
Abstract
Embryo implantation is a process that requires both temporal and spatial synchronization of the uterine endometrium and the embryo, and the endometrium becomes receptive to the embryo during the window of implantation. Although the expression patterns of many implantation-related molecules change dynamically during this process, the impact of CD82 on endometrial receptivity has not been elucidated. By immunohistochemical staining, we found that CD82 levels rose from the proliferative phase to the secretory phase in human endometrium. Specifically, the highest level appeared in mid- and late-secretory phases. Consistently, RL95-2 cells, representative of high-receptive endometrial epithelium, expressed higher levels of CD82 than did HEC-1A cells, which are representative of low-receptive endometrial epithelium, as detected by reverse transcription-polymerase chain reaction, Western blot and immunofluorescence. Furthermore, progesterone up-regulated the expression of CD82 in both epithelial cell lines. Down-regulation of CD82 in RL95-2 cells by either CD82 siRNA transfection or treatment with a CD82 antibody significantly decreased the adhesion of human embryonic JAR cells to RL95-2 cell monolayers (P < 0.01) and inhibited the phosphorylation of focal adhesion kinase (FAK). In contrast, up-regulation of CD82 in HEC-1A cells by CD82 cDNA transfection promoted embryonic JAR cell adhesion to HEC-1A monolayers (P < 0.05) and activated the phosphorylation of FAK. In conclusion, the expression of CD82 increases in endometrial tissues during the window of embryo implantation, CD82 expression affects endometrial receptivity of the uterine epithelial cells in vitro, and the FAK signaling pathway may be involved in this phenomenon. The correlation between CD82 and endometrial receptivity suggests that CD82 may serve as a potential marker of endometrial function.
Introduction
Embryo implantation is a key process in human reproduction that is divided into three steps: apposition, adhesion and invasion. 1 First, the embryo locates to the receptive endometrium, and adheres to uterine epithelial cells. Then, trophectoderm cells penetrate the endometrial surface. 2 During this process, the endometrium undergoes a concurrent transition from a non-receptive state to a receptive state, with clear morphological changes, as well as increased expression of adhesion molecules, on the surface of the endometrium, which regulate the process of embryo implantation. 3
Glycoproteins, found on cell surfaces, are involved in the adhesion of cells to other cells and adhesion of cells to the extracellular matrix. 4 CD82 is a glycoprotein that belongs to the transmembrane 4 superfamily, 5 and it has important roles in a variety of processes including cancer metastasis, immune response and cellular morphogenesis. 6–8 CD82 levels are reduced significantly in tumors of the endometrium, colon, breast, bladder and cervix. 9,10 Gellersen et al. 11 found that trophoblast cells do not produce CD82, but decidual cells at the maternal–fetal interface express a significant amount of CD82, indicating that CD82 may participate in embryo implantation. One of the key steps of embryo implantation is the adhesion of the embryo to the uterine epithelial cells, and to date, whether CD82 is involved in this adhesion process is largely unknown.
The endometrial cycle is regulated by estrogen and progesterone. 12 During the window of implantation (days 20–24), the endometrium is stimulated by progesterone. 13,14 Indeed, progesterone plays critical roles for the embryo implantation and is required for the maintenance of pregnancy. 15 Low progesterone levels can cause infertility in women, 16 and progesterone stimulates the uterine endometrium to enter a temporally limited receptive state by regulating various implantation-related molecules. 17
Genomic sequencing shows that the genes for the progesterone receptor and CD82 are on the same chromosome. 11 However, it is unclear whether progesterone is an upstream regulatory factor of CD82. In this study, we examined the expression of CD82 in both human endometrial samples and in vitro implantation models, using RL95-2 and HEC-1A human endometrial cell lines and the JAR human embryonic cell line. 18–20 Of note, RL95-2 and HEC-1A cell lines represent the high- and low-receptive endometrial epithelium, respectively. Our results demonstrate that CD82 expression levels are related to endometrial receptivity, and CD82 expression can be up-regulated by progesterone. In addition, our results suggest that CD82-regulated adhesion between the embryo and the endometrium may be related to the focal adhesion kinase (FAK) signaling pathway.
