
Abstract
Hypertrophy and impaired mineralization are two processes closely associated with osteoarthritis (OA). 1,25-dihydroxyvitamin D3 (1a,25(OH)2D3) and inorganic phosphate (Pi) are two important factors that are implicated in calcium and phosphate homeostasis of bone metabolism and both can be regulated by the circulating phosphaturic factor fibroblast growth factor 23 (FGF23). The objective of this study was to investigate the role of 1a,25(OH)2D3 and Pi and the molecular mechanism through which they contribute to hypertrophy and mineralization in human osteoarthritic chondrocytes. For this purpose, primary human chondrocytes were obtained from articular cartilage which was collected after total knee replacement surgery in OA patients. FGF23, fibroblast growth factor receptor 1c (FGFR1c), vitamin D3 receptor (VDR), and phosphate inorganic transporter-1 and -2 (PiT-1 and PiT-2) expression levels were evaluated and found to be significantly higher in OA chondrocytes compared with normal. In addition, we observed that the binding of FGF23 to FGFR1c was stronger in OA chondrocytes compared with normal. Chromatin immunoprecipitation (ChIP) assay revealed, for the first time, the presence of two vitamin D response elements (VDREs) in the FGF23 promoter. Treatment of normal chondrocytes with 1a,25(OH)2D3 or Pi resulted in significant up-regulation of VDR, FGF23, PiT-1, PiT-2 mRNA and protein levels, extracellular signal-regulated kinases 1/2 (ERK1/2) phosphorylation and induction of hypertrophy markers collagen type X (COL10A1), osteopontin (OPN), osteocalcin (OC), catabolic markers metalloproteinase-13 (MMP-13) and the apoptotic marker caspase-9. Furthermore, VDR silencing in OA chondrocytes negatively regulated FGF23, COL10A1, OPN, OC, MMP-13 and caspase-9 expressions and ERK1/2 phosphorylation. Finally, combined VDR silencing and PiT-1, PiT-2 inhibition in OA chondrocytes resulted in additive down-regulation of FGF23 expression, ERK1/2 activation and COL10A1, OPN, OC, MMP-13 and caspase-9 expression levels. We propose that 1a,25(OH)2D3 and Pi act synergistically through FGF23 signaling and ERK1/2 phosphorylation contributing to late hypertrophic events and impaired mineralization in osteoarthritic chondrocytes.
Introduction
Development and progression of osteoarthritis (OA) has been associated with altered mineral-ion homeostasis and chondrocyte hypertrophy. 1 During osteoarthritic cartilage development, chondrocytes resume the characteristics of hypertrophic chondrocytes in the growth plate, including fibrillation of cartilage surface, decreased proteoglycan content of the matrix, increased expression of type X collagen (COL10A1) and extracellular matrix (ECM) mineralization through the release of matrix vesicles able to accumulate calcium and phosphate. 2–5
Vitamin D3 and its active metabolites like 1,25-dihydroxyvitamin D3 (1a,25(OH)2D3) (1,25D) play an essential role in the mineralization process in the growth plate, inhibiting DNA synthesis of hypertrophic chondrocytes. 6 Moreover, 1a,25(OH)2D3 acts directly on matrix vesicles causing loss of their membrane integrity and release of metalloproteinases (MMPs) in the ECM, suggesting its role in modulating the degree of matrix degradation during chondrocyte hypertrophy. 7–10 In addition, 1a,25(OH)2D3 exerts other biological functions, such as regulation of cell proliferation, differentiation, immune function and apoptosis by binding to vitamin D receptors (VDRs), 11,12 which are expressed in different cell types, including chondrocytes, modulating thus their behavior within the different zones of the growth plate. 8,11
Among ECM components, extracellular inorganic phosphate (ePi) concentrations are critical determinants of mineralization and depend on the extracellular hydrolysis of pyrophosphate by nucleotide triphosphates and on sodium-dependent phosphate (Na/Pi type III) cotransporters, the phosphate inorganic transporter-1 and -2 (PiT-1 and PiT-2), which allow ePi to enter into the cells. 5,13–15 In situ hybridization analysis has revealed PiT-1 expression in a subpopulation of hypertrophic chondrocytes during endochondral bone formation, suggesting a potential role for this Pi carrier in primary events of extracellular matrix calcification. 16 Enhanced expression of PiT-1 has been found in chondrocytes throughout OA cartilage including hypertrophic chondrocytes and the implication of Pi cotransporters has been suggested in pathological calcification. 17 In addition, it has been reported that increased extracellular Pi triggers signal transduction and results in altered gene expression in some cell types, as osteopontin (OPN) in the mouse osteoblastic cell line MC3T3-E1. 18,19 Pi homeostasis has been also shown to be affected by 1a,25(OH)2D3 participating in a negative feedback loop with fibroblast growth factor 23 (FGF23) in mineralized tissue cells. 20–24
FGF23 is a circulating phosphaturic factor that plays a central role in renal reabsorption of Pi and vitamin D3 metabolism. 25 Premature-aging-like phenotype in fgf23–/– mice has been correlated with elevated levels of active vitamin D3. 26 FGFs exert their biological actions by binding to, dimerizing and activating FGF receptor (FGFR) tyrosine kinases. Among FGFRs, FGFR1 is suggested to be the biologically relevant receptor for FGF23. 27,28 It has been shown that FGF signalling induces extracellular signal-regulated kinases 1 and 2 (ERK1/2) activation resulting in the expression of target genes in order to inhibit phosphate transport in the proximal tubules. 29
In the present study, we investigated the role of extracellular Pi and 1a,25(OH)2D3, as well as the molecular pathway through which extracellular Pi and 1a,25(OH)2D3 contribute to hypertrophy and mineralization in osteoarthritic chondrocytes.
Materials and methods
Patients and articular cartilage samples
Articular cartilage samples were obtained from femoral condyles and tibial plateaus of patients with end-stage primary OA undergoing knee replacement surgery. A total of 40 patients were included in the study (30F/10M; mean age 64.18 ± 14.24 years). Normal cartilage was obtained from 10 individuals (7F/3M; mean age 44.60 ± 7.6), undergoing fracture repair surgery. Radiographs were obtained before surgery and graded according to the Kellgren–Lawrence (K/L) and Mankin system. All osteoarthritic cartilage samples had K/L score >3 and Mankin score 10–14, while normal cartilage had 0 Mankin score. Moreover, all osteoarthritic samples were characterized by the presence of osteophytes. Patients with rheumatoid arthritis and other autoimmune disease as well as chondrodysplasias, infection-induced OA and post-traumatic OA were excluded from the study. Both patients and healthy individuals' cartilage samples were obtained upon the individuals’ verbal informed consent. The method of obtaining verbal approval by all individuals was approved by Institutional Review Board of the University Hospital of Larissa. The protocol was also approved by the local ethics committee of University Hospital of Larissa. The study protocol conformed to the ethical guidelines of the 1975 Declaration of Helsinki as reflected in a priori approval by the Local Ethical Committee of the University Hospital of Larissa.
Primary cultures of human articular chondrocytes, normal and osteoarthritic
Articular cartilage was dissected and subjected to sequential digestion with 1 mg/mL pronase (Roche Applied Science, Mannheim, Germany) and 1 mg/mL collagenase P (Roche). Chondrocytes were counted and checked for viability using trypan blue staining. More than 95% of the cells were viable after isolation. Chondrocytes were then seeded in six-well plates with Dulbecco's modified Eagle's medium/Ham's F-12 (DMEM/F-12; Gibco, BRL, Paisley, UK) supplemented with 5% fetal bovine serum (FBS; Gibco) and 100 U/mL penicillin–streptomycin (Gibco) and incubated at 37°C under a humidified 5% CO2 atmosphere until reaching confluence.
RNA extraction from articular cartilage and cultured chondrocytes and realtime polymerase chain reaction
Total cellular RNA was extracted from fresh articular cartilage as well as from cultured chondrocytes using Trizol reagent (Invitrogen, Life Technologies, Paisley, UK). RNA was further purified using an RNeasy mini kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Preservation of 28S and 18S rRNA species was used to assess RNA integrity. All samples included in the study were with prominent 28S and 18S rRNA components. The yield was quantified spectrophotometrically.
Oligonucleotide primers used in realtime PCR assay
COL10A1, collagen type X alpha 1; MMP-13, metalloproteinase-13; OPN, osteopontin; OC, osteocalcin; FGF23, fibroblast growth factor 23; FGFR1c, fibroblast growth factor receptor 1c; Pit-1, sodium-dependent phosphate transporter 1; Pit-2, sodium-dependent phosphate transporter 2; VDR, vitamin D receptor; GAPDH, glyceraldehyde-3-phosphate dehydrogenase
Protein expression evaluation by Western blot analysis
Osteoarthritic and normal articular chondrocytes were lysed using Nonidet P-40 lysis buffer containing 30 mmol/L Tris (pH 7.5; Sigma-Aldrich, St Louis, MO, USA), 150 mmol/L NaCl (Sigma-Aldrich), 10% glycerol (Sigma-Aldrich), 1% Nonidet P-40 (Sigma-Aldrich) and a cocktail of protease inhibitors (Roche). Protein concentration was quantified using the Bio-Rad Bradford protein assay (BioRad, Hercules, CA, USA) with bovine serum albumen as standard.
