
Abstract
Capsaicin, a potent algogen, induces an itch-related behavior in the presence of inflammation. In this study, we tested whether bradykinin (BK) can evoke a similar response and investigated the potential mechanisms involved in this process. Local inflammation was induced by intradermal injection of complete Freund's adjuvant (CFA) into the back of the neck, left hind foot or left cheek of male C57BL/6J mice. BK was then injected intradermally into the same area on indicated days. Four days after CFA inflammation, BK treatment evoked scratching responses in a time- and dose-dependent manner. For BK receptor antagonist treatment, inflamed-mice were either given an intraperitoneal injection of B1 receptor (B1R) or B2 receptor (B2R) antagonist 30 min prior to BK administration, or an intradermal co-injection of antagonist and BK into the inflamed area. Our results indicate that B1R and B2R act in an opposite fashion during this process, as pretreatment with B1R antagonist by intraperitoneal injection significantly reduced BK-induced scratching behavior, whereas B2R antagonist treatment dramatically increased scratching behavior. Moreover, combined injection of BK and B2R antagonist enhanced BK-induced scratching activity in CFA-inflamed mice. In addition, pretreatment or co-injection with B2R antagonist dramatically reduced the pain-related licking behavior induced by BK injection. The data suggest that BK-induced scratching responses in CFA-inflamed mouse skin occur via activation of B1R. Furthermore, B1 and B2 receptors play different roles in modulating BK-induced itch-related behavior in CFA-inflamed mice.
Introduction
Itching, or pruritus, is an unpleasant sensory experience that usually accompanies various skin diseases such as atopic dermatitis. 1 Itching typically involves scratching behavior, which is considered as a self-defense mechanism to prevent potential injury. 2 Based on the clinical signs and distinguishing characteristics of skin lesion diseases, itching is generally classified into three categories: itching in inflamed (diseased) skin; itching in non-inflamed (non-diseased) skin; and itching presenting with severe chronic secondary scratch lesions. 3 A variety of stimuli generated within or applied to the skin can induce itching, including inflammatory infections, autoimmune cutaneous diseases, genodermatoses, drug reactions and skin lymphomas. 3 Most importantly, the primary skin disease may be aggravated by secondary scratch lesions. However, the pathophysiological basis of itching is largely unknown.
Various substances have been proven to induce itching, and histamine, which is released from mast cells in an early phase of inflammation, has been recognized as an essential mediator of itching sensations in inflamed human skin. 4 Furthermore, the active peptide bradykinin (BK), which is isolated from plasma globulins treated with either trypsin or a snake venom protease, 5 has been linked to a wide range of biological phenomena such as pain and inflammation. 6 Notably, BK is also known to be one of the most potent endogenous algogenic substances, with the ability to act as a potent histamine-independent pruritogen in the lesional skin of atopic dermatitis. 7 We have reported that capsaicin induced an itch-related response in the presence of inflammation evoked by complete Freund's adjuvant (CFA). 8 It is still unclear whether BK can evoke itch-related scratching behaviors in such situations.
BK exerts its biological effects through the activation of two seven-transmembrane G-protein-coupled receptors, termed bradykinin B1 receptor (B1R) and B2 receptor (B2R). 6 The upregulation of B1R has been detected following treatment with CFA, endotoxins and cytokines, 9,10 suggesting a close correlation between BK receptors and inflammation. Moreover, B1R contributes to acute pain following extraction of third molars. 11 In the present study, we observed BK-induced scratching responses in CFA-inflamed mice and demonstrated that a BK receptor-mediated pathway was involved in this process. Interestingly, we found that B1R and B2R play different roles in modulating BK-induced pruritus-related behavior in CFA-inflamed mice.
Materials and methods
Reagents
BK was purchased from Tocris (Bristol, UK). Transient receptor potential vanilloid subfamily 1 (TRPV1) antagonist (capsazepine), CFA, B1 kinin receptor agonist (des-Arg9-bradykinin) and B1 kinin receptor antagonist (des-Arg9-[Leu8]-bradykinin, DALBK) were obtained from Sigma Chemical Co (St Louis, MO, USA). B2 kinin receptor antagonist (
Animals
Male C57BL/6J mice, weighing 20–22 g, were obtained from the Center for Laboratory Animals, Sun Yat-sen University (Guangzhou, China). The animals were housed at 22 ± 1°C on a 12/12-h light/dark cycle, with free access to food and water. The animal use and care protocols were approved by the Committee on Ethical Use of Animals of Guangdong General Hospital (Guangzhou, China), following the National Institutes of Health (NIH) Animal Use and Care Guidelines. All efforts were made to minimize animal suffering and to reduce the number of animals used.
Development of local inflammation
To eliminate the influence of shaving on background scratching, mice were shaved at the rostral part of the back of the neck or the left cheek where intradermal injections were then given one day prior to experiments. Animals were randomly divided into two groups, an inflammation group (CFA-inflamed group) and a normal saline group (NS group), and were routinely given 30 min to acclimate in a small plastic chamber (22 cm×12 cm×20 cm) before injection. To induce local inflammation, mice in the inflammation group were removed from the chamber and received an intradermal injection of 50, 10 or 10 μL of CFA into the rostral back area (neck), the left cheek or the left hind foot, respectively, with a 30-gauge needle. In control groups, an identical volume of NS was administrated instead of CFA.
BK-induced scratching behavior with back injection
To evaluate BK-induced scratching behavior, between 0 and 1 mmol/L of BK was injected intradermally into the back of the neck on the indicated days after CFA or NS administration. For TRPV1 antagonist and BK receptor antagonist treatment, animals in the inflammation group were given intraperitoneal injections of capsazepine (4 mg/kg body weight), B1R antagonist (DALBK; 0.4, 2 or 10 mg/kg body weigh) or B2R antagonist (
BK-induced pain-related behavior with paw injection
Four days after intradermal injection of 10 μL CFA or NS into the left hind feet, mice were given NS, BK (1 mmol/L) or B1R agonist (0.4 mmol/L) in a volume of 10 μL. For BK receptor antagonist treatment, inflamed animals were given an intraperitoneal injection of B1R or B2R antagonist (10 mg/kg) 30 min prior to BK (1 mmol/L in 10 μL) administration to the hind feet. The combination treatment consisted of co-injection of B1R or B2R antagonist (1 nmol) with BK (1 mmol/L in 10 μL) into the left hind feet of CFA-inflamed mice. Animals were placed in individual cages and the time spent licking the injected hind foot was measured with a chronometer for five minutes after the final injection. Licking represented a response to a painful stimulus.