Materials and methods
Antibodies and reagents
RNA PCR Kit (AMV) was purchased from the Takara Company (Dalian, Liaoning, China). Dulbecco's Modified Eagle Medium/F12 (1:1), McCoy's 5A, RPMI 1640, fetal bovine serum (FBS), TRIzol® and Lipofectamine™ reagent were purchased from Invitrogen (Camarillo, CA, USA). The enhanced chemiluminescence (ECL) assay kit was purchased from Amersham (Pittsburgh, PA, USA). Progesterone and diaminobenzidine (DAB) were purchased from Sigma (St Louis, MO, USA). Mouse anti-human CD82 and horseradish peroxidase (HRP)-conjugated anti-rabbit or anti-mouse secondary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Rabbit anti-human glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibodies were purchased from Abcam (Cambridge, CA, USA). Mouse anti-human FAK antibodies were purchased from BD Biosciences (San Jose, CA, USA). Rabbit anti-human phosphorylated FAK antibodies were purchased from Biosource (Carlsbad, CA, USA). Mouse anti-human phosphorylated Tyr antibodies were purchased from Transduction Laboratories (San Jose, CA, USA). TRITC-conjugated goat anti-mouse IgG and FITC-conjugated goat anti-mouse IgG were purchased from Santa Cruz Biotechnology. Coomassie protein assay reagent was purchased from Bio-Rad (Hercules, CA, USA).
Tissue collection
The protocol for human study was approved by the Institutional Review Board of Dalian Medical University. All human specimens used in this study were collected from patients between the ages of 30 and 45. With the patients’ consent, endometrial blocks were obtained from 40 hysterectomy specimens (including eight specimens each from early proliferative phase, late-proliferative phase, early secretory phase, mid-secretory phase and late-secretory phase). These paraffin-embedded human endometrial tissues were selected after histological review.
Cell culture
Human uterine epithelial cell lines (RL95-2 and HEC-1A) and human embryonic cell line (JAR) were acquired from the American Type Culture Collection (Manassas, VA, USA). RL95-2 cells were grown in DMEM/F12 (1:1) supplemented with 10% FBS, 5 μg/mL insulin, 100 U/mL penicillin and 100 μg/mL streptomycin. HEC-1A cells were grown in McCoy's 5A supplemented with 10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin. JAR cells were maintained in RPMI 1640 supplemented with 10% FBS. All types of cells were maintained at 37°C under 5% CO2 in humidified air. Cells (1×106) were seeded onto six-well plates. When the cells reached 80% confluence, cells were washed three times with phosphate-buffered saline (PBS) and subsequently were starved of serum for three hours before progesterone treatment. The concentrations of progesterone used were 10−5, 10−6 and 10−7 mol/L. Cells grown in serum-free medium were used as a control. Cells were harvested for RNA at 48 h and for protein at 72 h. Each experiment was repeated three times.
Immunohistochemistry
Serial sections (4–6 μm each) were prepared from paraffin-embedded tissues. The sections were fixed at 60°C for one hour, deparaffinized in xylene and rehydrated in graded alcohol. For the detection of CD82, the slides were microwaved for 20 min in citrate buffer in order to unmask antigen and were washed with PBS after cooling for 20 min. Slides were then incubated in 3% H2O2 in methanol for 10 min to block endogenous peroxidase activity. After washing in PBS, sections were blocked with blocking buffer supplemented with normal goat serum at room temperature for 10 min to eliminate non-specific binding of conjugated secondary antibodies before incubation overnight at 4°C with 0.5 μg/mL mouse anti-human CD82. After several rinses with PBS, sections were incubated with secondary antibody, a biotinylated goat anti-mouse immunoglobulin, for 15 min at 37°C. The signal was visualized with peroxidase-labeled streptavidin-complexed DAB, and the sections were briefly counterstained with hematoxylin. Yellowish-brown stain indicated a positive result. The negative control was generated by replacing the primary antibody with isotype IgG. Slides were mounted and visualized at ×10 and ×40 magnifications on an inverted microscope (Olympus, Tokyo, Japan). Histological and immunohistochemical evaluations were performed independently by two pathologists. Immunostaining intensity was evaluated in each endometrial compartment (lumina epithelium and glandular epithelium) using a semi-quantitative method. Each sample was given a score in which both the intensity of the staining (none staining = 0; low staining = 1; medium staining = 2; strong staining = 3) and the percentage of stained epithelial cells were multiplied. In normal endometrium, the total score was calculated per compartment per sample, as follows: H score = ∑Pi (i + 1), where i is the intensity of staining from 0 (none) to 3 (strong), and Pi is the percentage of stained cells for each given i (0%–100%). In addition, the immunohistochemical localization pattern was recorded in digital images.
Transient transfection
Cells were seeded onto six-well plates. When cells reached 80% confluence, CD82 siRNA or CD82 cDNA was transiently transfected into the cells using 400 ng of plasmid and 4 μL of Lipofectamine™ reagent, following the manufacturer's instructions. The transfection was terminated six hours later and cells were harvested for RNA at 48 h and for protein at 72 h. Each experiment was repeated three times.