Cell lysates were electrophoresed and separated on a 4–10% Tris-HCl gel (Bio-Rad) and transferred to a Hybond-ECL nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ, USA), which was probed with antibodies against FGF23 (1:1000 dilution; Novus Biologicals Inc, Littleton, CO, USA), FGFR1c, total ERK1/2, phosphor-ERK1/2, COL10A1, OPN, OC, PiT-1 and PiT-2 (1:1000 dilution; Abcam, Cambridge, UK), VDR (1:1000 dilution; R&D Systems, McKinley Place, NE, Mineapolis, USA), caspase-9 (1:1000 dilution; Santa Cruz Biotechnology Inc, Heidelberg, Germany), and signals were detected using secondary antibody IgG conjugated with horseradish peroxidase (1:5000 dilution; Santa Cruz Biotechnology). The results were normalized using goat polyclonal beta-actin antibody (Santa Cruz Biotechnology).
FGF23 measurement in cell culture media
In order to determine the secreted levels of FGF23 from OA and normal samples, culture media from primary OA and normal chondrocyte cultures were analyzed for FGF23, using specific enzyme-linked immunosorbent assay (Immunotopics, Inc, San Clemente, CA, USA) according to the manufacturer's instructions. FGF23 concentrations were determined in duplicate. According to the manufacturers, the detection limit for FGF23 assay was ≤3 U/mL. The intra- and inter-assay coefficient of variation were 5% and 7.3%, respectively.
FGF23–FGFR1c immunoprecipitation assay
Sub-confluent cells of normal and OA chondrocytes cultures were lysed in 20 mmol/L Tris-HCl buffer (pH 8) (Sigma-Aldrich) containing protease inhibitors (Roche). In separate aliquots of cell lysates, antibody against FGFR1c (Abcam) or FGF23 (Novus Biologicals Inc) was added. Protein A/G agarose beads (Thermo Scientific, Rockford, IL, USA) were also added in every sample and they were incubated overnight at 4°C. Agarose beads were collected and washed with lysis buffer (pH 8). Based-bound proteins were resolved on 4–10% sodium dodecyl sulfate-polyacrylamide gels (Invitrogen), and the gels were blotted on nitrocellulose membranes (Amersham Biosciences). The membranes were incubated with 5% blocker and then with anti-FGF23 of FGFR1c antibody, respectively. The membranes were then exposed to photographic film (Kodak, Rochester, NY, USA). IgG from the same species as the antibody being used for the immunoprecipitation was used as a negative control.
Treatment of normal or OA chondrocytes
Three days post-seeding, normal chondrocytes were treated with varying concentrations (10−7, 10−6 mol/L) of 1a,25(OH)2D3 (Sigma-Aldrich) or DMEM/F-12 alone for 24, 48 and 72 h, at which time cells were harvested. In each experiment, the results from three out of the six-well plates were averaged and considered as n = 1. No significant variance was observed among the individual wells in each averaged group.
We also treated normal chondrocytes with varying concentrations of phosphate or phosphate together with phosphonoformic acid (PFA; Sigma-Aldrich), a Pi transporter inhibitor. In order to prepare the treatment media enriched with Pi, Pi (NaH2PO4; Sigma-Aldrich) was dissolved in warm DMEM/F12 (37°C). When phosphate salts were completely dissolved, pH was adjusted to 7.4, and the solution was filter sterilized. The actual concentration of Pi in the serum-free medium was 1 mmol/L and this value was set as control. The concentrations of Pi which were tested were 3, 5 and 7 mmol/L at 48 h. PFA was used at a concentration of 1 mmol/L in the presence of 1 (control) to 7 mmol/L Pi.
Vitamin D response element promoter analysis
Oligonucleotide primers used for amplification of DNA fragments isolated from ChiP assay
FGF23, fibroblast growth factor 23; OC, osteocalcin
Oligonucleotide transfections
Osteoarthritic chondrocytes were seeded in six-well plates with DMEM/F-12 containing 5% FBS. After overnight incubation, cells were transfected with specific siRNA against VDR (Invitrogen), scrambled siRNA (Invitrogen) and control which contained only OPTI-MEM, using Lipofectamine™ 2000 (Invitrogen) as described by the manufacturer. No cell toxicity was detected due to the transfection agent.
In five osteoarthritic cultures, after VDR silencing, the medium was changed to DMEM/F-12 containing 1 mmol/L PFA.
RNA and cell lysates were extracted 48 h after si-VDR transfection or si-VDR together with PFA. Expression levels of mRNA and protein of VDR, FGF23, FGFR1c, OPN, OC, COL10A1 and MMP-13 were determined by realtime PCR and Western blot, as described above. In addition, protein levels of caspase-9, phosphor- and total ERK1/2 were also determined by Western blot. In each experiment, the results from three out of six-well plates were averaged and considered as n = 1. No significant variance was observed among the individual wells in each averaged group.
Study of apoptosis
The study of apoptosis was performed with the TACS Annexin V-FITC Apoptosis Detection Kit (R&D Systems). Cells were incubated for 15 min with annexin V-FITC and propidium iodide and subsequently analyzed by flow cytometry using the FACSCalibur (Becton Dickinson, Franklin Lakes, NJ, USA). Analysis was performed using the WinMDI software (Salt Institute, La Jolla, CA, USA). At least 10,000 events were collected for each sample.
Statistical analysis
Data were analysed by unpaired t-test, the paired t-test, as well as analysis of variance (ANOVA) using a confidence level of 95% (P < 0.05). Numerical data were expressed as mean + standard deviation (SD). Statistical significance for mRNA expression analysis between OA and normal samples and for FGF23 protein levels between OA and normal culture media was determined with unpaired t-test. Moreover, sex-related differences between male and female in OA and normal samples were evaluated using two-way ANOVA. Paired t-test was used in order to evaluate statistical significance for mRNA expression analysis after treatment with 1a,25(OH)2D3, Pi, Pi together with PFA, VDR together with PFA. The effect of small interfering RNA (siRNA) against VDR on mRNA expression was evaluated utilizing ANOVA. Statistical significance of apoptosis percentages was evaluated with unpaired t-test. All experiments were performed in triplicate. Statistical analysis was performed using the SPSS software (version 17.0; IBM, Somers, NY, USA).
Results
Evaluation of chondrocyte hypertrophy by COL10A1 expression
All cartilage samples were evaluated for COL10A1 mRNA and protein expression levels. We found that chondrocytes from all OA cartilage samples had significantly higher COL10A1 mRNA (Figure 1a) and protein expression (Figure 1b) levels compared with normal ones (P < 0.0001). Moreover, there was no significant difference between male and female samples.
(a) Quantitative COL10A1 mRNA expression was higher in osteoarthritic compared with normal chondrocytes. GAPDH was used as a normalization control gene for realtime PCR analysis. Data are presented as the mean ± SD, *P < 0.0001. (b) COL10A1 protein expression was increased in osteoarthritic compared with normal chondrocytes. β-Actin was used as a loading control in Western blot analysis. PCR, polymerase chain reaction; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; COL10A1, collagen type X alpha 1; OA, osteoarthritis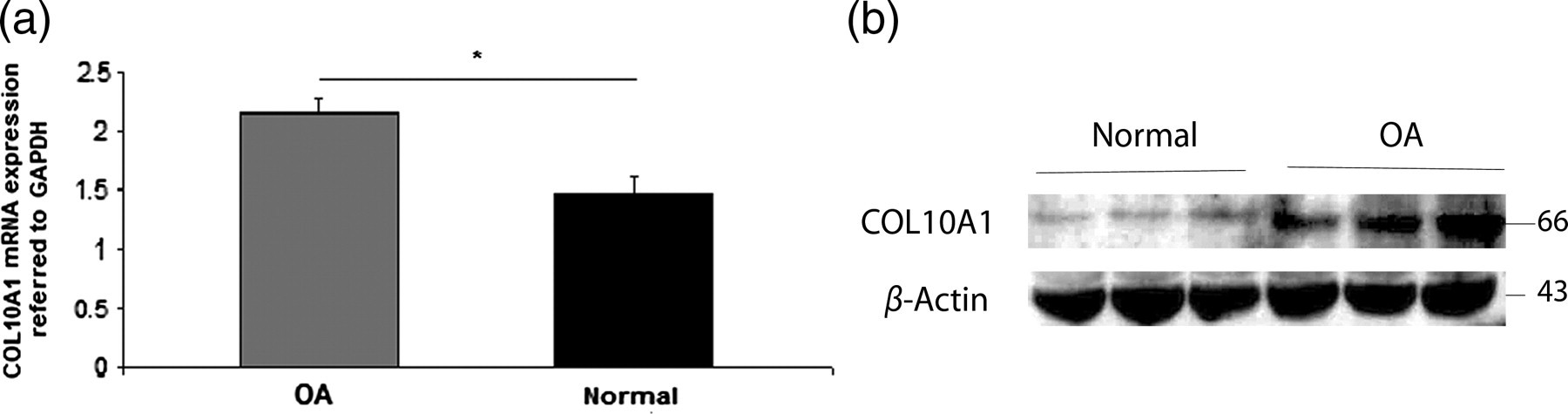
FGF23 and FGFR1c mRNA and protein expression in normal and osteoarthritic chondrocytes
Realtime PCR and Western blot analysis showed that both FGF23 and FGFR1c mRNA and protein levels were significantly higher in OA chondrocytes compared with normal (P < 0.0001) (Figures 2a and b). The statistical significance between male and female samples was also evaluated and there was no significant difference related to sex.