Cheek injection assessment and behavioral analysis
NS, BK (1 mmol/L) or B1R agonist (0.4 mmol/L) in a volume of 10 μL was injected intradermally into the left cheek on the indicated days after CFA or NS administration following the method introduced by Shimada and LaMotte. 14 The number of hindpaw scratches directed to the injected area and the number of ipsilateral forelimb wipes directed to the injected area as indicators of itch and pain were recorded for a period of 30 min after the final injection.
Histological assessment of inflammation
Skin from the back of the neck was harvested on day 4 after the induction of inflammation. Tissue was fixed in 10% neutral-buffered formalin for 48 h. Fixed tissues were dehydrated in a graded series of ethanol, cleared in xylene, infiltrated, then processed using a Tissue-Tek VIP automatic tissue processor (Sakura, Tokyo, Japan) with a standard protocol and embedded into paraffin wax (Tissue-Tek). Tissue blocks were sectioned at 5 μm, and slides were stained with hematoxylin—eosin. Tissue samples were evaluated by an experienced pathologist.
Statistical analysis
All data were plotted as mean ± SEM. Data were statistically evaluated by analysis of variance (ANOVA) followed by Bonferroni's test or, when only two means were to be compared, unpaired Student's t-test. A P value < 0.05 was considered statistically significant.
Results
Scratching response induced by BK in CFA-inflamed skin
Local inflammation was induced in mice by a 50 μL intradermal injection of CFA into the back of the neck. As shown in Figure 1, CFA-induced scratching activity occurred immediately after injection and subsided within 72 h (3 d). Scratching frequency was very low in NS-injected mice (Figure 1, non-inflammation group). To eliminate the influence of CFA on drug-stimulated scratching behavior, drugs were administrated four days after CFA inflammation. We next monitored the BK-stimulated scratching behavior in mice after four days of CFA inflammation. Dose-dependent scratching activity was observed when BK was injected intradermally into the back of the neck (Figure 2a). There was a significant difference (P < 0.05) in scratching frequency between animals receiving NS and BK treatments at all doses examined. To determine the correlation between BK-evoked scratching behavior and the duration of CFA-induced inflammation, mice were given 0.5 mmol/L BK via intradermal injection on each indicated day after CFA administration. We found that the scratching frequency induced by BK gradually dropped over time after 14 d of CFA inflammation (Figure 2b). These results demonstrate that BK treatment evoked time- and dose-dependent scratching responses in CFA-inflamed mice.