Reverse transcription-polymerase chain reaction
Total RNA was extracted with the TRIzol® reagent according to the manufacturer's protocol. Semi-quantifications and purity assessments were performed by optical density measurement at 260 and 280 nm. Total RNA was reverse transcribed into cDNA using RNA PCR Kit (AMV) version 3.0. The CD82 primers (polymerase chain reaction [PCR] products 276 bp) were 5′ TCAGCCTGTATCAAAGTCACC 3′ (sense) and 5′ ATCAGGAGCAGGAAAGCAAAG 3′ (antisense). The GAPDH primers (PCR products 288 bp) were 5′ GGAGTCCACTGGCGTCTTCAC 3′ (sense) and 5′ GAGGGGCCATCCACAGTCTTCT 3′ (antisense). For each reaction, 100 ng cDNA was added into a PCR master mixture to the total volume of 25 μL. PCR reactions were performed as follows: initial denaturation at 94°C for five minutes, 28 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 30 s and a final extension for 72°C for 10 min. The PCR reaction products were electrophoresed on 1% agarose gel, and the bands were visualized by ethidium bromide staining, followed by analysis with Labworks 4.6 (UVP, Upland, CA, USA).
Western blot
Cells were washed with PBS (pH 7.4), and incubated with lysis buffer (1% Triton X-100, 150 mmol/L NaCl, 10 mmol/L Tris, pH 7.4, 1 mmol/L EDTA, 1 mmol/L EGTA, pH 8.0, 0.2 mmol/L Na3VO4, 0.2 mmol/L phenylmethylsulfonyl fluoride, 0.5% Nonidet P-40) for 15 min on ice. Cell lysates were clarified by centrifugation at 10,000 ×
Indirect immunofluorescence staining
After washing with PBS, cells grown on coverslips were fixed in 4% paraformaldehyde following 20 min. After being blocked with 3% complete serum for two hours at 37°C, cells were incubated with mouse anti-CD82 antibody (1:100) at 4°C overnight. The cells were then incubated with TRITC-conjugated goat anti-mouse IgG (1:100) or FITC-conjugated goat anti-mouse IgG (1:100) for 40 min. Images were captured with the Olympus BX51 fluorescence microscope.
Adhesion of embryonic cells JAR to RL95-2 cells and HEC-1 monolayer
The percentage adhesion of JAR to the endometrial cells was calculated as described.
21
In brief, RL95-2 or HEC-1A cells were grown on coverslips to form a confluent monolayer. Cultured JAR cells were harvested, counted and gently delivered onto RL95-2 or HEC-1A cell monolayers in JAR medium. One hour later, unbound JAR cells were removed by centrifuging the coverslips (cell surface facing down) at 12 ×
Statistical analysis
Each experiment was repeated 3–6 times, with results presented as the mean ± SEM. Statistical differences between test groups were analysed by one-way analysis of variance and Student-Newman-Keuls q value tests; P < 0.05 was considered to be significant and P < 0.01 was considered to be highly significant.
Results
Expression of CD82 in human endometrial tissues
Immunohistochemistry was used to determine whether CD82 levels change cyclically in the human endometria during the menstrual cycle (Figure 1). Results show that in the early proliferative phase, CD82 was minimally expressed (Figure 1a). In the late-proliferative stage, CD82 was expressed weakly in part of the luminal and glandular epithelia (Figure 1b). However, in the secretory phase, CD82 levels were significantly higher (P < 0.05). In the early secretory phase, CD82 was expressed in almost all of the luminal and glandular epithelia (Figure 1c), and the highest expression of CD82 occurred in the mid- and late-secretory phases (Table 1). Moreover, CD82 was observed in stromal cells in the mid- and late-secretory phases, although the level was much weaker than that in the epithelia (Figures 1f and g).