FGF23 and FGFR1c expression in osteoarthritic compared with normal chondrocytes. (a) FGF23 and FGFR1c had higher mRNA expression levels in OA chondrocytes compared with normal. Data are presented as the mean ± SD, *P < 0.0001. (b) FGF23 and FGFR1c had higher protein expression levels in OA chondrocytes compared with normal. β-Actin was used as a loading control in Western blot analysis. (c) Secreted FGF23 levels from primary OA chondrocytes cultures were significantly higher compared with its levels from normal chondrocytes cultures. Data are presented as the mean ± SD, *P < 0.002. FGF23, fibroblast growth factor 23; FGFR1c, fibroblast growth factor receptor 1c; OA, osteoarthritis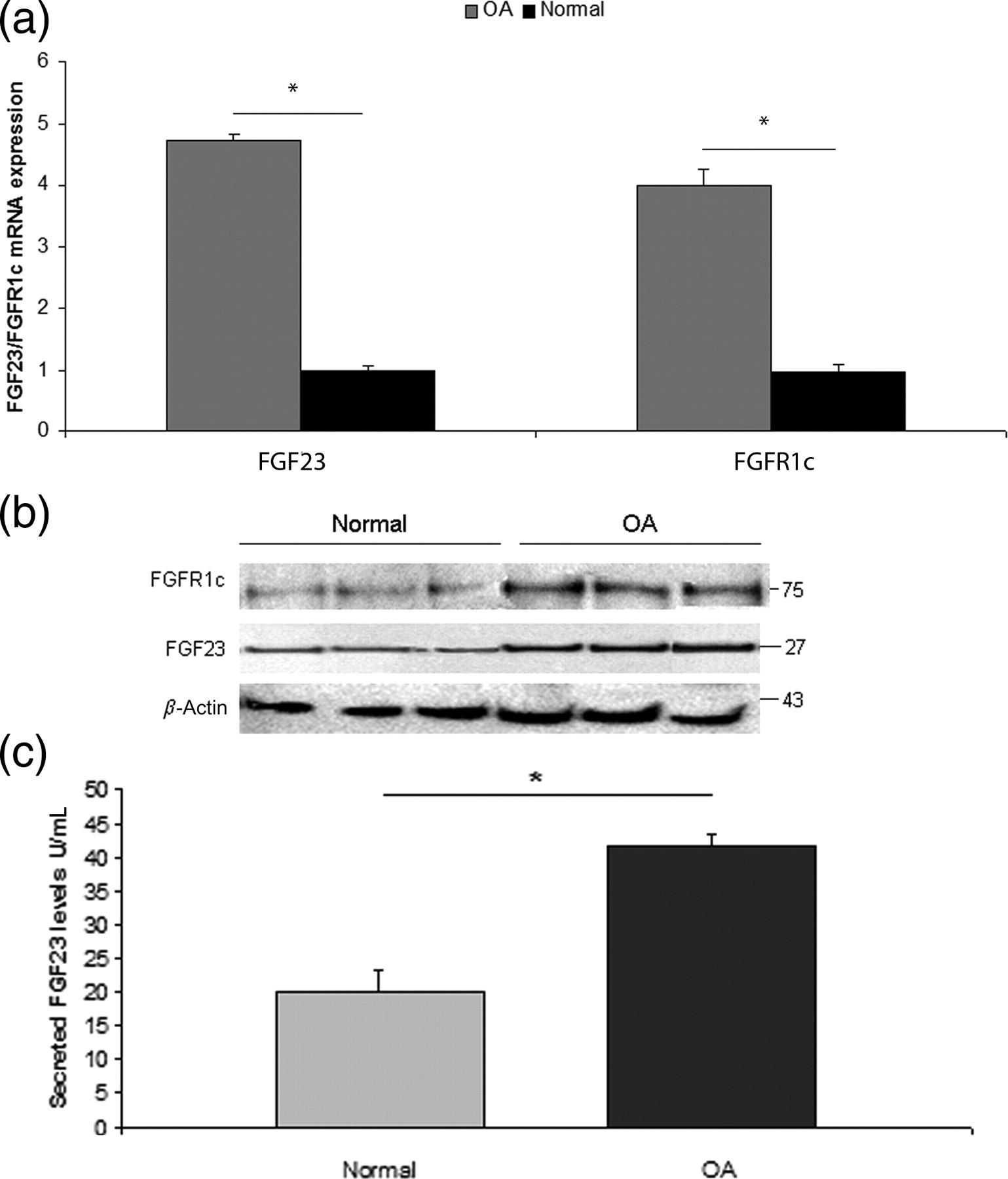
Detection of FGF23 in normal and osteoarthritic culture media
FGF23 levels were evaluated in culture media samples obtained from OA and normal primary chondrocyte cultures. We found that secreted FGF23 levels were significantly higher in OA primary chondrocyte cultures (37–44.81 U/mL, mean 41.61 ± 1.76 U/mL) compared with normal chondrocytes (16.38–26.69 U/mL, mean 20.02 ± 3.34 U/mL) (P < 0.002) (Figure 2c), whereas there was no significant difference in secreted FGF23 levels between male and female samples.
FGF23 and FGFR1c interactions in normal and osteoarthritic chondrocytes
The differences in FGF23 and FGFR1c mRNA and protein expression between OA and normal chondrocytes, prompted us to investigate the interaction between FGF23 and FGFR1c using immunoprecipitation in cultured normal and osteoarthritic chondrocytes. All cultured chondrocytes were at the same passage when harvested in order to avoid altered expression pattern. We observed that FGF23 exhibits stronger binding to FGFR1c in OA chondrocytes compared with normal (Figure 3).
Interactions between FGF23 and FGFR1c in normal and osteoarthritic chondrocytes. In the first row, immunoprecipitation (IP) was performed with antibody against FGF23 and followed by immunoblot (IB) against FGFR1c. In the second row, IP was performed using antibody against FGFR1c and IB with FGF23. IgG is a non-specific human IgG used as a negative control. FGF23, fibroblast growth factor 23; FGFR1c, fibroblast growth factor receptor 1c
VDR expression analysis in normal and osteoarthritic chondrocytes
To test the possible implication of VDR signaling triggered by 1a,25(OH)2D3 in normal and OA chondrocytes, we determined VDR protein expression levels and found that OA chondrocytes exhibited significantly higher VDR protein expression compared with normal (P < 0.0001) (Figure 4).
Quantification of VDR expression levels in OA and normal chondrocytes. VDR protein levels were higher in OA chondrocytes compared with normal. β-Actin was used as a loading control in Western blot analysis. VDR, vitamin D3 receptor; OA, osteoarthritis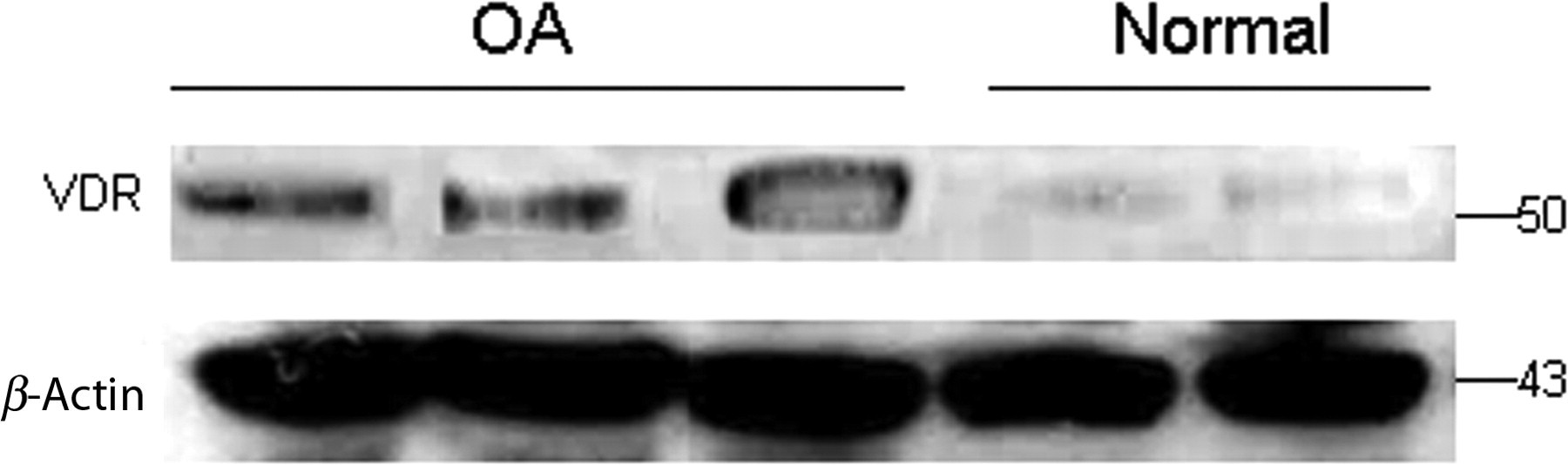
ChiP assay supports the binding of VDR to FGF23 promoter regulating its transcription
The concomitant increased expression of VDR and FGF23 in osteoarthritic chondrocytes prompted us to explore VDR's ability to bind to FGF23 promoter via the possible existence of vitamin D receptor elements (VDREs) and subsequently up-regulate its mRNA expression. ChiP with specific primers for FGF23 promoter revealed that VDR can bind to at least two regions within the FGF23 promoter, situated between −589 and −823 and between −921 and −1094 from the initiator ATG codon. We confirmed that the 1a,25(OH)2D3–VDR complex in osteoarthritic chondrocytes binds to VDRE at −547 bp in the human osteocalcin gene serving as positive control (Figure 5).
