and enteric cavity bacteria, and the abundant utilization of tetrahydrofolic acid (citrovorum factor [CF]) for DNA synthesis, bacterial flora is easily influenced by the pharmacological action of methotrexate (MTX). Numerous bacteria in the enteric cavity die when tumor cells are killed by MTX, and the intestinal flora lose the interface of colonization and survival because of intestinal mucosa injury, which easily leads to dysbacteriosis and influences the disease condition and prognosis of children with leukemia.
To understand the effects of HDMTX on intestinal bacterial flora in children with ALL, and to provide evidence for the rational use of microbial ecological agents for adjuvant therapy of leukemia, we quantitatively analyzed changes of intestinal Bifidobacteria, Lactobacillus and Escherichia coli in children with ALL before and after HDMTX chemotherapy by application of 16SrRNA/DNA fluorescent quantitative polymerase chain reaction (PCR).
Patients and methods
Patients
ALL group
Children with ALL who were treated with HDMTX were selected from the Department of Pediatrics of the First Hospital of Kunming Medical College between 2005 and 2006. The children were selected according to the following criteria: (1) clinical symptoms, physical signs and marrow cytological examination consistent with the diagnostic criteria for ALL; (2) the dose of MTX in the regimen was 3–5 g/m2, and assisted by hydration and alkalization; 3 (3) no use of antibiotics or microbial ecological agents for two weeks before HDMTX chemotherapy; (4) biochemical indices, such as hemograms and hepatorenal function, were almost normal before chemotherapy; and (5) routine examination of the stool before chemotherapy was normal. The stool samples of 36 children with ALL on the day before chemotherapy and on the third and seventh days after chemotherapy were collected in aseptic centrifuge tubes and stored at −70°C.
Healthy control group
Thirty-six healthy children were randomly selected from a kindergarten class according to the following criteria: (1) no diarrhea or other gastrointestinal diseases within four weeks; (2) no use of antibiotics and microbial ecological agents; and (3) routine examination of the stool was normal. The samples were collected and stored by the aforementioned method. The average age in the ALL group was 6.1 ± 1.1 y, with a male-to-female ratio of 1:1.15. The average age in the healthy control group was 5.8 ± 0.7 y, with a male-to-female ratio of 1:1.2. There were no statistical significances regarding age and gender between the two groups (t = 1.42, P > 0.05).
Methods
The methods are similar to previously described methods, 4–10 and modified as below.
Preparation of intestinal bacteria DNA
Fecal samples were taken from the −70°C freezer and put in the −20°C freezer for 24 h. They were then transferred to the 4°C refrigerator for another 24 h, and then placed at room temperature. Stool (0.2 g) was weighed and fully mixed with 1.5 mL of phosphate buffer solution (PBS; 0.05 mol/L [pH 7.4]) for five minutes and centrifuged at 1000 rpm for 10 min. The supernatant was collected, and this sequence was repeated three times according to the above method. The entire supernatant was collected and centrifuged at 14,000 rpm for 10 min. The supernatant was then discarded, and the sediment was collected and mixed with 1 mL of PBS, followed by centrifugation at 14,000 rpm for five minutes. Subsequently, the supernatant was discarded and the sediment was washed repeatedly with PBS, and then washed once with water. Bacteria were suspended in 100 μL of distilled water and lysed with 100 μL of 1% Triton X-100. Sample DNA from the control and ALL groups was detected with a spectrophotometer at A 260.
Purification of DNA
Bacterial liquid was centrifuged at 14,000 rpm; 200 μL of supernatant was removed and agitated with isovolumetric phenol/chloroform for 10 s, and then centrifuged at 14,000 rpm for 10 s. Subsequently, the supernatant was collected and fully mixed with a 2–2.5-fold volume of 100% iced alcohol and then placed on ice for 20 min. After centrifugation at 14,000 rpm, the supernatant was discarded and the pellet was precipitated once with 70% alcohol. The centrifuge tubes were heated and dried at 100°C, and then 50 μL of TE buffer were added to the tubes at room temperature.