CD82 expression in the human endometrium. Paraffin-embedded endometrial tissues at various points of the menstrual cycle were analyzed by immunohistochemistry. CD82 expression in (a) early proliferative, (b) late-proliferative and (c) early secretory phases. (d) Hematoxylin–eosin stain in mid-secretory phase. (e) Negative control in mid-secretory phase (isotype IgG). (f) CD82 expression in mid-secretory phase. (g) CD82 expression in late-secretory phase. LE, lumina epithelium. GE, glandular epithelium. Panels are magnified ×100 and insets are magnified ×400. Bar = 1 mm. (A color version of this figure is available in the online journal) Expression of CD82 protein image analysis in human endometrial epithelium of different phases EP, early proliferative phase; LP, late-proliferative phase; ES, early secretory phase; MS, mid-secretory phase; LE, late-secretory phase *Results were expressed as the mean ± SEM of H-score. Eight specimens each from different phases. a,b,c,d,e: Significant difference for EP, LP, ES, MS and LS groups, respectively (P < 0.05)
Expression of CD82 in RL95-2 and HEC-1A cells
To assess whether the expression of CD82 correlates with endometrial epithelium receptivity, CD82 expression was examined in RL95-2 and HEC-1A cells by reverse transcription-PCR (RT-PCR) (Figure 2a), Western blot (Figure 2b) and indirect immunofluorescence staining (Figure 2c). Results show that both CD82 mRNA and protein were more highly expressed in high-receptive RL95-2 cells than in low-receptive HEC-1A cells (P < 0.05).
Expression of CD82 in RL95-2 and HEC-1A cells. (A) RT-PCR analysis of CD82 mRNA in RL95-2 and HEC-1A cells. (B) Western blot detection of CD82 protein in RL95-2 and HEC-1A cells. (C) Expression of CD82 protein is detected by indirect immunofluorescence staining in (a) RL95-2 and (c) HEC-1A cells. Control (b) RL95-2 and (d) HEC-1A cells as seen under a light microscope (*P < 0.05, **P < 0.01). All images are magnified ×400. Bar = 15 μm. RT-PCR, reverse transcription-polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. (A color version of this figure is available in the online journal)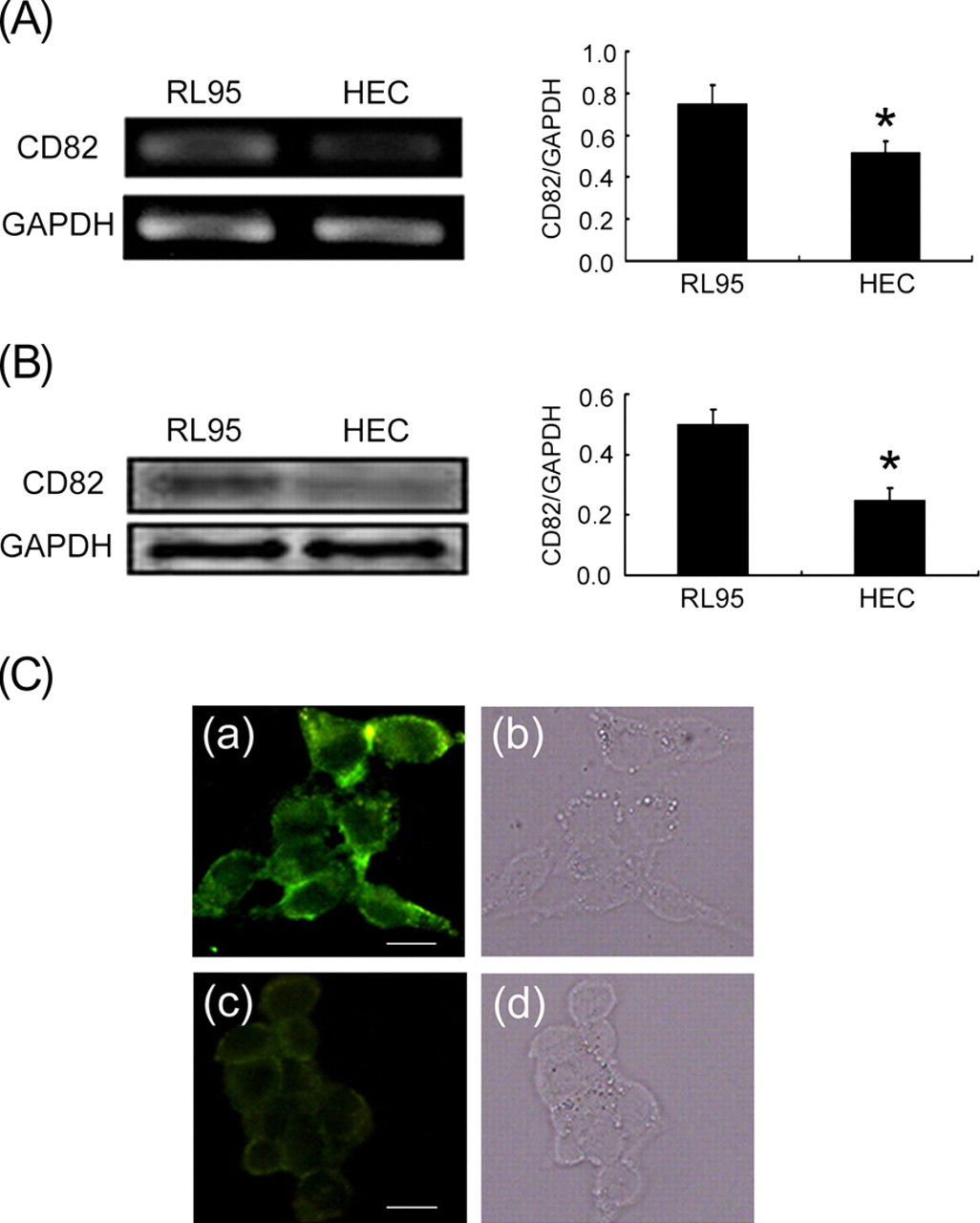
The effect of progesterone on CD82 in RL95-2 and HEC-1A cells
RL95-2 and HEC-1A cell lines were both treated with progesterone as described. RT-PCR and Western blot were used to detect the effect of progesterone treatment on CD82 expression (Figure 3). Results show that CD82 expression at both gene and protein levels was stimulated by progesterone in a dose-dependent manner (P < 0.01).