Osteoarthritic chondrocytes were subjected to ChIP using antibody against VDR. Amplification of input DNA is also shown. Binding of VDR to osteocalcin gene promoter was used as a positive control. IgG is a non-specific human IgG used as a negative control. DNA precipitates were isolated and then subjected to PCR. Two possible VDREs are situated in FGF23 promoter between -589 and -823 and between -921 and -1094, from the initiator ATG codon. ChIP, chromatin immunoprecipitation; VDR, vitamin D3 receptor; PCR, polymerase chain reaction; VDREs, vitamin D response elements; FGF23, fibroblast growth factor 23; OC, osteocalcin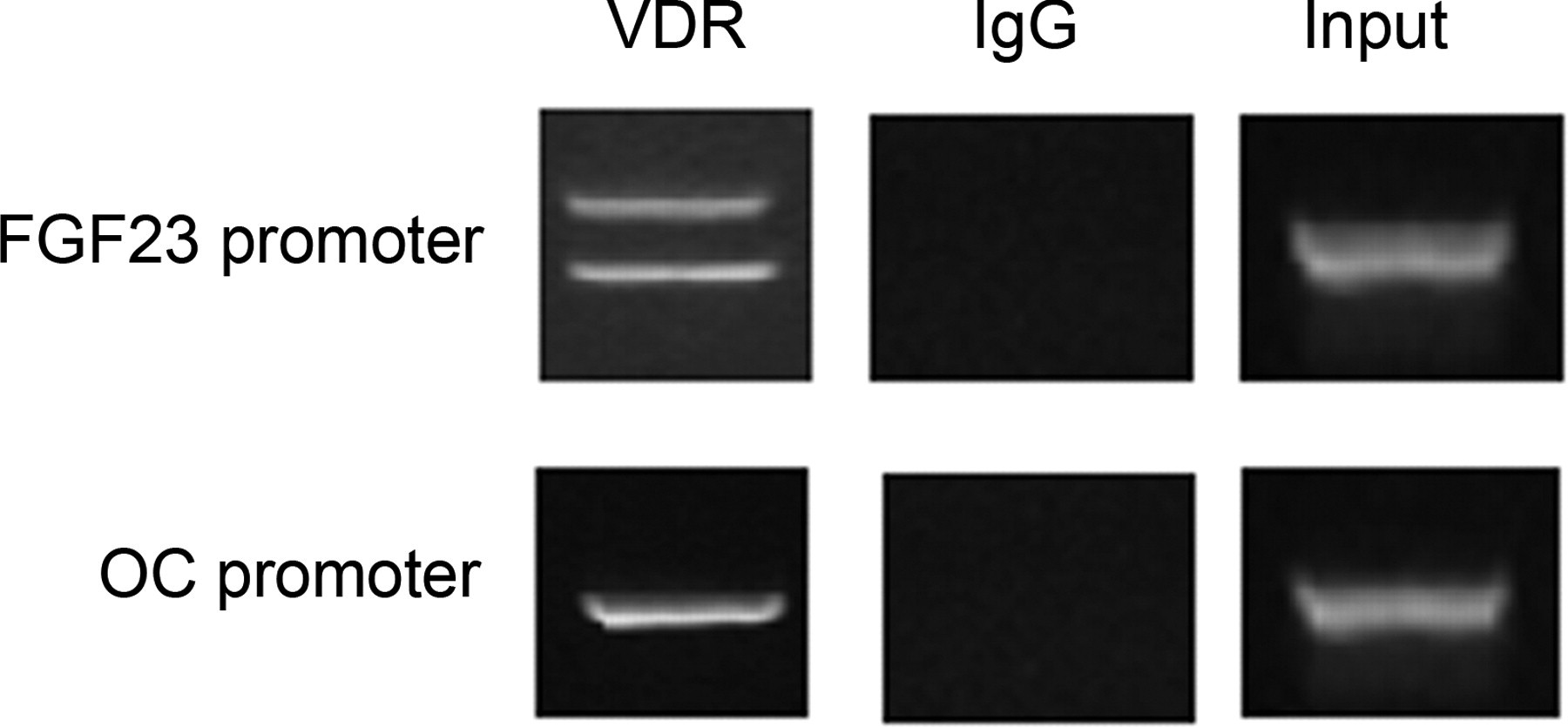
Type III sodium/phosphate cotransporters (PiT-1, PiT-2) expression levels in OA and normal chondrocytes
Driven by the knowledge that increased extracellular Pi leads to PiT-1 up-regulation and Pi influx into chondrocytes, we investigated PiT-1 and PiT-2 mRNA and protein expression levels and found that they were significantly increased in osteoarthritic chondrocytes compared with normal (P < 0.0001) (Figure 6). The statistical significance between male and female samples was also evaluated and there was no significant difference related to sex.
PiT-1 and PiT-2 expression levels in osteoarthritic and normal chondrocytes. (a) PiT-1 mRNA and protein expressions levels were higher in OA compared with normal chondrocytes. (b) PiT-2 had higher mRNA and protein expression levels in OA chondrocytes compared with normal. GAPDH was used as a normalization control gene for realtime PCR analysis and β-actin as a loading control in Western blot analysis. Data are presented as the mean ± SD, *P < 0.0001. PiT, phosphate inorganic transporter; OA, osteoarthritis; PCR, polymerase chain reaction; GAPDH, glyceraldehyde-3-phosphate dehydrogenase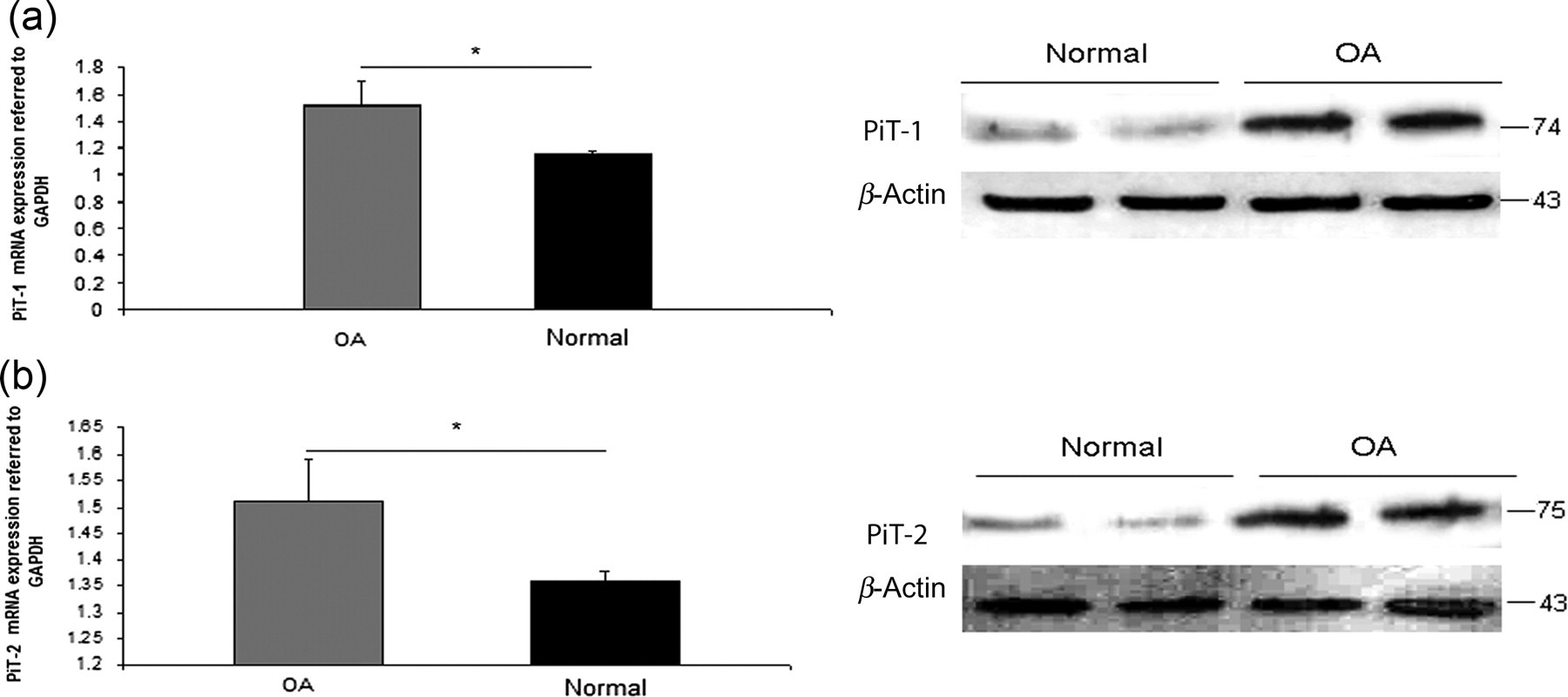
Effect of 1a,25(OH)2D3 in normal chondrocytes
Treatment of normal chondrocytes with 10−7 and 10−6 mol/L 1a,25(OH)2D3 for 24, 48 and 72 h resulted in a time- and dose-significant up-regulation of VDR, FGF23, FGFR1c, OPN, OC, COL10A1 and MMP-13 mRNA and protein levels (P < 0.0001 for all) and caspase-9 and phosphorylated ERK1/2 protein levels (Figure 7).