CFA-induced scratching behavior. Mice received an intradermal injection (50 μL) of CFA (inflammation group) or normal saline (non-inflammation group) into the back of the neck. The hind limb scratching behavior directed toward the injected area was recorded for a period of 30 min at 0, 6, 24, 48, 72 or 96 h after injection. Data are expressed as mean ± SEM (n = 10 in each group). *P < 0.05 compared with the NS group. CFA, complete Freund's adjuvant; NS, normal saline BK-induced scratching behavior. (a) Four days after CFA or normal saline administration, mice were given different doses of BK (from 0 to 1 mmol/L, 50 μL) by intradermal injection into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 8 in each group). *P < 0.05 compared with the non-inflammation group. (b) On each indicated day after CFA administration, mice were given 50 μL of 0.5 mmol/L BK by intradermal injection into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 6 in each group). *P < 0.05 compared with injection of 0.5 mmol/L BK four days after normal saline administration. CFA, complete Freund's adjuvant; BK, bradykinin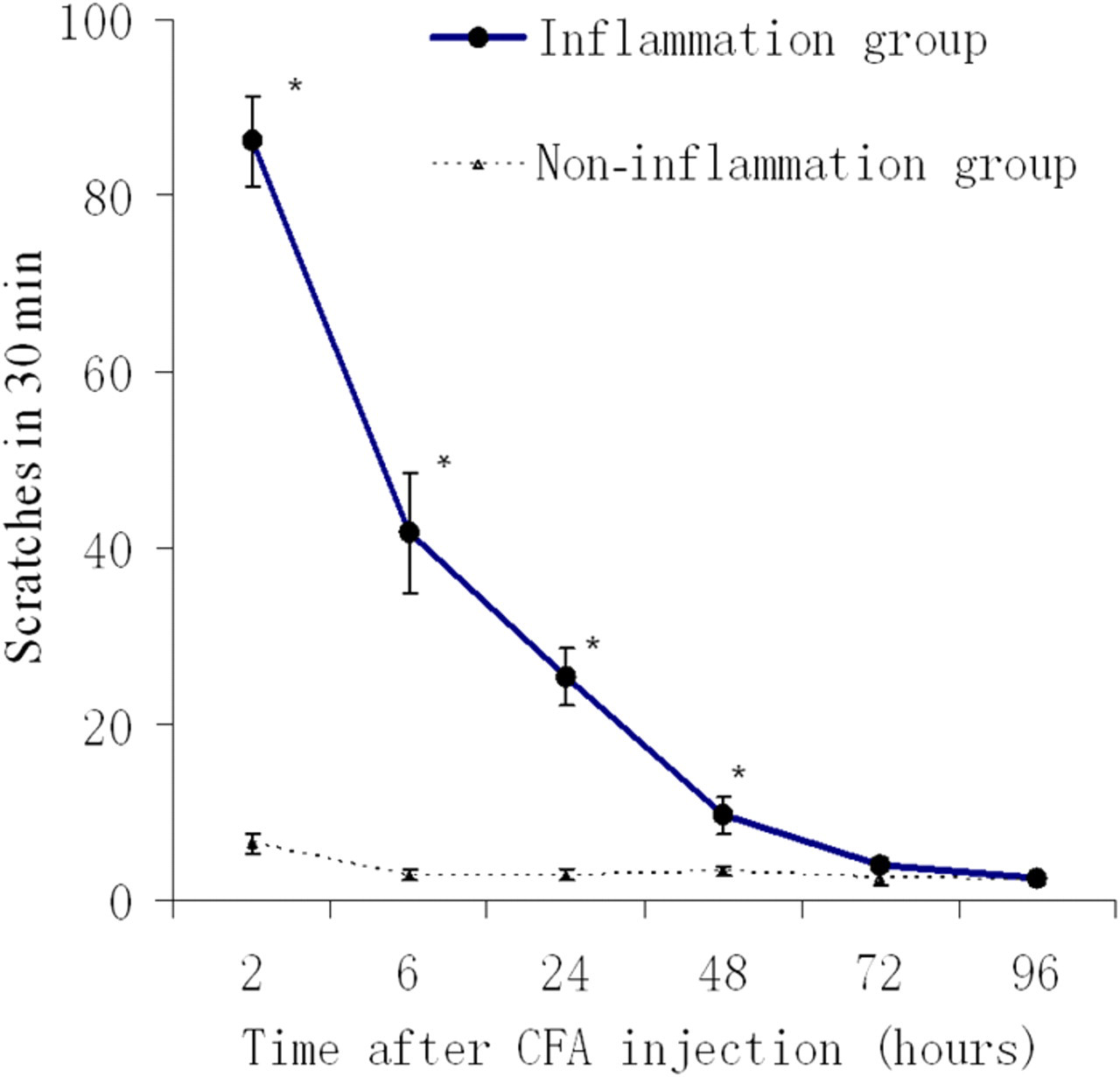

The effects of BK receptor antagonist on BK-induced scratching behavior
To evaluate the effects of BK receptor antagonist on BK-induced scratching activity, B1R (DALBK) and B2R antagonists ( Effects of BK receptor antagonist on BK-induced scratching behavior. (a) Four days after CFA inflammation, mice were given an intraperitoneal injection of B1R or B2R antagonist (0.4, 2 or 10 mg/kg body weight) 30 min prior to BK (0.5 mmol/L, 50 μL) or normal saline (50 μL) injection into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 8 in each group). *P < 0.05 compared with BK injection after intraperitoneal injection of normal saline (NS) in CFA-treated mice. (b) After four days of CFA inflammation, B1R, B2R antagonist (5 nmol) or NS were intradermally co-injected with BK (0.5 mmol/L) in a total volume of 50 μL into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 8 in each group). *P < 0.05 compared with co-injection of solvent and BK in CFA-treated mice. CFA, complete Freund's adjuvant; BK, bradykinin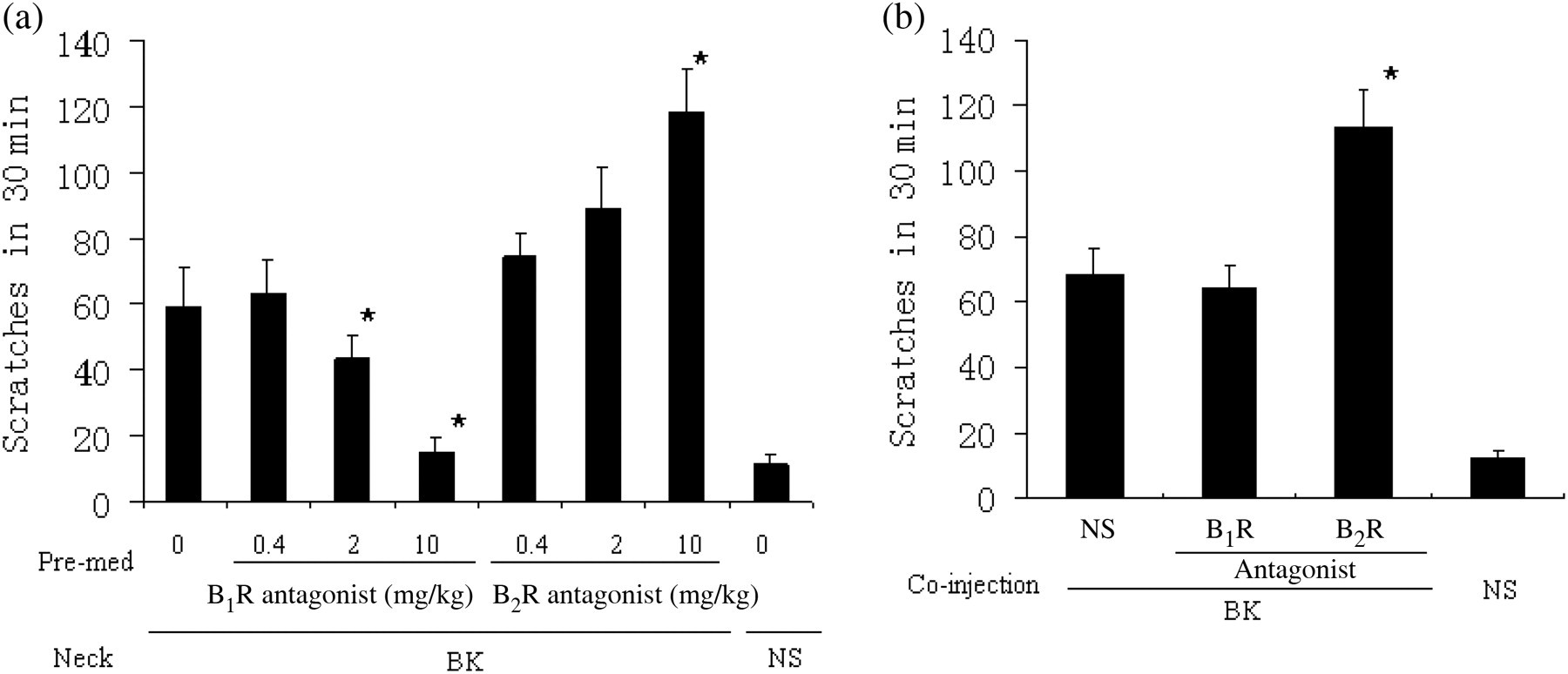
The effects of capsazepine on BK-induced scratching behavior
Antagonism or gene deletion of B1R can lead to the abolishment of phorbol myristate acetate (PMA)-induced nociceptive behavior.
17
This behavior is also reduced in TRPV1 gene knockout mice.
18
In addition, BK-induced pain in mice can be dose-dependently prevented by both B2R antagonist and TRPV1 antagonist.
19
To evaluate the effects of TRPV1 antagonist on BK-induced scratching activity, mice were given an intraperitoneal injection of capsazepine (4 mg/kg in 1% DMSO) or vehicle (1% DMSO) prior to BK administration. Pretreatment with systemically administered capsazepine did not significantly affect the number of scratches induced by BK (Figure 4). The dose of capsazepine was sufficient to inhibit the scratching response induced by capsaicin in CFA-treated mice.
8
These results suggest that activation of B2R inhibits the scratching response in CFA-inflamed skin, possibly through a non-TRPV1 pathway. However, the underlying mechanism should be elucidated further.