Effect of progesterone on CD82 expression in RL95-2 and HEC-1A cells. Cells were treated with different concentrations of progesterone. (A) RT-PCR analysis of CD82 mRNA in RL95-2 cells. (B) Western blot detection of CD82 protein in RL95-2 cells. (C) Expression of CD82 mRNA, as measured by RT-PCR, in HEC-1A cells. (D) Expression of CD82 protein, as observed by Western blot, in HEC-1A cells. (*P < 0.05, **P < 0.01). RT-PCR, reverse transcription-polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase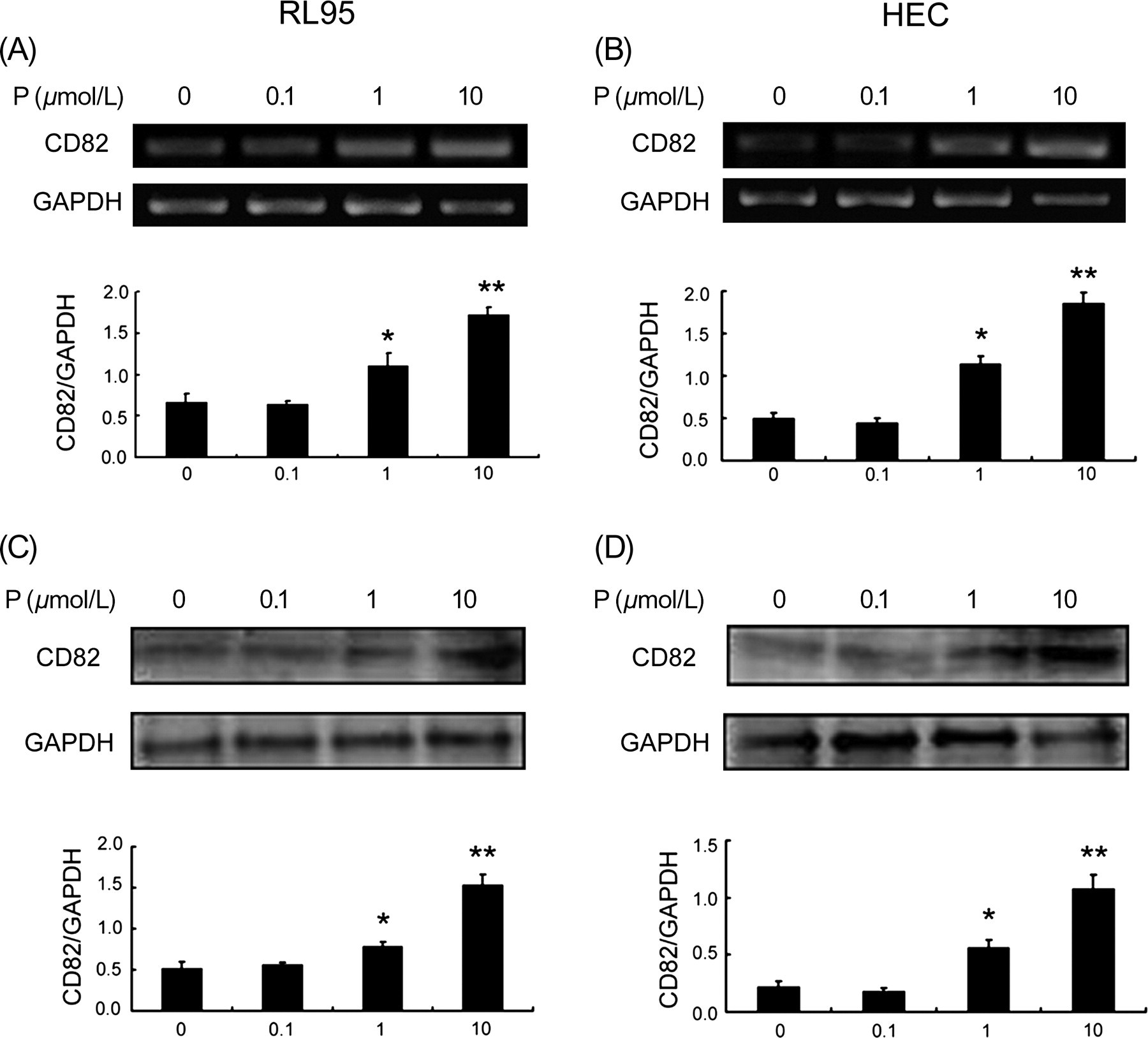
CD82 plasmid transfection changes the expression of CD82 in RL95-2 and HEC-1A cells
CD82 mRNA expression in CD82 siRNA-transfected RL95-2 cells was dramatically lower than in control RL95-2 cells, as determined by RT-PCR (Figure 4a). Western blot (Figure 4b) and indirect immunofluorescence staining (Figure 4c) showed that the expression of CD82 protein was also much lower in siRNA-transfected cells than in control cells. These results show that CD82 siRNA effectively suppresses the expression of CD82 in RL-95 cells (P < 0.01). In comparison, in CD82 cDNA-transfected HEC-1A cells, the expression of CD82 was much higher than the control group (Figures 4d–f). The results show that CD82 cDNA strongly increases the expression of CD82 in HEC-1A cells (P < 0.01).
Regulation of CD82 expression by transfection of CD82 plamids in RL95-2 and HEC-1A cells. (CD82 GenBank: NM_002231) (A) RT-PCR of CD82 expression in RL95-2 cells after transfection with CD82 siRNA, (B) Western blot and (C) indirect immunofluorescence staining with (a, b, c) CD82 primary antibody and FITC-conjugated anti-mouse IgG secondary antibody or (d) FITC-conjugated