Routine PCR
Bacterial primers were designed with primer-design software and aligned and verified with the BLAST database ( Bands 1 to 4 are fragments of Escherichia coli (95 bp) and bands 5 to 8 are fragments of Bifidobacteria (203 bp) Band 1 is the marker, bands 2 to 5 are blank, bands 6 and 7 are fragments of Escherichia coli (95 bp) and bands 8 and 9 are fragments of Lactobacillus (166 bp)

Fluorescent quantitative PCR
The reaction included 0.5 μL of forward primer (25 μmmol/L), 0.5 μL of reverse primer (25 μmmol/L), 5 μL of DNA template, 25 μL of SYBR Green (including 10× buffer, 4× dNTP, MgCl2 and Taq polymerase) and 19 μL of distilled water, for a total of 50 μL. Sequences were amplified by a GeneAmp PCR system 5700 fluorescent quantitative PCR equipment (Applied Biosystems, Inc, Foster City, CA, USA), and a melting curve was made. The reaction conditions were set as follows: pre-denaturation for five minutes at 95°C, denaturation for 15 s at 95°C, annealing for one minute at 60°C and extension for 45°C at 72°C. A total of 40 cycles were performed and then ended with a final extension for 10 min at 72°C. The temperature for removing the primer–dimers was set with the melting curve and added to the cycle program; the fluorescent quantitative amplification was made under the following conditions. After five minutes of pre-denaturation, the conditions were set as follows: Bifidobacteria: 95°C for 15 s, 60°C for one minute, 72°C for 45 s and 88°C for five seconds; E. coli: 95°C for 15 s, 60°C for one minute, 72°C for 45 s and 82°C for five seconds. A total of 40 cycles were performed and ended with a final extension for 10 min at 72°C. The initial cycle number was obtained from the fluorescent curve when the fluorescent sign appeared.
Drawing standard curve and quantitative analysis
Bifidobacteria and Lactobacillus freeze-dried powder (batch number 11054; The National Strain Management Center of China, Beijing, PR China). E. coli (provided by the laboratory department of our hospital) were inoculated in liquid MRS media and cultured in anaerobic environment for 24 h at 37°C (90% N2 + I 0% CO2). One milliliter of medium was removed and centrifuged at 3000 rpm for 15 min, the supernatant was discarded and the sediment was obtained. Fifty microliters of DNA solution dissolved in TE was obtained according to the aforementioned isolation and DNA purification methods. DNA absorbance at 260 nm was detected with a spectrophotometer. The concentration of DNA templates of standard strains was calculated according to the formula, A 260 value = 4.74 × 1013 molecules. After a 10-fold serial dilution, a concentration gradient from 107 to 104 was obtained. DNA templates with different concentrations were amplified by fluorescent quantitative PCR. The initial cycle number (Ct ) in the PCR procedure was designated as the ordinate when the fluorescent sign appeared, the logarithm of the copy number of different DNA templates was designated as the abscissa, the standard curve was drawn, and then the intercept (b), slope (k) and correlation coefficient were obtained. Finally, the initial cycle number of each sample was put into the regression equation, log Co = (Cn − b)/k and the corresponding DNA template number was obtained.
Statistical analysis
After logarithmic transformation, the results were displayed in the form of X lgx ± S lgx. The bacterial numbers from the ALL and control groups were compared by SPSS, version 13.0 (SPSS Inc, Chicago, IL, USA), and the results from the ALL and control groups were analyzed by one-way analysis of variance (ANOVA). A paired t-test was used in the ALL group for comparison before and after chemotherapy. P < 0.05 indicated a significant difference.
Results
Amount of flora in the stool was notably less in children with ALL than in healthy children
After isolation and purification of bacterial DNA from the stool, the spectrophotometer reading at A
260 indicated the total amount of bacteria in the stool. The results showed that the amount of bacteria in the healthy control group was 3479.3 ± 870.5 ng/μL and the amount of bacteria in the ALL group on the day before chemotherapy was 2436.3 ± 768.6 ng/μL (Figure 3). The homogeneity test of variances between the two groups suggested that the variance was homogeneous (P 0.05). The one-way ANOVA showed that the difference between the two groups was significant (F = 50.78, P 0.01). The total amount of flora in the stools of children with ALL decreased by 29.6% compared with healthy children (P 0.01).