Treatment of normal chondrocytes with 0, 10−7 and 10−6 mol/L 1,25D at 24, 48 and 72 h. mRNA expressions of (a) VDR, (b) FGF23, (c) FGFR1c, (d) COL10A1, MMP-13, OPN and OC was up-regulated after 10−7 or 10−6 mol/L 1,25D treatment of normal chondrocytes compared with untreated (0 mol/L) at 24, 48 and 72 h. GAPDH was used as a normalization control gene. Data are presented as the mean ± SD, *P < 0.0001 for 0 versus 10−7 mol/L 1,25D at 24, 48 and 72 h, **P < 0.0001 0 versus 10−6 mol/L 1,25D at 24, 48 and 72 h, ***P < 0.0001 10−7 versus 10−6 mol/L 1,25D at 24, 48 and 72 h, ****P < 0.0001 10−7 mol/L at 24 h versus 10−7 mol/L 1,25D at 48 h, 10−7 mol/L at 24 h versus 10−7 mol/L 1,25D at 72 h and 10−7 mol/L at 48 h versus 10−7 mol/L 1,25D at 72 h, *****P < 0.0001 10−6 mol/L at 24 h versus 10−6 mol/L 1,25D at 48 h, 10−6 mol/L at 24 h versus 10−6 mol/L 1,25D at 72 h, 10−6 mol/L at 48 h versus 10−6 mol/L 1,25D at 72 h, NS for non-significant. (e) Increased VDR, FGF23, FGFR1c, OPN, OC, COL10A1, MMP-13 and caspase-9 protein levels after 0, 10−7 and 10−6 mol/L at 72 h treatment in normal chondrocytes normalized to β-actin. 10−7 and 10−6 mol/L 1,25D for 72 h treatment in normal chondrocytes induced pERK1/2 phosphorylation compared with total levels of ERK. 1,25D, 1,25-dihydroxyvitamin D3; VDR, vitamin D3 receptor; OPN, osteopontin; OC, osteocalcin; MMP-13, metalloproteinase-13, ERK, extracellular signal-regulated kinase; FGF23, fibroblast growth factor 23; FGFR1c, fibroblast growth factor receptor 1c; COL10A1, collagen type X alpha 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase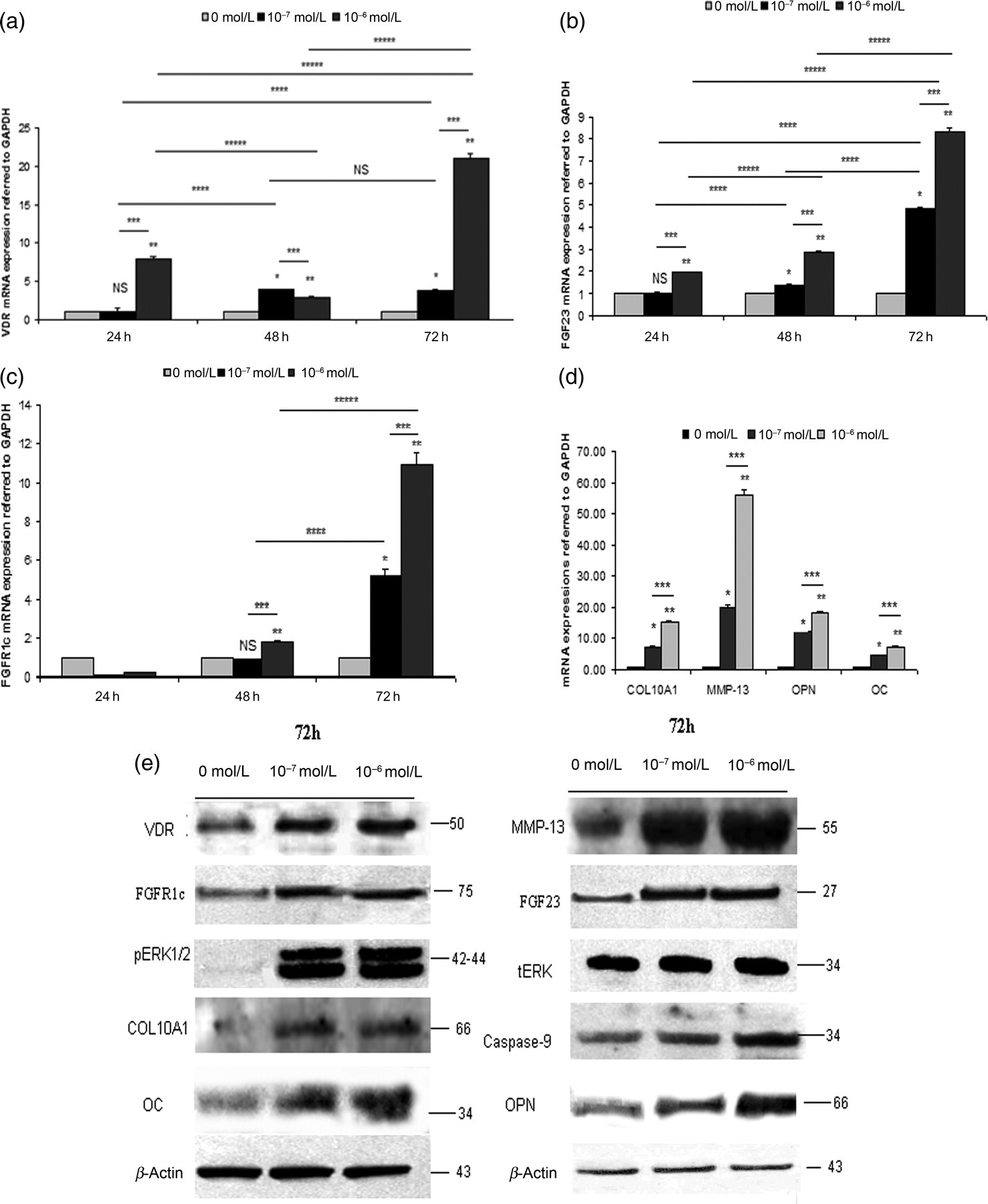
VDR knockdown in osteoarthritic chondrocytes
We next successfully blocked VDR expression in OA chondrocytes using siRNA against VDR and transfection with liposomes and found that VDR mRNA and protein expressions were down-regulated by 97% and 38%, respectively (P < 0.0001) (Figures 8a and b).