anti-mouse IgG secondary antibody alone. (a, d) Untransfected cells; (b) cells transfected with vector; and (c) cells transfected with CD82 siRNA. Expression of CD82 in HEC-1A cells after transfection with CD82 cDNA (D) observed by RT-PCR, (E) Western blot and (F) indirect immunofluorescence staining with (a, b, c) CD82 primary antibody and TRITC-conjugated anti-mouse IgG secondary antibody or (d) TRITC-conjugated anti-mouse IgG secondary antibody alone. (a,d) Untransfected cells; (b) cells transfected with vector; and (c) cells transfected with CD82 cDNA. (*P < 0.05, **P < 0.01). Bar = 15 μm. RT-PCR, reverse transcription-polymerase chain reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. (A color version of this figure is available in the online journal)
CD82 influences the adhesion of embryonic cells JAR to both RL95-2 and HEC-1A monolayers
To further study the function of CD82 in embryo adhesion to the uterine epithelium, both RL95-2 cells and HEC-1A cells were used (Figure 5a). The adhesion percentage was calculated and is shown in Figure 5b. Vector-transfected RL95-2 cells did not significantly elevate embryo adhesion (74.36 ± 1.7%) compared with the non-transfected control cells (72.54 ± 2.1%) (P > 0.05). However, embryo adhesion to RL95-2 cells was significantly decreased when these cells were transfected with 50 nmol/L CD82 siRNA (64.83 ± 2.0%), and even more with 200 nmol/L CD82 siRNA (46.17 ± 1.5%), when compared with the vector-transfected cells (P < 0.01). Similarly, blocking CD82 in RL95-2 cells with CD82 antibody also significantly decreased the percent adhesion of embryos (38.28 ± 1.4%) when compared with the non-transfected control (P < 0.01).