Comparison of bacterial DNA-A
260 between the control and the acute lymphoblastic leukemia (ALL) groups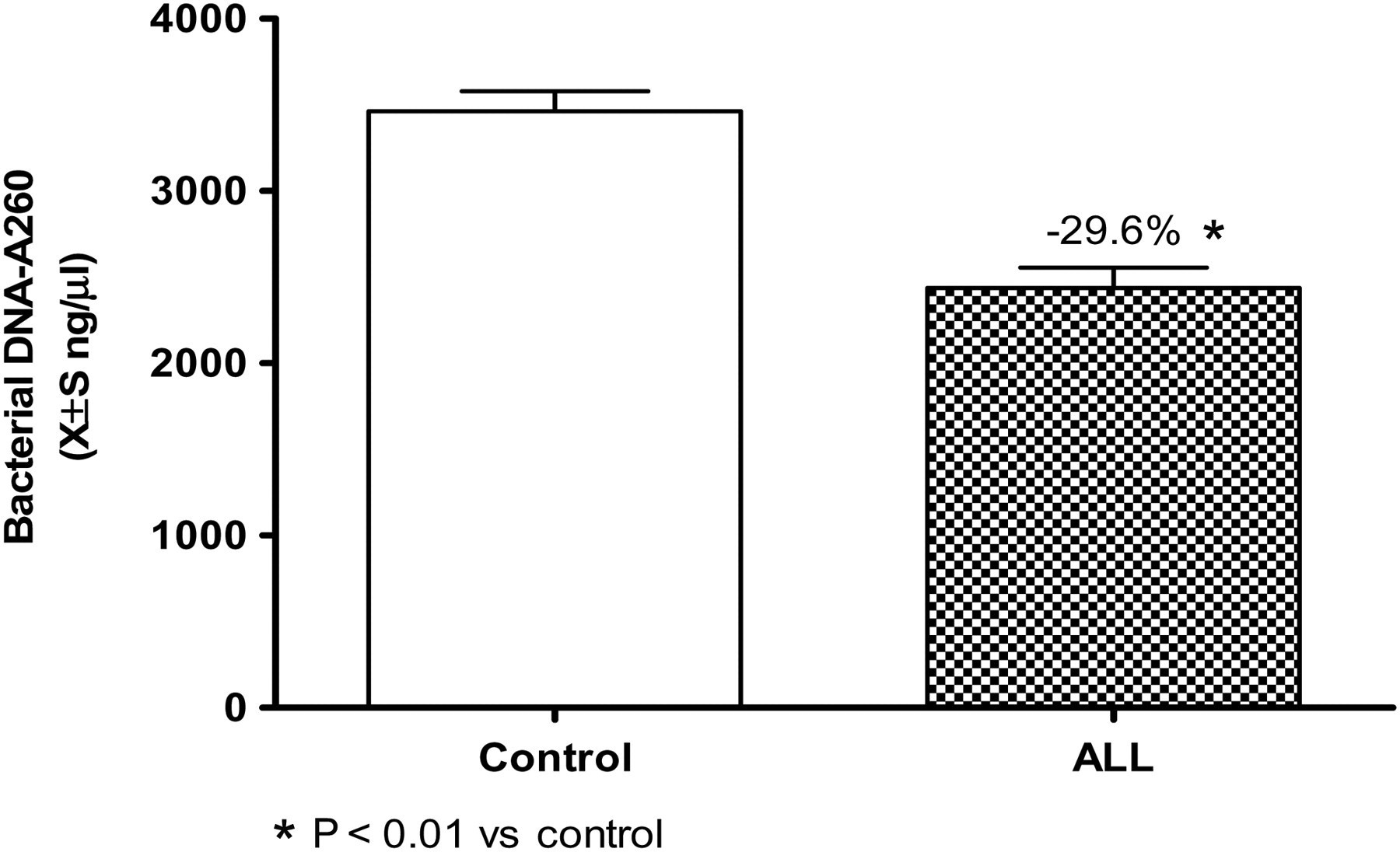
Bifidobacteria, Lactobacillus and E. coli in the stool were analyzed. In the control group, Bifidobacteria had 9.49 ± 0.41 copies/g moist stool, Lactobacillus had 9.12 ± 0.50 copies/g moist stool and E. coli had 7.52 ± 0.43 copies/g moist stool. However, in the ALL group, Bifidobacteria had 7.24 ± 0.33 copies/g moist stool, Lactobacillus had 8.10 ± 0.43 copies/g moist stool and E. coli had 6.62 ± 0.42 copies/g moist stool. The difference in bacterial amounts between the two groups was statistically significant (F = 739.61, 178.19 and 88.67, respectively; P 0.01). The amount in the control group was larger than in the ALL group (Figure 4). Therefore, either the total amount of bacteria or the amount of Bifidobacteria, Lactobacillus and E. coli in the stool of children with ALL was significantly less than in healthy children.