Effects of capsazepine on BK-induced scratching behavior. After four days of CFA inflammation, mice were given an intraperitoneal injection of TRPV1 antagonist (capsazepine, 4 mg/kg body weight in 1% DMSO) or vehicle (1% DMSO) 30 min prior to BK (0.5 mmol/L, 50 μL) injection into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 8 in each group). CFA, complete Freund's adjuvant; BK, bradykinin; DMSO, dimethylsulfoxide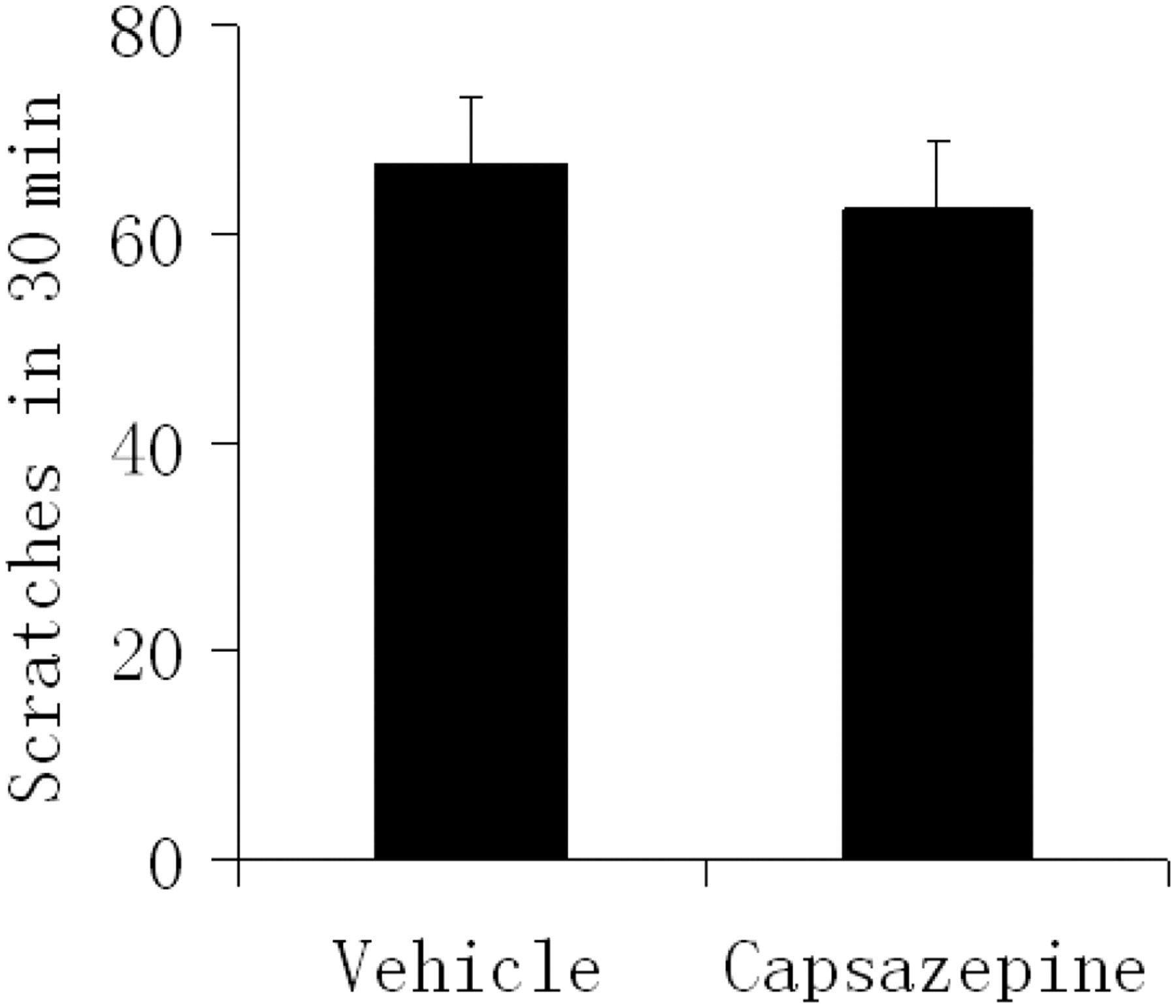
Scratching behavior induced by B1R agonist and the role of BK receptor antagonists
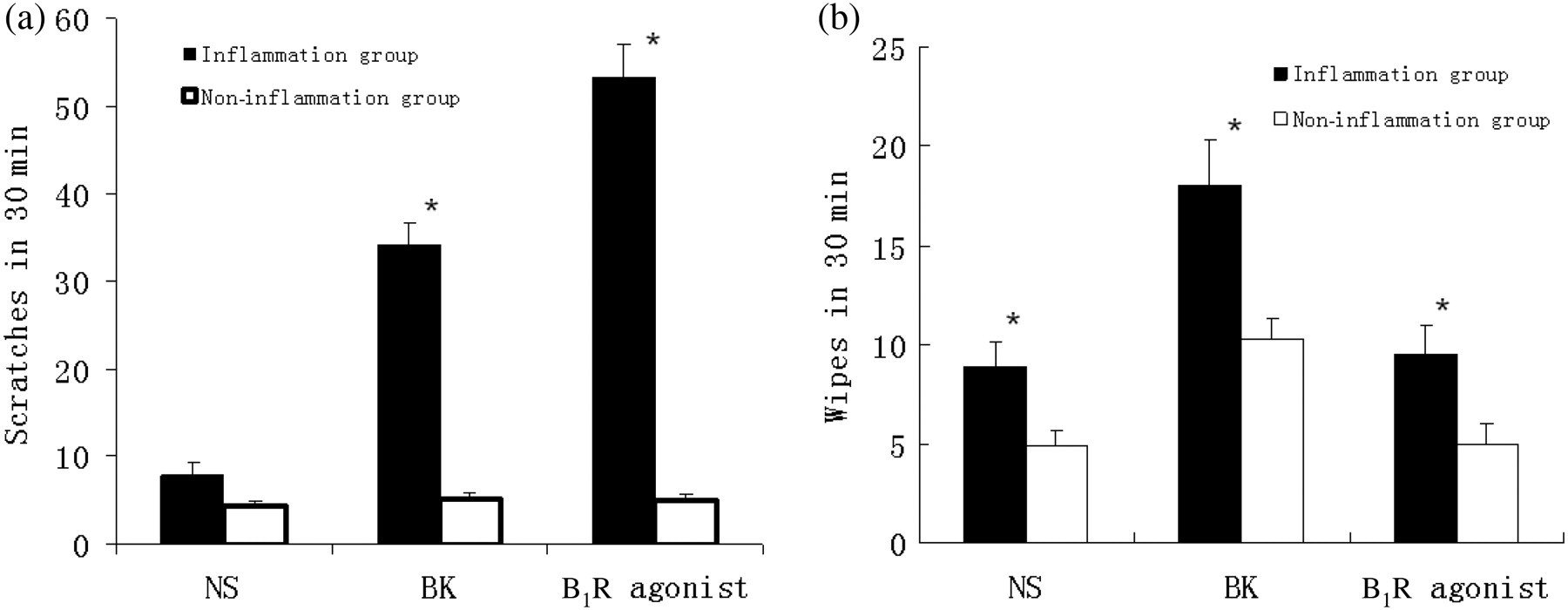
We further analyzed the potential influence of a combination treatment with selective B1R agonist and BK receptor antagonists on CFA-inflamed scratching behavior. The optimal concentration of a widely used B1R agonist (des-Arg9-bradykinin) was determined via intradermal injection at doses ranging from 0 to 2 mmol/L in CFA-inflamed mice. B1R agonist-treated mice exhibited elevated scratching behavior at doses over 0.016 mmol/L, with no additional increases observed at 0.4 and 2 mmol/L agonist application (Figure 5a). Therefore, we used the 0.4 mmol/L dose. Either B1R or B2R antagonist (10 mg/kg body weight) was injected into CFA-inflamed mice intraperitoneally 30 min prior to intradermal injection of B1R agonist (0.4 mmol/L), or the receptor antagonists were co-injected with B1R agonist (0.4 mmol/L) into CFA-inflamed mice. We found that a 30-min pretreatment with B1R antagonist suppressed the scratching behavior induced by B1R agonist, whereas no obvious effect was observed upon B2R antagonist treatment or B1R antagonist co-injection (Figures 5b and c).
Effects of B1R agonist and BK receptor antagonist on scratching behavior. (a) After four days of CFA inflammation, mice were given B1R agonist (des-Arg9-bradykinin) at a dose of 0, 0.016, 0.08, 0.4 or 2 mmol/L (in 50 μL) by intradermal injection into the back of the neck. Data are expressed as mean ± SEM (n = 8 in each group). *P < 0.05 compared with 0 mmol/L. (b) Four days after CFA inflammation, mice were given an intraperitoneal injection of B1R or B2R antagonist (10 mg/kg body weight) 30 min prior to B1R agonist (0.4 mmol/L in 50 μL) or normal saline (50 μL) injection into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 8 in each group). *P < 0.05 compared with B1R agonist injection after intraperitoneal injection of normal saline (NS) in CFA-treated mice. (c) After four days of CFA inflammation, B1R, B2R antagonist (5 nmol) or NS were intradermally co-injected with B1R agonist (0.4 mmol/L) in a total volume of 50 μL into the back of the neck. The scratching activity was recorded for a period of 30 min. Data are expressed as mean ± SEM (n = 8 in each group). *P < 0.05 compared with co-injection of solvent and B1R agonist in CFA-treated mice. CFA, complete Freund's adjuvant; BK, bradykinin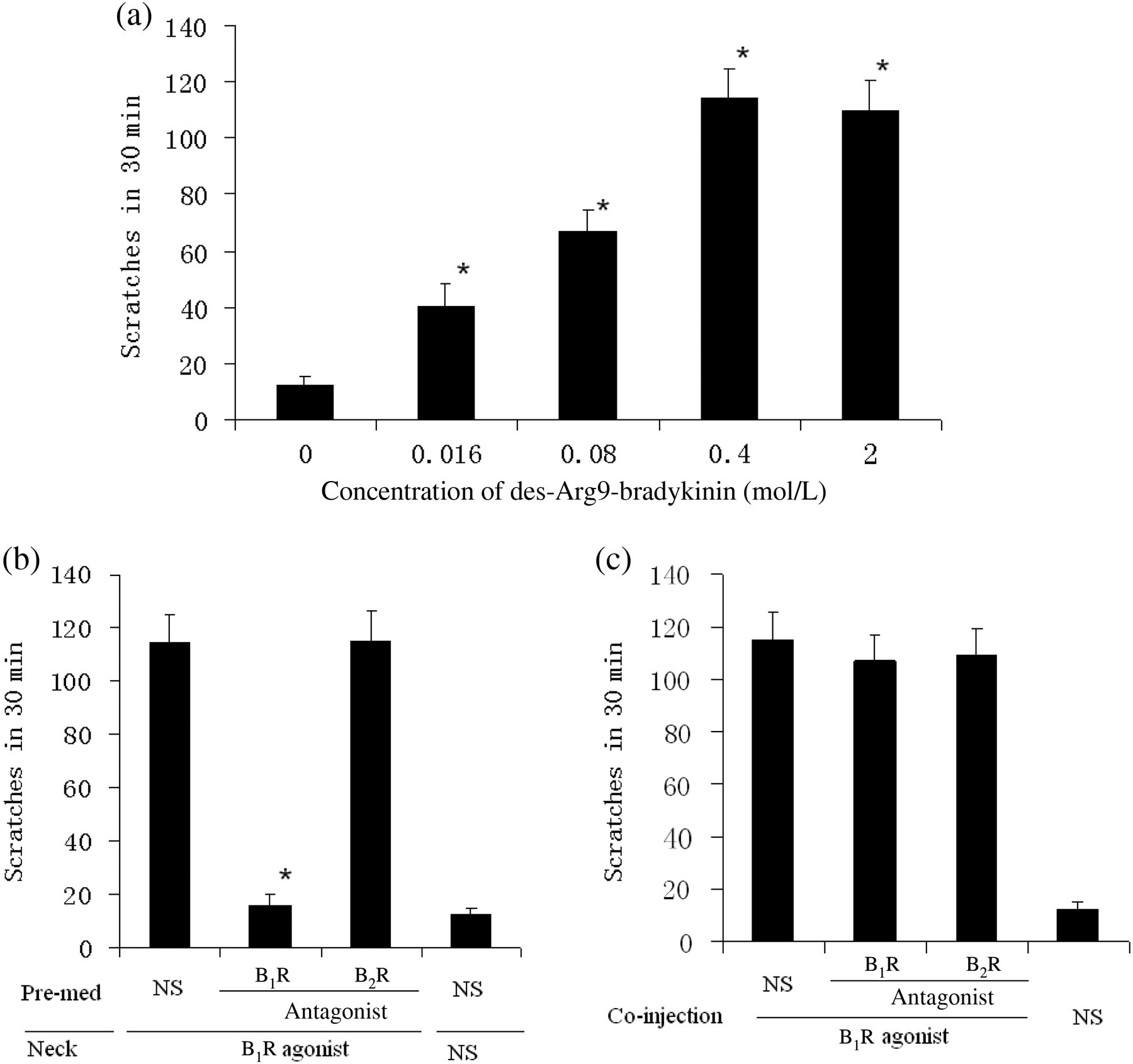
BK receptor modulates BK-induced pain-related behavior in CFA-inflamed mice
Finally, we examined the effects of BK receptor activation on BK-induced pain-related behavior in CFA-inflamed mice. Unilateral inflammation was induced in the left hind feet of mice by CFA injection, and mice were then given NS, BK (1 mmol/L) or B1R agonist (0.4 mmol/L) in the inflamed area four days later. Licking behavior was monitored immediately after injection for a period of five minutes. Paw licking time was increased in CFA-inflamed mice compared with non-inflamed mice (NS group) with each treatment (P < 0.05) (Figure 6a). However, pretreatment or co-injection with B2R antagonist dramatically reduced the elongated licking time induced by BK injection (P < 0.05 compared with BK treatment alone) (Figures 6b and c), while B1R antagonist treatment had no significant effect. This suggests that B2R plays a critical role in modulating BK-induced pain-related behavior in CFA-inflamed mice, while B1R does not.