Adhesion analysis of JAR embryonic cells to RL95-2/HEC-1A uterine epithelial cells. (A) Adhesion of JAR cells to RL95-2/HEC-1A cells. (a) RL95-2 cell monolayer; (b) a representative image of JAR cells attached to RL95-2 cells; (c) HEC-1A cell monolayer and (d) a representative image of JAR cells attached to HEC-1A cells. R, RL95-2; H, HEC-1A; J, JAR. (B) Percentage adhesion was calculated as stated in the Materials and methods section. (*P < 0.05, **P < 0.01). Bar = 15 μm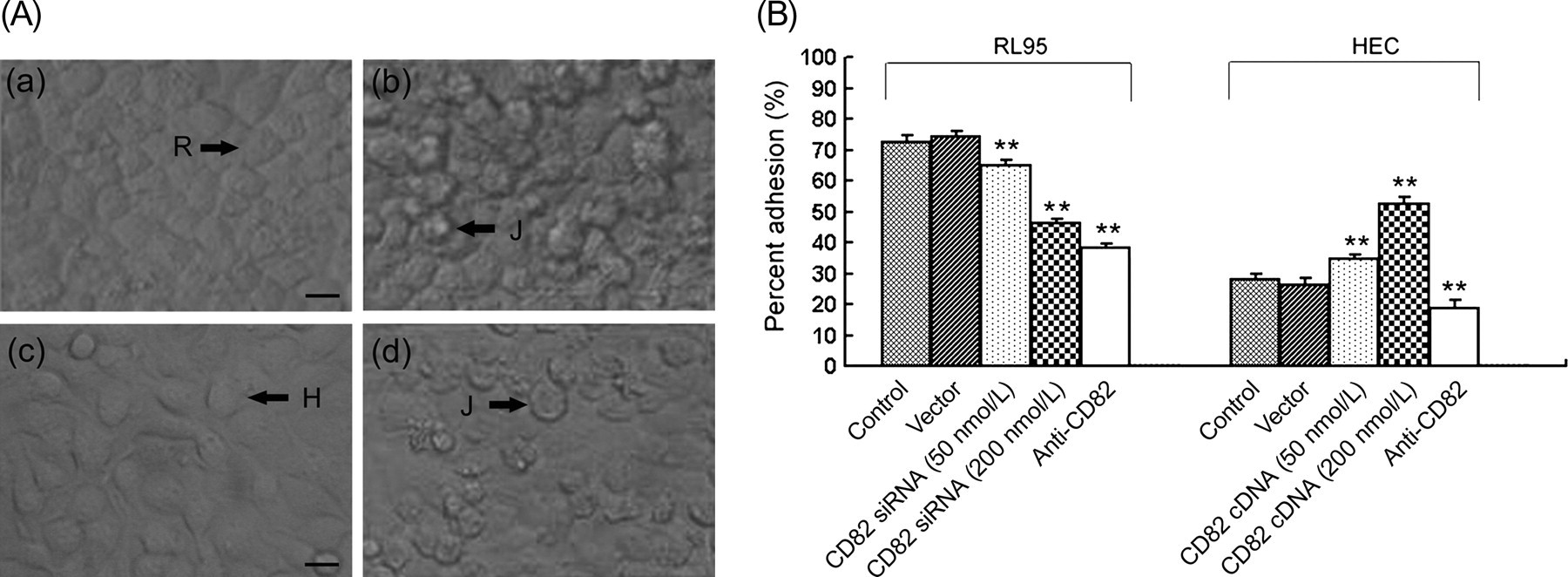
In HEC-1A cells, no significant differences were seen in the percentage adhesion between non-transfected cells (28.16 ± 1.8%) and those transfected with a control vector (26.43 ± 2.2%) (P > 0.05). In contrast, the percentage adhesion of embryos in HEC-1A cells transfected with 50 or 200 nmol/L CD82 cDNA was significantly increased when compared with vector-transfected cells (P < 0.01). When HEC-1A cells were preincubated with the CD82 antibody, the percentage adhesion was also decreased (18.83 ± 2.4%) compared with the non-transfected control (P < 0.01).
The expression of CD82 correlates with the FAK signaling pathway
Western blots were used to study the effect of CD82 on the FAK signaling pathway. In RL95-2 cells, CD82 siRNA transfection decreased expression of both p-FAK and p-Tyr (Figure 6a), but up-regulation of CD82 by cDNA transfection significantly increased p-Tyr and p-FAK levels in HEC-1A cells (Figure 6b). When RL95-2 and HEC-1A cells were blocked with CD82 antibody, the levels of p-Tyr and p-FAK were both inhibited.