Comparison of the amount of Bifidobacteria, Lactobacillus and Escherichia coli in stool between the control and acute lymphoblastic leukemia (ALL) groups before chemotherapy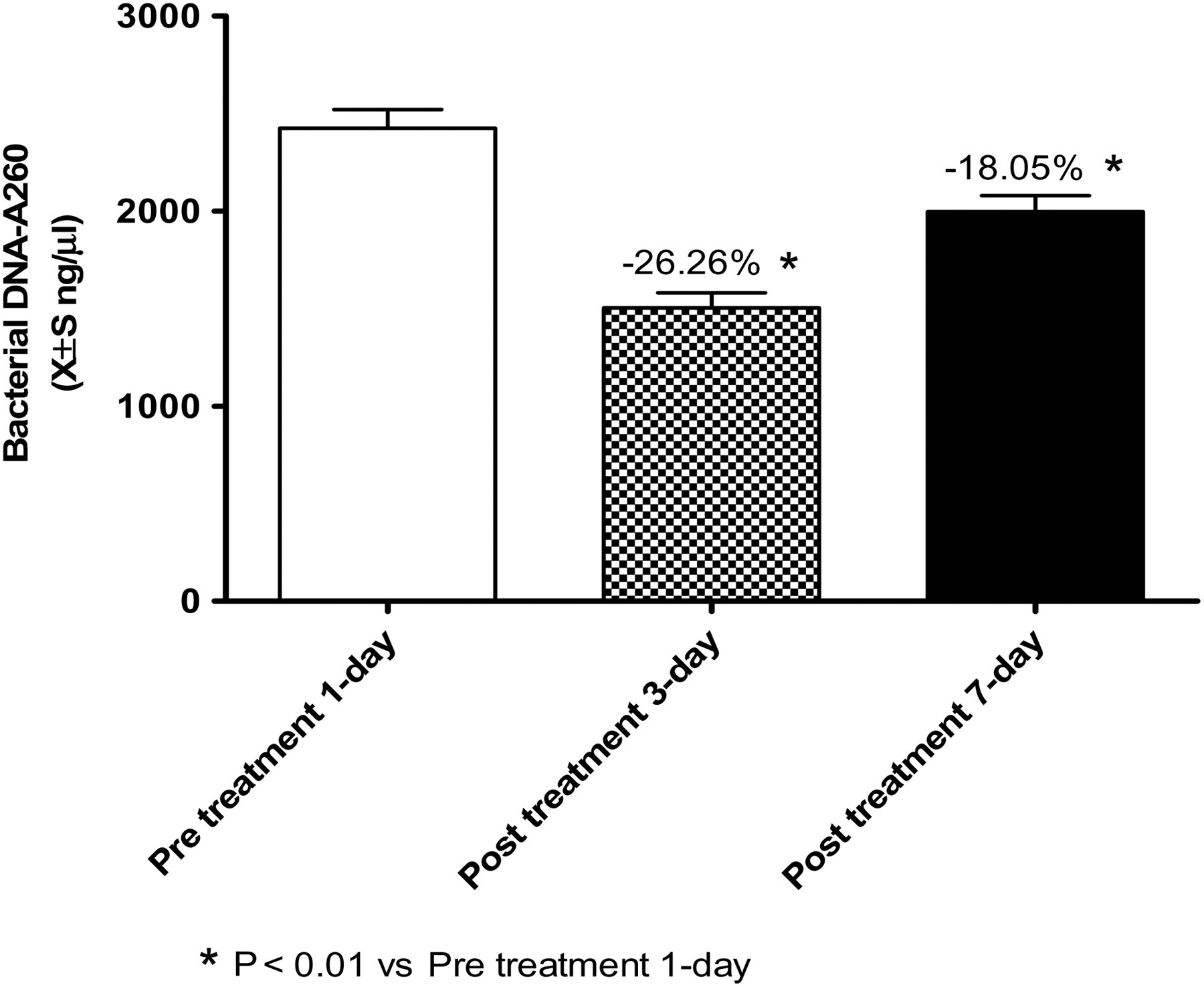
Amount of bacteria in the stool after chemotherapy was notably less than before chemotherapy
The amount of bacteria in the stools of children with ALL was determined the day before chemotherapy, and on the third and seventh days after chemotherapy. The results showed that the amount of bacteria in the stool on the day before chemotherapy was 2436.3 ± 768.6 ng/μL and the amount of bacteria on the third day after chemotherapy dramatically decreased to 1496.5 ± 577.1 ng/μL, a reduction of 26.26%. The amount on the seventh day after chemotherapy partially recovered to 1966.6 ± 598.3 ng/μL, decreasing by 18.05% compared with the amount on the day before chemotherapy. The amounts of bacteria on the third and seventh days after chemotherapy decreased markedly compared with the amount of bacteria the day before chemotherapy, and there were statistically significant differences (Figure 5).