BK receptor modulates BK-induced pain-related behavior in CFA-inflamed mice. (a) Four days after intradermal injection of 10 μL CFA (inflammation group) or NS (non-inflammation group) into the left hind feet, mice were given NS, B1R agonist (0.4 mmol/L) or BK (1 mmol/L) in a volume of 10 μL. Licking behavior was assayed immediately after injection for a period of five minutes. Data are expressed as mean ± SEM (n = 7 in each group). *P < 0.05 compared with the same treatment in the non-inflammation group. (b) Four days after CFA inflammation, mice were given an intraperitoneal injection of B1R or B2R antagonist (10 mg/kg body weight) 30 min prior to BK (1 mmol/L in 10 μL) or normal saline (10 μL) injection into the hindpaw. Licking behavior was assayed immediately after injection for a period of five minutes. Data are expressed as mean ± SEM (n = 7 in each group). *P < 0.05 compared with BK injection after intraperitoneal injection of normal saline (NS) in CFA-treated mice. (c) After four days of CFA inflammation, B1R, B2R antagonist (1 nmol) or NS were intradermally co-injected with BK (1 mmol/L) in a total volume of 10 μL into the hindpaw. Licking behavior was assayed immediately after injection for a period of five minutes. Data are expressed as mean ± SEM (n = 7 in each group). *P < 0.05 compared with co-injection of solvent and BK in CFA-treated mice. CFA, complete Freund's adjuvant; BK, bradykinin; NS, normal saline
BK-induced scratching and wiping behavior with cheek injection
Local inflammation was induced in mice by a 10 μL intradermal injection of CFA into the left cheek. As shown in Figure 1, CFA-induced scratching and wiping activity occurred immediately after injection (P < 0.05 compared with the non-inflammation group; Figure 7). We next monitored the B1R agonist- and BK-evoked scratching and wiping behavior in mice after four days of CFA inflammation. Scratching activity was observed when B1R agonist or BK was injected intradermally into the cheek (P < 0.05 compared with the non-inflammation group; Figure 8a). Wiping activity was increased in all animals in the inflammation group when compared with the non-inflammation group (Figure 8b). B1R agonist evoked more scratches than BK; on the contrary, less wipes were evoked by B1R agonist when compared with BK (Figure 8).
CFA-induced scratching and wiping behavior. Mice received an intradermal injection (10 μL) of CFA (inflammation group) or NS (non-inflammation group) into the left cheek. Data are expressed as mean ± SEM (n = 8 in each group). (a) The hind limb scratching behavior directed toward the injected area was recorded for a period of 30 min after injection of CFA or NS. *P < 0.05 compared with the NS group. (b) The left forelimb wiping behavior directed toward the injected area was recorded for a period of 30 min after injection of CFA or NS. *P < 0.05 compared with the NS group. CFA, complete Freund's adjuvant; NS, normal saline BK- and B1R agonist-induced scratching and wiping behavior. Four days after intradermal injection of 10 μL CFA (inflammation group) or NS (non-inflammation group) into the left cheek, mice were given NS, B1R agonist (0.4 mmol/L) or BK (1 mmol/L) in a volume of 10 μL. Data are expressed as mean ± SEM (n = 8 in each group). (a) The hind limb scratching behavior directed toward the injected area was recorded for a period of 30 min after injection. *P < 0.05 compared with the non-inflammation group. (b) The left forelimb wiping behavior directed toward the injected area was recorded for a period of 30 min after injection of CFA or NS. *P < 0.05 compared with the non-inflammation group. CFA, complete Freund's adjuvant; BK, bradykinin; NS, normal saline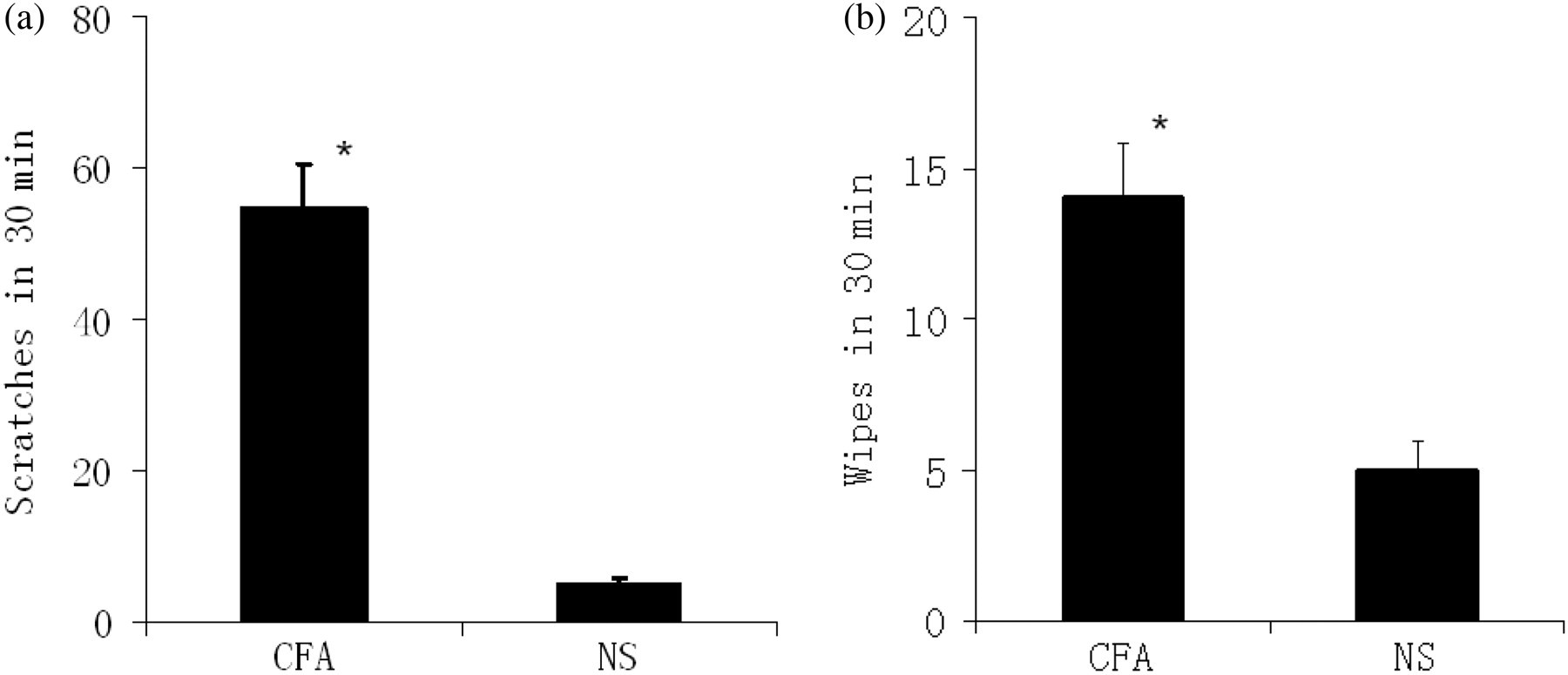
Histological findings
Diffuse infiltration of inflammatory cells was observed in the subcutaneous area in the CFA-inflammation mice. Scattered necrosis was also found in the subcutaneous area (Figure 9).
Histological features of CFA-inflammation skin. Mouse skin of the back of the neck was harvested on day 4 after the induction of inflammation. Tissue was fixed and stained with hematoxylin—eosin. (a) Section of skin treated with NS (n = 4). (b) Section of skin treated with CFA (n = 4). Diffuse infiltration of inflammatory cells was observed in the subcutaneous area in the CFA-inflammation mice. Scattered necrosis was found in the subcutaneous area. Arrows indicate the area of necrosis. Original magnification ×100. CFA, complete Freund's adjuvant; NS, normal saline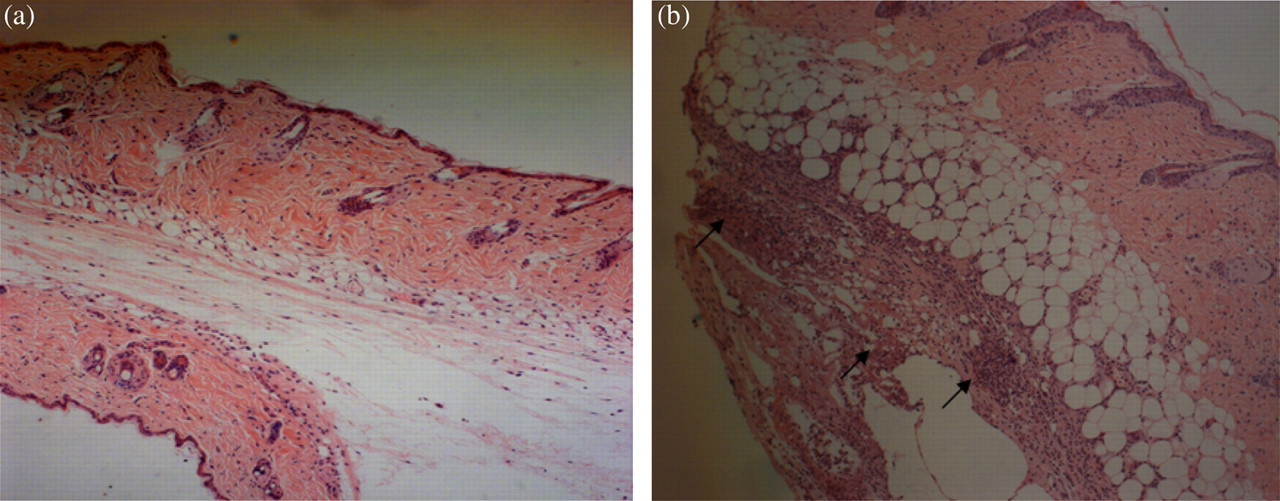
Discussion
Peripheral administration of CFA has been widely used to evoke an inflammatory response in mice and rats. 20–22 For instance, acute inflammation after unilateral injection of CFA into the plantar surface of the rat's hindpaw causes intense edema and hyperalgesia within a few hours. 23 Samad et al. 24 demonstrated that this effect could be mediated by several pathways, including local nociceptor sensitization; systemic neuronal mechanisms, such as central sensitization; and immune responses, such as increased cytokine serum levels. In the present study, local inflammation was induced by intradermal injection of CFA into the back of the neck, the hind foot or the cheek of the mouse. CFA-induced scratching activity occurred immediately after injection to the back or the cheek. More scratches than wipes were evoked by the cheek injection of CFA. It is interesting that itchness rather than pain was evoked by CFA at the early time points. Many inflammatory mediators and pathways were found to be involved in the process of CFA-induced inflammation. The early inflammation induced by CFA has been shown to be mast cell–dependent. 25 The mechanism of itch induced by CFA in the acute phase should be investigated further. We found that BK dose-dependently evoked scratching responses in the CFA-inflamed area. However, whether or not scratching behavior is a result of itch, pain or some other sensation is difficult to determine in mice. A previous study confirmed that scratching behavior induced by intradermal injection of pruritogenic agents into the back of the mouse neck was itch-related. 26 In addition, we have verified that itchiness was induced by BK in inflamed skin by injecting into the cheek. Our results showed that B2R antagonist enhanced the scratching activity induced by BK while reducing pain-associated licking behavior, suggesting the scratching behavior detected here is in fact an itch-related activity.