Effects of CD82 on FAK signaling pathway. Total protein was isolated from the whole cell lysate after stimulation with JAR cells for 30 min. Total protein from specific cell lines was prepared and separated using the 12% SDS-PAGE mini-gel, and transferred onto nitrocellulose membrane as indicated in ‘Materials and methods’. FAK, phosphorylated FAK (p-FAK) and tyrosine kinase were detected with antibodies directed against FAK (top row), p-FAK (second row) and p-Tyr (third row) from RL95-2 (left) cells and HEC-1A cells (right) by Western blot. SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; FAK, focal adhesion kinase
Discussion
During the window of implantation, endometrial epithelial cells undergo ultrastructural changes and express a variety of molecules to facilitate the adhesion of the embryo to the uterine endometrium. 22 Adhesion-related molecules such as mucins and integrins increase in the window of implantation, and are known to participate in the establishment of endometrial receptivity. 23,24 For example, the expression of integrin αvβ3 is significantly increased throughout the mid-secretory and late-secretory phases, 25 and has been recognized as a marker of the implantation window. 26,27 CD82 is ubiquitously expressed in normal tissues. 6 Alterations in CD82 are associated with different pathological and physiological phenotypes in human. For example, reduced CD82 correlates with enhanced malignant metastatic phenotypes in tumors of endometrium, prostate, colon, lung, pancreatic, breast and ovary, 9,28 and elevated CD82 is related to augmented cell adhesion. 29 In addition, interaction of CD82 on tumor cells with DARC (Duffy antigen/receptor for chemokines) on vascular endothelium suppresses metastasis, 30 which suggests that CD82 may act as an adhesive molecule at the fetal–maternal interface. In this study, we found that CD82 expression was significantly increased in the mid- and late-secretory phases of the endometrial cycle. Furthermore, high-receptive RL95-2 cells expressed higher levels of CD82 than did low-receptive HEC-1A cells. Altogether, these results suggest that CD82 may be necessary for the establishment of endometrial receptivity and its expression may serve as a potential marker in the evaluation of endometrial receptivity.
As the endometrium switches from the proliferative to the secretory phase, progesterone is increased consistently, and reaches its peak in the mid-secretory phase. Increased progesterone facilitates embryo implantation in humans. 13,15 Progesterone is an upstream regulatory factor for many molecules related with implantation, 31 such as osteopontin, integrin, insulin-like growth factor, vascular endothelial growth factor and prostaglandin E2 in the human endometrial epithelium. 32–35 By immunohistochemistry staining, we found that the highest expression of CD82 occurred in the mid- and late-secretory phases. Moreover, we found that progesterone up-regulated CD82 expression in both RL95-2 and HEC-1A cells. These results in vitro are consistent with the immunohistochemical staining results of patient tissues, and suggest that the progesterone facilitates embryo implantation by up-regulating CD82-dependent embryo–endometrial adhesion.
In the window of implantation, other signal transduction molecules expressed in the endometrium are known to regulate embryo adhesion. Among them, FAK is well recognized as a key molecule. FAK is an important tyrosine kinase, which plays a decisive role in signaling regulation between cells and extracellular matrices, 36 and mediates cell adhesion. 37 CD82 has been shown to mediate cell adhesion and signal transduction. 38,39 Blocking CD82 with anti-CD82 antibody led to increased endogenous Src kinase activity in human prostate cancer cells, 40 and high activity of Src kinase results in the inhibition of the FAK signaling pathway. 37 This evidence suggests that CD82-mediated cell adhesion may be associated with the FAK signaling pathway. In terms of embryo–endometrium adhesion, Ruseva et al. 41 found that CD82 over-expression drastically stimulates integrin αvβ3/vitronectin-dependent ovarian-cancer-cell adhesion. Previous work in our laboratory also showed that blocking RL95-2 cells with integrin αvβ3 antibody reduced the adhesion of embryonic cells and inhibited the phosphorylation of FAK. 42 Therefore, we hypothesized that the increase in the adhesion of embryonic cells to the RL95-2 monolayer, representative of receptive endometrium, due to enhanced expression of CD82, may also be related to the FAK signaling pathway. Results from the current study support this hypothesis.
In conclusion, CD82 expression, up-regulated by progesterone, is likely related to endometrial receptivity, and can serve as a marker for the window of implantation. Specifically, our results suggest that CD82 facilitates the adhesion between the embryo and the endometrium, and this process may involve the FAK signaling pathway. Finally, CD82-facilitated adhesion may be what links progesterone and improved embryo implantation.
Footnotes
ACKNOWLEDGEMENTS
The project was supported by the National Natural Science Foundation of China Research Grant (No. 31070729). We thank Dr David L K Chen for his careful review of the manuscript.