Comparison of bacterial DNA-A
260 before and after chemotherapy between the control and acute lymphoblastic leukemia (ALL) groups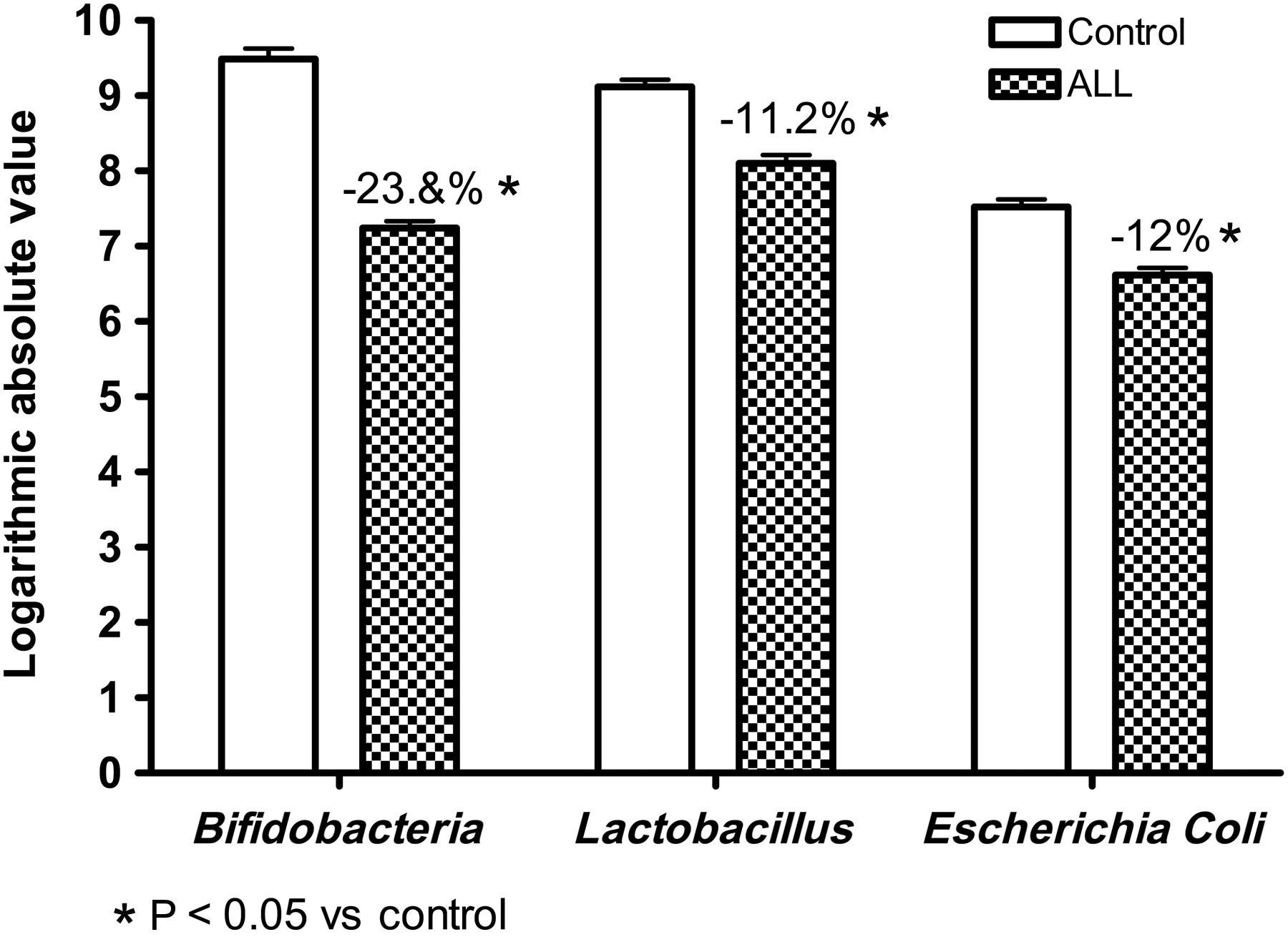
Amount of Bifidobacteria, Lactobacillus and E. coli in stool after chemotherapy had an apparent change compared with before chemotherapy in children with ALL
The amounts of Bifidobacteria, Lactobacillus and E. coli in stool from children with ALL were determined the day before chemotherapy and the third and seventh days after chemotherapy (Figure 6). The results showed that the amount of Bifidobacteria in the stool the day before therapy was 7.24 ± 0.33 copies/g moist stool, and the amount on the third and seventh days after therapy decreased by 17.13% (6.00 ± 0.27 copies/g moist stool) and 8.98% (6.59 ± 0.33 copies/g moist stool), respectively. The latter two amounts significantly decreased compared with the amount the day before chemotherapy, a statistically significant difference existed. Similarly, the amount of Lactobacillus decreased significantly compared with the amount before chemotherapy. Specifically, the amount of Lactobacillus in the stool the day before chemotherapy was 8.10 ± 0.43 copies/g moist stool, and on the third and seventh days after chemotherapy decreased by 16.91% (6.73 ± 0.45 copies/g moist stool) and 8.02% (7.45 ± 0.43 copies/g moist stool), respectively. Interestingly, the amount of E. coli in stool the day before chemotherapy was 6.62 ± 0.42 copies/g moist stool, and the amount on the third day after chemotherapy decreased by 10.88% (5.96 ± 0.42 copies/g moist stool), with a significant difference in the amount the day before chemotherapy. However, the amount on the seventh day recovered to 7.02 ± 0.41 copies/g moist stool, and increased by 6.04%, which was a significant difference compared with the amount the day before chemotherapy.