Itch, a common skin sensation, is an essential symptom of skin diseases and is usually linked to the desire to scratch. 1 It can be generated by a variety of stimuli, including inflammatory infections. It was reported that a majority of children accidentally injected with the Bacille Calmette-Guérin vaccine will have an itchy, painful skin reaction at the injection site. 27 An unexpectedly high incidence of persistent itching nodules and delayed hypersensitivity to aluminium in children were also reported after the use of adsorbed vaccines from a single manufacturer. 28 In the present study, we established a novel mouse model of itch-related scratching behavior evoked by BK, a pain-producing substance in CFA-inflamed skin. In patients with atopic dermatitis, a spontaneously occurring and frequent skin disease, pain can be induced independently of itch in lesional skin. 7,29 Disease models for atopic dermatitis in rodents include hapten-induced dermatitis in mice, NC/Nga mice and genetically engineered mouse models. 30 However, in this current study, B2R antagonist greatly enhanced the scratching behavior evoked by BK in inflamed mice, implying that BK might stimulate pain in the inflamed area, and that this pain possibly suppressed the itch sensation. However, the current rodent model is different from atopic dermatitis, in which the pain stimuli are perceived as itch but do not suppress the itch sensation.
Our results demonstrate that BK can induce scratching responses in CFA-inflamed mouse skin via activation of B1R. Additionally, B1 and B2 receptors appear to play different roles in modulating BK-induced itch-related behavior in CFA-inflamed mice. BK has been linked to a wide range of biological phenomena such as pain, inflammation and itch, 6,31 and exerts its effects through the activation of B1 and B2 receptors. 6 B1R activation has been shown to contribute to inflammatory pain and hyperalgesia. 12,32 Recently, B1R and B2R have been shown to play a crucial role in controlling the pruriceptive signaling triggered by proteinase-activated receptor 2 (PAR2) activation in mice. 33 In this study, we find that pretreatment with B1R antagonist dramatically reduced BK-induced scratching behavior, whereas B2R antagonist pretreatment promoted the scratching activity induced by BK. Furthermore, pretreatment with B1R antagonist remarkably suppressed the scratching behavior induced by B1R agonist. These findings strongly suggest that both BK receptors are involved in and contribute to BK-induced scratching behavior in CFA-inflamed mice, but their effects depend on the treatment paradigm. However, B1R and B2R clearly play opposite roles in this process. Systemic, but not local injection of B1R antagonist DALBK reduced scratches evoked by BK or selective B1R agonist. Co-injection of DALBK was also not able to reduce scratches evoked by SLIGRL-NH2, 33 while peripheral injection of DALBK abolished the nociceptive behavior induced by intraplantar injection of PMA. 17 These studies, together with our results, suggest that peripheral B1R was less likely to be involved in this scratching response than was a lack of potent B1R antagonism, as the dose of DALBK used in this study was larger than that used previously. 17
Abnormal expression or activity of serine proteases and PAR2 has been associated with several inflammatory skin disorders involving barrier abnormalities, including atopic dermatitis, Netherton syndrome and psoriasis. 34 The endogenous PAR-2 agonist tryptase was increased in atopic dermatitis patients, 35 and PAR2 agonists induce pruritus in patients and animals. 16,33,35 Both B1R and B2R antagonists were found to be effective in reducing the scratch response induced by PAR2 activation in mice. 33 B2R is constitutively expressed in many cell types and in most tissues, and modulates physiological functions under normal conditions. 36 In contrast, B1R is almost absent in normal tissues and is usually induced during inflammation. 37 Upregulated B1R expression has been observed following inflammation induced by a number of stimuli. 17,38,39 It will be important to explore the role of B1R in pathophysiological inflammatory itch conditions, as B1R antagonists might be effective antipruritic drugs for some chronic itchiness.
With regard to BK-induced pain-related behavior in CFA-inflamed mice, pretreatment or co-injection with B2R antagonist dramatically reduced the elongated licking time induced by BK injection, while no obvious effect was observed upon B1R antagonist treatment, suggesting that B2R plays a critical role in modulating BK-induced pain-related behavior in CFA-inflamed mice. These observations were consistent with previous reports showing that CFA inflammation increased sensitivity of C-fiber nociceptors to BK through B2R, and B2R antagonism significantly inhibited CFA-induced ipsilateral mechanical hyperalgesia in mice. 21,40 However, the opposite result has been obtained by other research groups, which have shown that B1R, but not B2R, is involved in inflammatory pain. For instance, accumulating evidence suggests that upregulation of B1R protein levels contributes to inflammatory pain and hyperalgesia. 12,32 In addition, B1R antagonist inhibited carrageenin-induced inflammatory hypernociception, as well as tumor necrosis factor alpha (TNF-α) and interleukin 1 beta (IL-1β) release in lipopolysaccharide-primed mice, suggesting that B1R mediates hypernociception through a mechanism dependent on TNF-α and IL-1β. 12 B2R antagonist had no such effect. Furthermore, p38 stress-activated protein kinase has been demonstrated to contribute to the development of inflammatory hyperalgesia in a rat model by promoting B1R activity. 12 The discrepancy between these results may be due to differences in the applied experimental paradigms. For instance, local injection of B1R or B2R antagonist could inhibit spontaneous pain by reducing licking time in mice inoculated with melanoma, whereas only B2R antagonist attenuated allodynia in mice with a melanoma-bearing paw. 41 Therefore, it is possible that BK receptors can act through different mediators or in concert, depending on the context. The stimulation of B1R also contributes to the acute inflammatory pain process, as B1R antagonist or B1R gene deletion can lead to the abolishment of PMA-induced nociceptive behavior. 17 Several signaling mechanisms related to PMA-evoked nociception act via protein kinase C. 42 On the other hand, acute and chronic inflammatory pain models may lead to different results.
In summary, this study demonstrated that BK induces itch-related response in CFA-inflamed mouse skin via activation of B1R, while activation of B2R inhibits the itch-related response.
Footnotes
ACKNOWLEDGEMENTS
The authors would like thank Dr Ping Mei for technical assistance. This study was supported by the National Natural Science Foundation (Nos. 30872437 and 81171040) and Natural Science Foundation of Guangdong Province, China (No. 10151008002000005).