Transfection of osteoarthritic chondrocytes with siRNA against VDR. (a) VDR, FGF23, FGFR1c, COL10A1, MMP-13, OPN and OC mRNA expression levels were reduced after 48 h siRNA-VDR. SiRNA-scrambled was used as a control. GAPDH was used as a normalization control gene. *P < 0.0001 for untransfected or siRNA-scrambled versus siRNA against VDR. (b) Reduced VDR, FGF23, FGFR1c, OPN, OC, COL10A1, MMP-13, caspase-9 protein levels after 48 h siRNA-VDR transfection in osteoarthritic chondrocytes normalized to β-actin. siRNA-VDR transfection in osteoarthritic chondrocytes abolished pERK1/2 phosphorylation compared with total levels of ERK. (c) siRNA-VDR transfection for 48 h in OA chondrocytes reduced apoptosis by 4.3-fold compared with untransfected ones or siRNA-scrambled *P < 0.0001. VDR, vitamin D3 receptor; OPN, osteopontin; OC, osteocalcin; MMP-13, metalloproteinase-13, ERK, extracellular signal-regulated kinase; FGF23, fibroblast growth factor 23; FGFR1c, fibroblast growth factor receptor 1c; COL10A1, collagen type X alpha 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase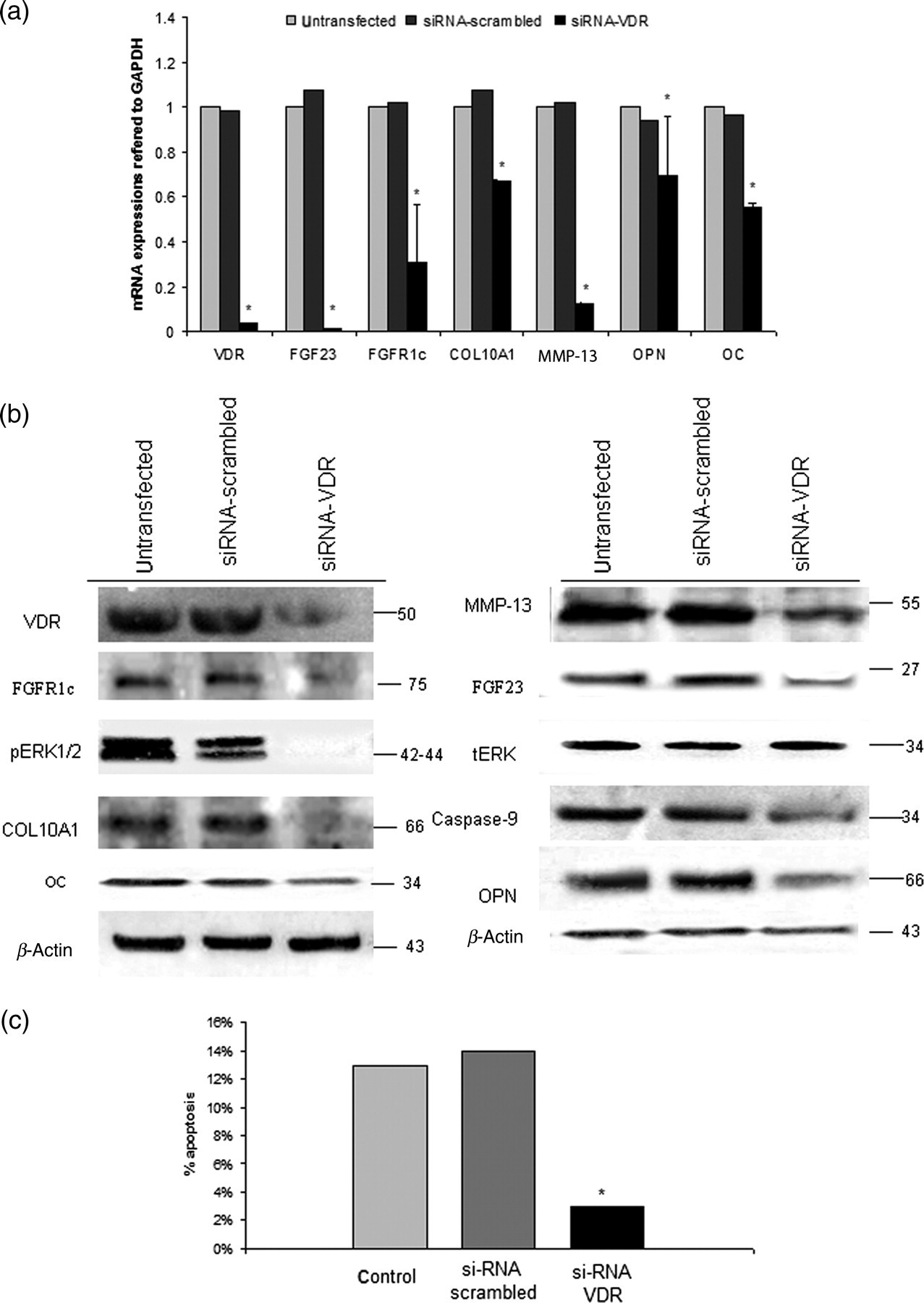
Furthermore, we observed that inhibition of VDR in osteoarthritic chondrocytes resulted in significant down-regulation of FGF23, FGFR1c, OPN, OC, COL10A1 and MMP-13 mRNA and protein expression levels (P < 0.0001) as well as caspase-9 and phosphorylated ERK1/2 protein levels (Figures 8a and b).
Finally, we determined the apoptotic effect of VDR inhibition in OA chondrocytes by annexin V staining and flow-cytometry. Osteoarthritic chondrocytes demonstrated lower apoptotic rates compared with untransfected cells after VDR knock-down (P < 0.0001) (Figure 8c).
Effect of Pi in normal chondrocytes
Treatment of normal chondrocytes with 3, 5 and 7 mmol/L Pi for 48 h resulted in a time- and dose-dependent up-regulation of PiT-1, PiT-2, FGF23, OPN and MMP-13 mRNA and protein levels (P < 0.0001) (Figures 9a and f) as well as caspase-9 and phosphorylated ERK1/2 protein levels (P < 0.0001) (Figure 9f). However, 3, 5 and 7 mmol/L Pi treatment of normal chondrocytes had no significant effect on COL10A1 mRNA expression.
Effect of 3, 5 or 7 mmol/L extracellular Pi in normal chondrocytes. Quantitative mRNA expression of (a) PiT-1, (b) PiT-2, (c) FGF23, (d) OPN, (e) MMP-13 was up-regulated after 3, 5 or 7 mmol/L extracellular Pi treatment of normal chondrocytes compared with control (1 mmol/L Pi) for 48 h. 1 mmol/L PFA pretreatment of normal chondrocytes abolished or reduced the Pi-induction of PiT-1, PiT-2, FGF23, OPN and MMP-13 mRNA expression. GAPDH was used as a normalization control gene. Data are presented as the mean ± SD, *P < 0.0001 for 1 mmol/L versus 3, 5 and 7 mmol/L Pi without PFA,**P < 0.0001 for 1 mmol/L versus 3, 5 and 7 mmol/L Pi with 1 mM PFA, ***P < 0.0001 for 3, 5 and 7 mmol/L Pi without PFA versus 3, 5 and 7 mmol/L Pi with PFA, NS for non-significant. (f) PiT-1, PiT-2, FGF23, OPN, MMP-13 and caspase-9 protein expression levels were induced in the presence of 3, 5 and 7 mmol/L Pi at 48 h in normal chondrocytes compared with control (1 mmol/L Pi). β-Actin was used as a loading control. Phosphorylation of ERK1/2 was increased after 3, 5 and 7 mmol/L Pi at 48 h in normal chondrocytes compared with total levels of ERK. Pi-induced effects were abolished by PFA in normal chondrocytes. (g) Apoptosis was increased after 7 mmol/L Pi in normal chondrocytes compared with control (1 mmol/L Pi), but reduced in the presence of PFA, *P < 0.0001 for 1 mmol/L Pi versus 7 mmol/L Pi, **P < 0.0001 for 1 mmol/L Pi with 1 mmol/L PFA versus 7 mmol/L Pi with 1 mmol/L PFA, ***P < 0.0001 for 7 mmol/L Pi without PFA versus 7 mmol/L Pi with 1 mmol/L PFA, NS for non-significant. Pi, inorganic phosphate; PiT, phosphate inorganic transporter; PFA, phosphonoformic acid; VDR, vitamin D3 receptor; OPN, osteopontin; OC, osteocalcin; MMP-13, metalloproteinase-13, ERK, extracellular signal-regulated kinase; FGF23, fibroblast growth factor 23; GAPDH, glyceraldehyde-3-phosphate dehydrogenase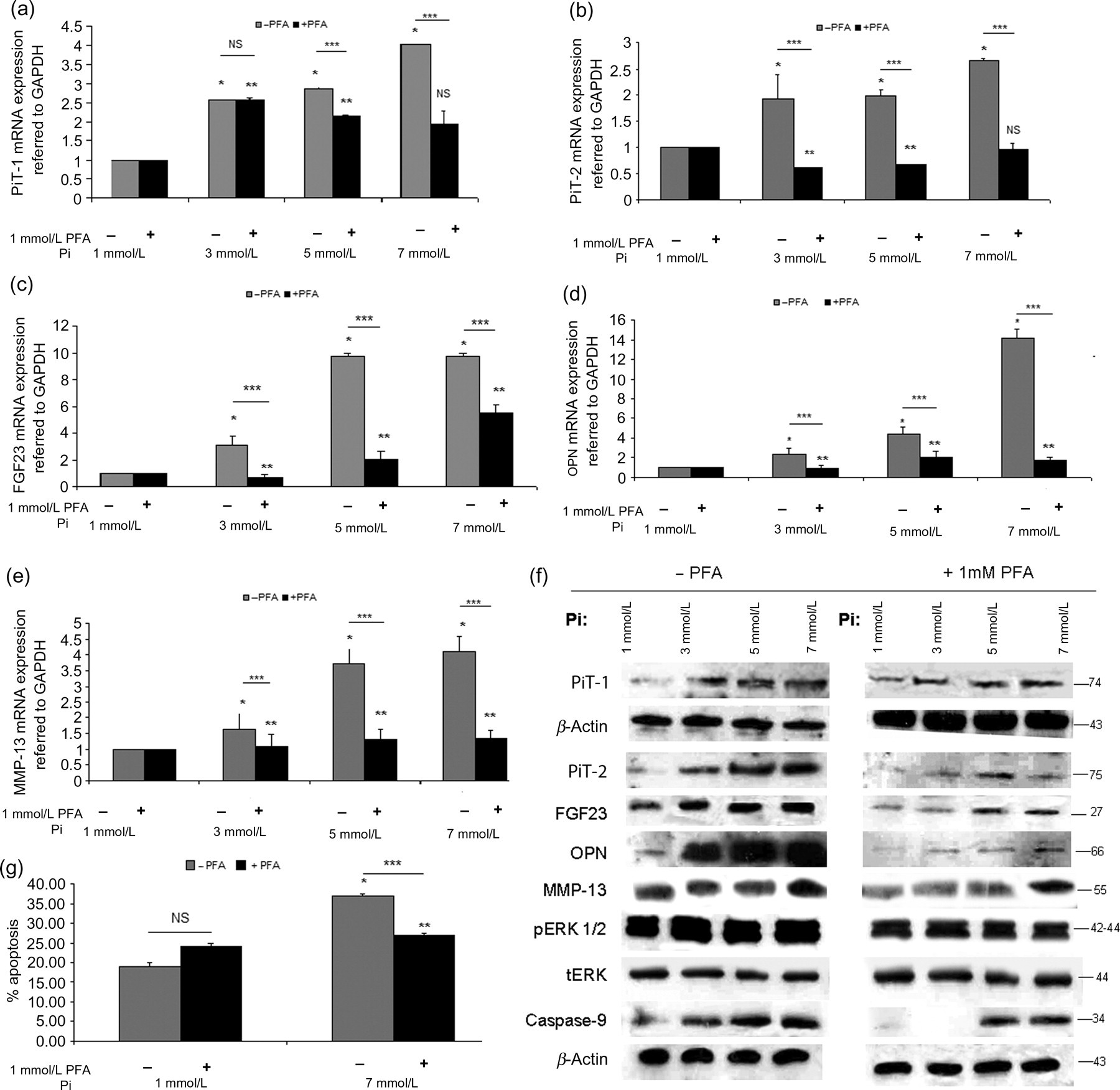
Addition of 1 mmol/L PFA, a synthetic inhibitor of Pi, together with Pi resulted in significant inhibition of PiT-1, PiT-2, FGF23, OPN and MMP-13 mRNA and protein levels and reduced caspase-9 and phosphorylated ERK1/2 protein levels (P < 0.001) (Figures 9a and f).