Comparison of the amount of Bifidobacteria, Lactobacillus and Escherichia coli in the stool of the acute lymphoblastic leukemia (ALL) group before and after chemotherapy. The amount of Bifidobacteria on the third and seventh days after therapy decreased by 17.13% and 8.98%, respectively. The amount of Lactobacillus on the third and seventh days after chemotherapy decreased by 16.91% and by 8.02%, respectively. The amount of Escherichia coli on the third day after chemotherapy decreased by 10.88% and increased by 6.04% on the seventh day after chemotherapy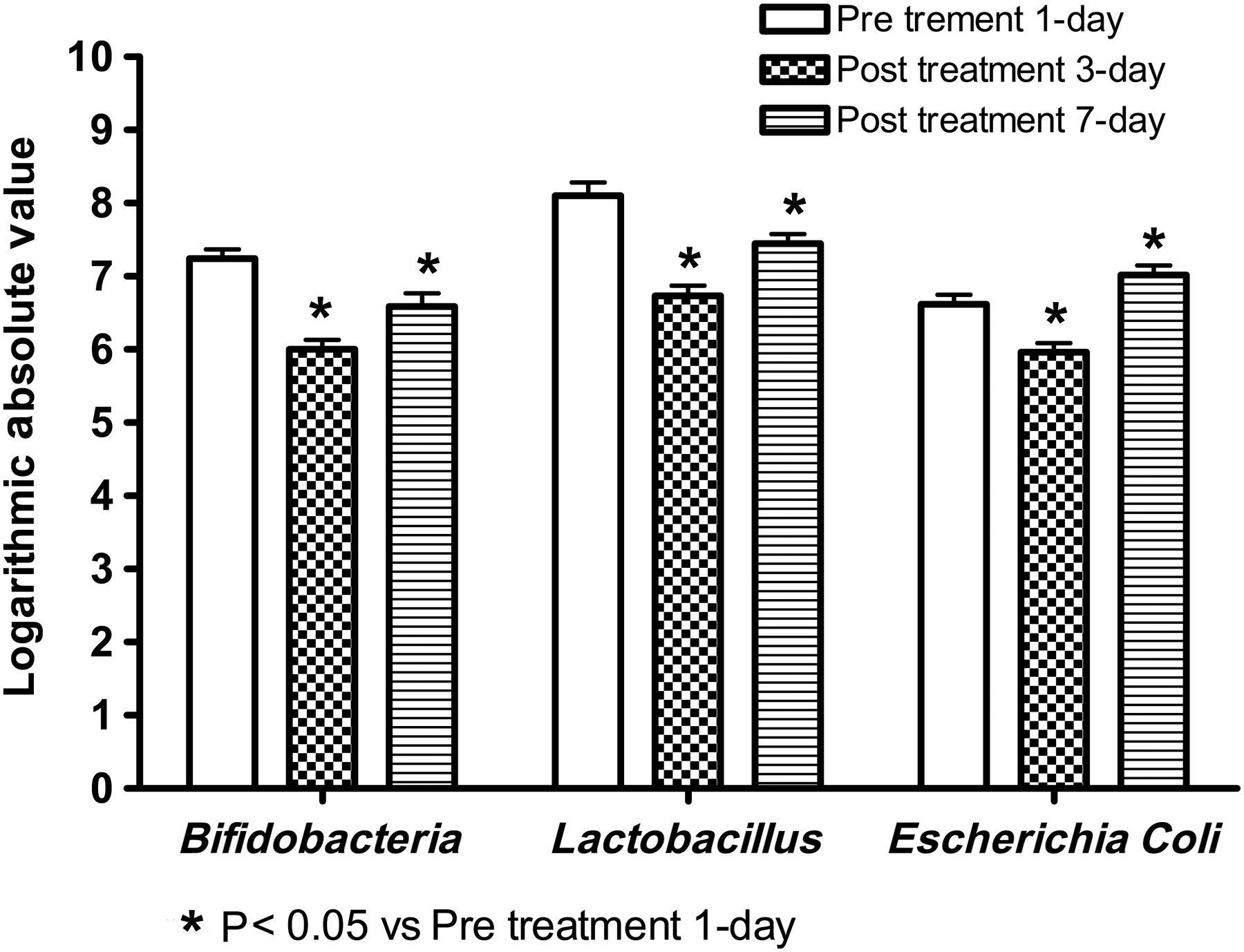
Discussion
The human gut is the natural habitat for a large and dynamic bacterial community, but recently developed molecular biology tools discovered significant inter-subject variability and differences between the composition of stool and mucosal communities. Characterization of this immensely diverse ecosystem is the first step in elucidating its role in health and disease. The human endogenous intestinal microflora is an essential ‘organ’ in providing nourishment, regulating epithelial development, preventing mucosal barrier dysfunction, and instructing innate immunity. Intestinal microflora might also be an essential factor in certain pathological conditions, including multisystem organ failure, colon cancer and inflammatory bowel diseases. Nevertheless, bacteria are also useful in promoting human health. Probiotics and prebiotics are known to have roles in the prevention or treatment of some diseases. 11–13
The intestinal microflora is the largest and most important microecosystem in the body. Normal flora is formed with long-term evolution of the host. Owing to the poor equilibrium stability, the intestinal microflora is influenced easily by various factors, such as feeding patterns, age, living habits, hygienic environment, radiotherapy, chemotherapy and antibiotics, which could result in dysbacteriosis, 14 increasing intestinal permeability, 15–20 intestinal barrier dysfunction 21–23 and intestinal bacteria and/or endotoxin (including bacterial cell wall components, such as lipopolysaccharide and peptidoglycan polysaccharide translocation). 24–28 It is possible that enteric bacteria, their cell wall components, or both, may disseminate throughout the body, causing sepsis, shock, multisystem organ dysfunction or death of the host. 27 These disturbances in critically ill patients may influence their outcome and prognosis.
A better understanding of the factors that influence translocation could result in the implementation of interventions that contribute to improved patient outcomes. Glutamine supplementation and targeted nutritional intervention (including Bifidobacteria, maintaining splanchnic flow, the judicious use of antibiotics and directed selective gut decontamination regimens), hold some promise in limiting bacterial translocation. 15,20,22,23,27 It is hoped that improving our understanding of the role of the gut in disease processes will lead to the development of novel therapeutic strategies in the future.