Moreover, treatment of normal chondrocytes with 7 mmol/L Pi for 48 h resulted in significant increase of apoptotic cells (P < 0.001), whereas the combined treatment of 1 mmol/L PFA together with 7 mmol/L Pi resulted in inhibition of the Pi effect (P < 0.001) (Figure 9g).
1a,25(OH)2D3 and Pi synergistic effect on FGF23 signaling
We proceeded with simultaneous inhibition of VDR and treatment with 1 mmol/L PFA in OA chondrocytes, in order to test the possible synergistic action of VDR silencing and PiT-1, PiT-2 inhibition on FGF23 expression and the subsequent effects on downstream targets. We observed that the combined VDR silencing and PFA treatment had a synergistic effect and resulted in higher FGF23, FGFR1c mRNA and protein down-regulation (P < 0.001) than VDR silencing and PFA treatment alone (Figures 10a and b). In addition, the combined treatment also resulted in significant down-regulation of mRNA and protein expression levels of all target genes – COL10A1, OC, OPN and MMP-13 (Figure 10c).
Synergistic action of siRNA against VDR and PFA in osteoarthritic chondrocytes. (a) FGF23, (b) PiT-1 and PiT-2, (c) FGFR1c, COL10A1, OC, OPN and MMP-13 exhibited lower mRNA expression levels after siRNA-VDR transfection together with 1 mmol/L PFA treatment than siRNA-VDR transfection or 1 mmol/L PFA treatment alone in OA chondrocytes. GAPDH was used as a normalization control gene. Data are presented as the mean ± SD, *P < 0.0001 for untransfected versus siRNA-VDR, **P < 0.0001 for untransfected versus 1 mmol/L PFA, ***P < 0.0001 for untransfected versus 1 mmol/L PFA together with siRNA-VDR, ****P < 0.0001 for siRNA-VDR versus 1 mmol/L PFA together with siRNA-VDR, *****P < 0.0001 for 1 mmol/L PFA versus 1 mmol/L PFA together with siRNA-VDR. (d) FGF23, FGFR1c, MMP-13, PiT-1, PiT-2, COL10A1, OC and OPN protein levels were down-regulated more by siRNA-VDR together with 1 mmol/L PFA treatment in OA chondrocytes than by siRNA-VDR or 1 mmol/L PFA alone. PiT-1 and PiT-2 protein levels were not affected by siRNA-VDR transfection. β-Actin was used as a loading control. Phosphorylation of ERK1/2 was reduced at greater extent by the treatment with siRNA-VDR and 1 mmol/L PFA than by the treatment with siRNA-VDR or 1 mmol/L PFA alone in OA chondrocytes compared with ERK total levels. PiT, phosphate inorganic transporter; PFA, phosphonoformic acid; VDR, vitamin D3 receptor; OPN, osteopontin; OC, osteocalcin; MMP-13, metalloproteinase-13, ERK, extracellular signal-regulated kinase; OA, osteoarthritis; FGF23, fibroblast growth factor 23; FGFR1c, fibroblast growth factor receptor 1c; COL10A1, collagen type X alpha 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase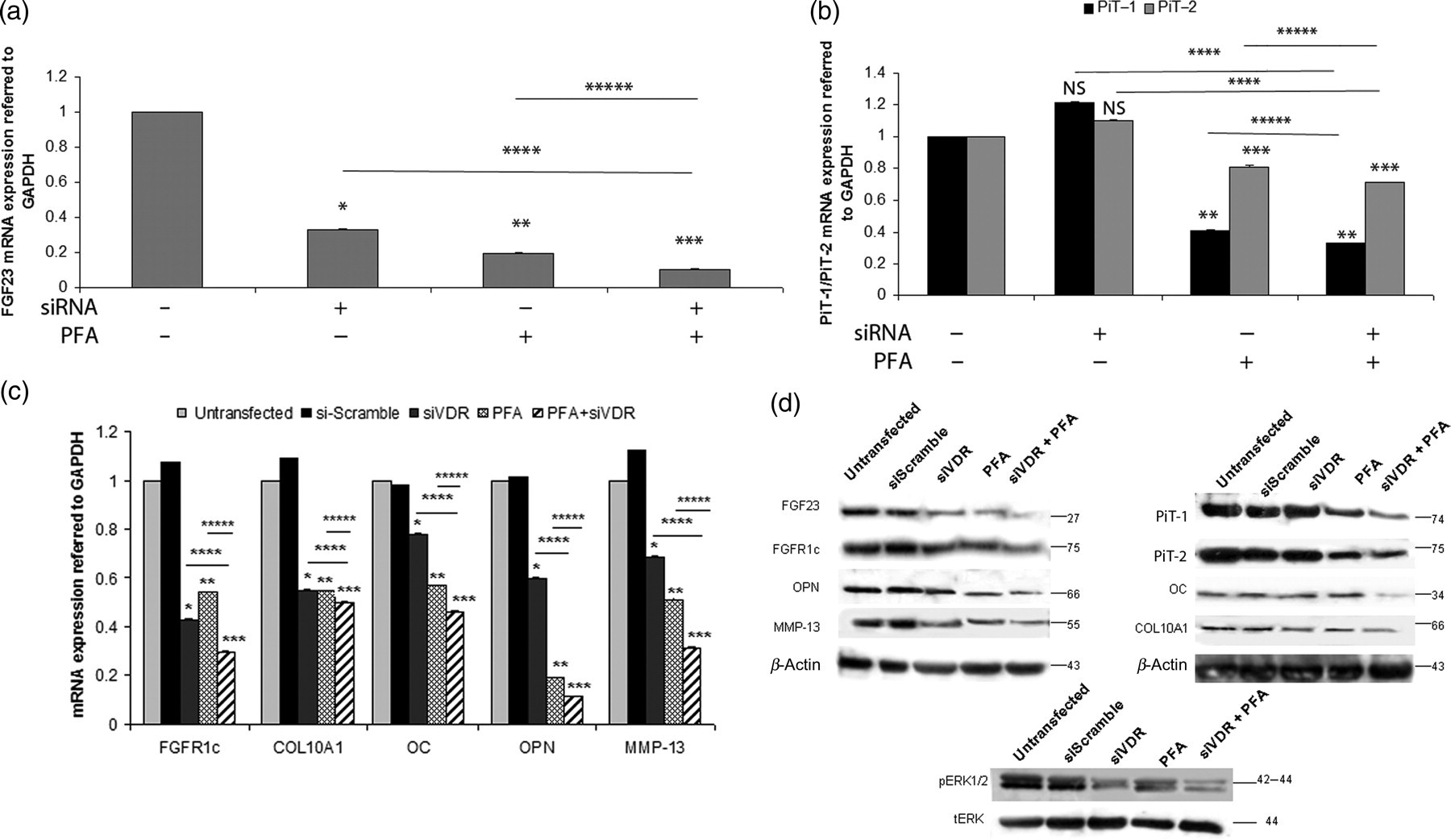
Phosphorylated ERK1/2 protein expression levels showed the strongest down-regulation after combined inhibition of VDR and Na/Pi type III contransporters PiT-1, PiT-2 (Figure 10d).