To determine whether dysbacteriosis or the graduation of dysbacteriosis exists in clinical practice, concrete quantitative analysis on the intestinal flora is needed. In this study, Bifidobacteria, Lactobacillus and E. coli in stool among a control group and children with ALL were quantified before and after HDMTX chemotherapy by 16SrRNA/DNA fluorescent quantitative PCR. The results showed that the amount of Bifidobacteria, Lactobacillus and E. coli in the stools of the ALL group were much less than in the control group. After chemotherapy, the amount of Bifidobacteria, Lactobacillus and E. coli in the stools from the ALL group decreased significantly compared with the amount before chemotherapy, especially on the third day after chemotherapy; however, there was partial recovery by the seventh day after chemotherapy. The amount of E. coli was greater after chemotherapy than before chemotherapy, indicating that HDMTX chemotherapy could lead to a further disorder of intestinal flora in children with ALL. This reduction and disorder gradually improved with drug metabolism. In general, it takes two weeks or more to completely recover from this disorder.
MTX was toxic to normal cells that rapidly proliferate and differentiate, such as intestinal epithelial cells and bone marrow cells. Sufficient hydration, alkalization of urine and rational CF rescue was used to ensure the safety of HDMTX application (12 g/m2), but animal experiments and long-term clinical observation showed that HDMTX still caused notable intestinal mucosal injury and toxic side-effects, such as diarrhea and hematochezia. 29 The intestinal flora lost the interface of colonization because of intestinal mucosal injury, and numerous bacteria that acted as cell bodies were probably killed, leading to a reduction in probiotics and dysbacteriosis. It was discovered in mice fed in a bacteria-free environment that the growth rate of pit cells, which differentiate and produce intestinal epithelial cells, decreased and the crypt cell number was significantly less than in mice with normal flora in the colon. 30 Experimental data have also shown that Bifidobacteria and Lactobacillus, as well as their metabolites (mainly short-chain fatty acids), are able to not only promote the synthesis of intestinal epithelial cells (IEC-6) DNA to quicken the proliferation of intestinal epithelial cells, 31 but can also prevent intestinal epithelial cells from generating tumor necrosis factor (TNF), which can relieve inflammatory injury and protect the mucosal barrier. 32 This suggests that loss of nutrition, protection and repair of intestinal normal flora of intestinal mucosa was probably the reason that hematochezia, constipation and other symptoms occurred and developed.