Discussion
The link between chondrocyte hypertrophy and matrix mineralization is well established in endochondral ossification and in late-stage OA, where it has been associated with disease progression. 30–32 Vitamin D has a recognized role in calcium and phosphorus homeostasis – promoting degrading enzymes and regulating the expression of key target genes such as FGF23.
In the present study, we investigated the role of 1a,25(OH)2D3 and Pi in hypertrophy and mineralization in osteoarthritic chondrocytes mediated by FGF23 signaling through the activation of mitogen-activated protein kinase (MAPK) pathway.
At first, we tested the expression of COL10A1, a hypertrophy marker, in osteoarthritic and normal chondrocytes and found that all osteoarthritic cartilage samples had significantly higher COL10A1 expression levels compared with normal, confirming that all osteoarthritic cartilage samples were hypertrophic.
In order to test the possible implication of VDR, PiT-1 and PiT-2 (Na/Pi type III cotransporters) in the mineralization process in OA, we evaluated their expression levels and found that VDR and PiT-1, PiT-2 had significantly higher mRNA and protein levels in OA chondrocytes compared with normal. Type III Na+/Pi cotransporters, PiT-1 and PiT-2, are widely distributed, as it has been shown that they are expressed in various tissues such as bone and hypertrophic chondrocytes of the growth plate, suggesting that they play a housekeeping role in Pi homeostasis. 17,33,34 In addition, VDR is expressed in the intestine, kidney, bone, bone marrow and chondrocytes and plays critical roles in calcium and phosphorus homeostasis, bone development and mineralization, as well as in the control of cell growth and differentiation. 35 In this study, the observed VDR up-regulation in OA chondrocytes could be attributed to the local production of 1a,25(OH)2D3 within the impaired joint, which influences the production of catabolic enzymes such as MMP-13 and contributes to joint pathology. 36 All previous data together with our results support the concept that VDR and 1a,25(OH)2D3 may play a regulatory role in chondrocytes’ OA phenotype.
As our aim was to investigate the molecular pathway through which 1a,25(OH)2D3 and Pi contribute to hypertrophy in OA chondrocytes, we evaluated FGF23 expression, a circulating phosphaturic factor playing a key role in renal Pi absorption and metabolism, as well as its receptor FGFR1c in osteoarthritic and normal chondrocytes. We found that osteoarthritic chondrocytes exhibited significantly higher FGF23 and FGFR1c mRNA and protein levels compared with normal. It has been shown that FGF23 induces a premature exit of proliferating chondrocytes into hypertrophy, leading to shorter bone growth. 37 Furthermore, we have previously showed that FGF23 positively regulates RUNX-2, a hypertrophy marker in OA chondrocytes. 38 Knowing that FGF23 acts as a circulating phosphaturic hormone, we evaluated its secreted levels in culture media from OA and normal chondrocyte cultures and found that they were significantly elevated in OA compared with normal chondrocytes. The main regulator of FGF23 is 1,25(OH)2D3, which induces its production in osteoblasts in a negative feedback loop. The observed elevated levels of FGF23 in OA chondrocyte culture media could demonstrate that 1,25(OH)2D3 induces its production in order to suppress 1,25(OH)2D3 in OA patients. 23,24,29,39 As FGFR1c is suggested to be the biologically relevant receptor for FGF23, 28 we also tested for the binding of FGF23 to FGFR1c. We observed for the first time, that FGF23 binds to FGFR1c with greater ability in OA than in normal chondrocytes, confirming thus the enhanced activation of FGF23 signaling in OA.
In order to confirm whether FGF23 is a target gene of 1a,25(OH)2D3 in OA chondrocytes, we investigated for the presence of possible VDREs on FGF23 promoter. We identified, for the first time to our knowledge, the presence of two VDREs situated from −589 until −823 and −921 until −1094 in the FGF23 promoter, providing evidence for the effect of 1a,25(OH)2D3 on FGF23 transcriptional regulation during the impaired calcium homeostasis observed in OA.
We next treated normal chondrocytes with 1a,25(OH)2D3 or Pi and we observed increased VDR or PiT-1 and PiT-2 mRNA and protein expression levels, respectively. We also found, for the first time in normal chondrocytes, that both 1a,25(OH)2D3 and Pi resulted in FGF23 up-regulation, suggesting that VDR triggered by 1a,25(OH)2D3 is a transcriptional regulator of FGF23 contributing in mineralization and hypertrophy in OA. In addition, 1a,25(OH)2D3 or Pi treatment in normal chondrocytes resulted in up-regulation of FGFR1c mRNA and protein expression, thus allowing binding with FGF23. Similar results were observed by Kolek et al., 40 who showed that injection of 1a,25(OH)2D3 in mice markedly increased FGF23 expression in several tissues, including the bone, acting as a negative regulator of the active form of vitamin D and preventing ectopic calcification.
Interestingly, we also observed that treatment of normal chondrocytes with 1a,25(OH)2D3 or extracellular Pi resulted in significant phosphorylation of ERK1/2, indicating a central role of the MAPK pathway in signal transduction by extracellular Pi or 1a,25(OH)2D3 in chondrocytes. Recent studies have showed that ERK phosphorylation is associated with maturation of hypertrophic chondrocytes in murine metatarsal cultures and that extracellular calcium stimulates VDR expression in parathyroid glands through the ERK1/2 pathway. 41,42 Moreover, it was shown in osteoblastic MC3T3-E1 cells and in chondrocytic ATDC5 cells, that increase in extracellular Pi induced the phosphorylation of ERK1/2 in order to up-regulate mineralization-related genes, 13,43 suggesting that activation of the MAPK pathway by extracellular Pi or 1a,25(OH)2D3 might regulate different target genes among different cell types.
Furthermore, our observation that the MAPK pathway is activated by extracellular Pi or 1a,25(OH)2D3, led us to investigate the expression of several target genes that could reflect the induced hypertrophy and impaired mineralization observed in OA. We found that hypertrophy markers (COL10A1, OPN and OC), catabolic enzymes (MMP-13) and apoptotic proteins (caspase-9) were significantly up-regulated in a time- and dose-dependent manner after Pi or 1a,25(OH)2D3 treatment. Moreover, we observed that Pi treatment in normal chondrocytes resulted in an increased number of apoptotic cells.
To confirm our previous findings, we knocked down VDR expression in osteoarthritic chondrocytes using siRNA against VDR. We found that silencing of VDR expression resulted in significant down-regulation of FGF23 and FGFR1c mRNA and protein expression levels and subsequent reduction of phosphorylated ERK1/2, suggesting, for the first time, a direct role of 1a,25(OH)2D3 on FGF23 signaling through ERK1/2 phosphorylation in osteoarthritic chondrocytes. In addition, we observed that VDR silencing in OA chondrocytes resulted in significant down-regulation of COL10A1, OPN, OC, MMP-13 and caspase-9 protein levels, as well as in reduced apoptotic rate, confirming the implication of 1a,25(OH)2D3 in the impaired mineralization and hypertrophy in OA. Moreover, treatment of normal chondrocytes with Pi together with its synthetic inhibitor PFA resulted in significant reduction of Pi-induced responses, suggesting that the effects of Pi on chondrocytes were specific and dependent on the active transport of the ion through the Na/Pi type III cotransporters, PiT-1 and PiT-2.
Finally, we investigated the convergence point of the signaling triggered by extracellular Pi and 1a,25(OH)2D3 and found that inhibition of both signaling pathways using PFA against Na/Pi type III cotransporters and siRNA against VDR had a synergistic effect and resulted in additive down-regulation of FGF23 and FGFR1c expression levels as well as ERK1/2 phosphorylation. It has been previously shown in growth-plate hypertrophic chondrocytes that 1a,25(OH)2D3 and Pi interact in a feedback loop in which 1a,25(OH)2D3 contributes to Pi release into the ECM and its subsequent action on chondrocyte-inducing apoptosis. 7 We finally observed that the combined treatment with siRNA against VDR and PFA resulted in significant down-regulation of late hypertrophic (COL10A1, OPN, OC) and catabolic (MMP-13) markers, compared with each inhibitor alone. It is interesting that OPN, a phosphoprotein, exhibited the strongest down-regulation. OPN has been shown to play a pivotal role in the differentiation of late hypertrophic murine chondrocytes requiring ERK1/2 activation by Pi to regulate its expression. 43
In conclusion, we provide evidence that Pi and 1a,25(OH)2D3 activate ERK1/2 through FGF23 signaling and are implicated in hypertrophy and impaired mineralization in osteoarthritic chondrocytes, suggesting that VDR/Pi signaling may be potential targets for therapeutic intervention in late-stage OA.