There is a close relationship between the occurrence, development and prognosis of ALL, and the immune status of patients. Lymphocyte carcinogenesis and immune inhibition of chemical drugs have always been the main reasons underlying decreased immune function and secondary infection in children with ALL. The reduction of intestinal probiotics and dysbacteriosis during chemotherapy affects immune function directly or indirectly. A study showed that lymphocytes in associated lymphoid tissues of germ-free animals were decreased, 33 whereas after bacterial inoculation in the intestinal tract, lymphocytes in the mesentery increased, the speed and strength of lymphocytes against antigen increased, the clonal diversity of peripheral hyperglobulinemia increased and the function of phagocytes was enhanced, suggesting that bacterial inoculation of germ-free cells in the intestinal tract enhances the local and systemic immune functions of the intestinal tract. Liang et al. 34 discovered in mouse experiments that dysbacteriosis leads to a reduction of humoral and cellular immunity and non-specific immunity, as well as hematopoietic function. Chemotherapy can damage the intestinal flora of leukemia patients, and dysbacteriosis can lead to a reduction in immunity function. Chen and Wen 35 analyzed the intestinal flora of leukemia patients before chemotherapy and found that the amount of Bifidobacteria and E. coli decreased significantly compared with healthy children. The degree of reduction was closely related to the incidence of infection and septicemia after chemotherapy. Therefore, the effect of intestinal flora on immunity in chemotherapy should not be negligible. As proposed by Delaloye et al., 36 protection of body immunity and the killing of tumor cells are equally important in the process of tumor therapy. It is known that Bifidobacteria promote the secretion of cytokines, such as interleukin 26 and TNF2α, by activating macrophages, which play an indirect role in tumor inhibition. 37 In addition, peptidoglycan, teichoic acids and polysaccharides in the cell walls of Bacillus bifidus also have antitumor effects. A study by Tan and Yao 38 showed that after tumor therapy with Bifidobiogen–Livzon, tumors were significantly smaller than in the control group, which is evidence of the view mentioned above. Data from our experiments showed that a notable reduction in Bifidobacteria led to declines in antitumor ability. Therefore, the active prevention of dysbacteriosis after chemotherapy in children with ALL will not only enhance body immunity and reduce the incidence of complications but may also profoundly improve long-term prognoses.
E. coli, the main physiological symbiotic bacteria, forms an intestinal biological barrier with other anaerobic bacteria, which is resistant to the colonization and invasion of pathogenic bacteria, and plays a role in inhibiting the growth of opportunistic pathogens. 39 In addition, it has a stimulatory function that is similar to antigens, and mediates specific and non-specific immunity. 40 This study showed that chemotherapy also resulted in a significant reduction of E. coli. However, on the seventh day after chemotherapy, E. coli levels recovered greatly and exceeded that after chemotherapy. We postulate that MTX, especially at high doses, can damage the gastrointestinal structure, which may then affect anaerobic bacteria including Bifidobacteria colonization, because Bifidobacteria must have tight conjunctions with the intestinal epithelial cell to survive and form the biological barrier in physiological conditions. However, E. coli do not depend on intestinal epithelial cells and can grow in the intestinal tract. The quicker recovery of E. coli colonies compared with Bifidobacteria after influence by external factors, may be because E. coli is a kind of facultative anaerobe and can grow in aerobic and anaerobic environments. Owing to the great reduction of other aerobic and anaerobic bacteria during chemotherapy, the peripheral biological antagonism obviously weakens, and thus provides a favorable microhabitat for the propagation of E. coli. The quick proliferation of E. coli might overgrow and easily result in strain imbalance, and then cause dysbacteriosis or dual infection, as well as septicemia.
Our study also showed that there was a significant difference of intestinal flora between the ALL group and the healthy control group before therapy, which was likely caused by children with ALL receiving other therapies before admission. During the treatment process, various kinds of chemotherapeutics that destroy DNA and RNA, or inhibit the synthesis of DNA, RNA and proteins, were used in the children. In addition, repeated application of antibiotics for concurrent infections destroyed the ecological balance of the intestinal flora. The relationship between the prevalence, improvement rate and prognosis of children with leukemia deserves further study and investigation. Specifically, our data indicate that due attention should be paid to intestinal microecology and the microenvironment with rational prevention against dysbacteriosis and proper use of antibiotics to regulate the intestinal colony balance.
Footnotes
ACKNOWLEDGEMENT
No financial assistance was received in support of this study.