
Abstract
The tumor-suppressor ING3 has been shown to be involved in tumor transcriptional regulation, apoptosis and the cell cycle. Some studies have demonstrated that ING3 is dysregulated in several types of cancers. However, the expression and function of ING3 in human hepatocellular carcinoma (HCC) remains unclear. The aim of this study is to investigate ING3 expression in hepatic tumors and its clinical relevance in hepatic cancer. The expression of ING3 protein was examined in 120 dissected HCC tissues and 47 liver tissues adjacent to the tumor by immunohistochemical assays and confirmed by Western blot analysis in 20 paired frozen tumor and non-tumor liver tissues. The relationship between ING3 staining and clinico-pathological characteristics of HCC was further analyzed. The mRNA expression of ING3 in the dissected tissues was also analyzed by reverse transcriptase polymerase chain reaction (RT-PCR) and realtime PCR. Both mRNA and protein concentrations of ING3 were found to be downregulated in the majority of HCC tumors in comparison with matched non-tumor hepatic tissues. Analysis of the relationship between ING3 staining and clinico-pathological characteristics of HCC showed that the low expression of ING3 protein is correlated with more aggressive behavior of the tumor. Kaplan–Meier curves demonstrated that patients with a low expression of ING3 have a significantly increased risk of shortened survival time. In addition, multivariate analysis suggested that the level of ING3 expression may be an independent prognostic factor. Our findings indicate that ING3 may be an important marker for human hepatocellular carcinoma progression and prognosis, as well as a potential therapeutic target.
Introduction
Human hepatocellular carcinoma (HCC) is a highly fatal cancer that has poor prognosis. It ranks as the third cause of cancer death, especially in East Asia and sub-Saharan Africa. 1,2 HCC is the second leading cause of cancer death among men in China. 3 HCC has also been increasing in the USA and Europe in recent years. 4,5 Currently, only 30% of HCC patients are eligible for potentially curative treatments such as liver resection, transplantation or local ablation, according to an established therapeutic algorithm. 6 The majority of patients present with advanced or unresectable disease, making the prognosis of HCC dismal, and systemic chemotherapy is quite ineffective in HCC treatment. To improve patient survival, it is of great clinical importance to find reliable prognostic markers that could predict relapse and dissemination of HCC. The molecular pathogenesis of HCC seems to involve multiple genetic aberrations in the molecular control of hepatocyte proliferation, differentiation and the maintenance of genomic integrity. This process is influenced by the cumulative activation and inactivation of oncogenes, tumor suppressor genes and other genes. Clarification of the genes involved in the development of HCC will contribute to our understanding of mechanisms of hepatocarcinogenesis.
The human ING gene family, including ING1, ING2, ING3, ING4 and ING5, has been of interest because of these genes’ putative roles as tumor suppressors. 6,7 The ING3 gene is mapped to 7q31.3, consists of 12 exons and encodes a 46.8 kDa protein, which shares the conserved carboxyl-terminal plant homeodomain and a nuclear localization sequence with other ING family members. 8 ING3 has been found to regulate the cell cycle and apoptosis of cancer cells. Several studies have reported that ING3 plays critical roles in several types of cancers. Similar to ING1, ectopic expression of ING3 in RKO cells decreased colony formation, possibly by decreasing the number of cells in the S phase. 8,9 ING3 over-expression also upregulated Fas expression and promoted apoptosis in ultraviolet (UV)-treated cells. 10 However, it was found that the ING3 gene sequence of the Caenorhabditis elegans was highly similar to the human ING3 gene. ING3 co-localizes with chromatin in embryos, the germline and somatic cells. The ING3 gene is part of an operon but is also transcribed from its own promoter. Both ING3(RNAi) and ING3 mutant strains demonstrate that the gene likely functions in concert with the C. elegans p53 homolog, cep-1, to induce germ-cell apoptosis in response to ionizing radiation. 11 These findings, together with other research, 8 suggest that ING3 may cooperate with p53 to induce apoptosis and cellular senescence.
Biochemical purification of ING protein-containing complexes from HeLa cell nuclear extracts revealed that ING3 is found as a subunit of the NuA4/Tip60 HAT complex. Tip60 is involved in the cellular response to DNA damage, apoptosis, metastasis suppression and maintenance of embryonic stem cell identity. 12,13 Mice deficient for Tip60 die during embryogenesis prior to implantation (E4), and haploinsufficiency for Tip60 results in accelerated lymphomagenesis in transgenic Em-Myc mice. 14 These data suggest that ING3 might also function in these diverse processes through its association with chromatin remodeling. 15
Reduced ING3 expression was observed in malignant melanoma when compared with the levels of ING3 expression in dysplastic nevi. 16 As with ING2, the decreased level of nuclear ING3 was associated with a poor five-year survival rate of cancer patients. Survival rates were 93% for strong nuclear ING3 staining and dropped to 44% for patients with negative to moderate nuclear staining. 16 ING3 expression has also been reported to be associated with poor overall survival (OS) and tumor initiation in head and neck cancers; at five-year follow up, approximately 60% of the patients with normal to high ING3 expression survived, whereas this was 35% in the patients with low ING3 expression in head and neck cancers, 17 and a missense mutation in ING3 codon 20 has been observed in head and neck squamous cell carcinoma. 18 ING3 shares structural similarities with p33ING1b and ING2 which may play important roles as tumor suppressor genes in HCC. 19,20 p53 plays a critical role in HCC; 21,22 it has a close relationship with ING3. However, the role of ING3 in HCC remains largely unclear. In the present research, we investigated the expression of ING3 protein in HCC using immunohistochemistry and Western blotting assays. We then determined the association between the expression of ING3 protein and clinico-pathological features and evaluated prognostic value to postresectional survival. Furthermore, we investigated the expression of ING3 mRNA in HCC by reverse transcriptase polymerase chain reaction (RT-PCR) and realtime PCR, and determined whether the dysregulation of ING3 in HCC occurred at the transcriptional or translational level.
Materials and methods
Tissue samples
For Western blot, RT-PCR and realtime PCR analysis, 20 paired fresh surgical hepatocellular carcinoma, adjacent normal and fibrotic tissue samples (>2 cm from border of the tumor) were obtained from HCC patients who underwent curative hepatectomy from October 2008 to April 2009 at the Department of General Surgery (First Affiliated Hospital, Harbin Medical University, Harbin, China). The adjacent normal samples were from outside the margin of the tumors, and had no cancer cell invasion as demonstrated by hematoxylin and eosin (H&E) staining. Informed consent was signed by all patients. All fresh tumor tissues were immediately frozen in liquid nitrogen and stored at −70°C until use.
For immunohistochemistry (IHC) analysis, HCC tissues (mean size, 3.8–6.5 cm) were obtained from 120 patients (88 men and 32 women, age range from 30 to 79 y; median, 54 y). All underwent curative surgery with postoperative systemic chemotherapy. Follow-up data of the postoperative outcomes were retrospectively obtained from all of the patients, and the follow-up period ranged from 12 to 124 months (median, 35 months). Sixty-one patients were positive for hepatitis B virus (HBV) surface antigen, two were positive for hepatitis C virus (HCV) antibody and the others were negative for both. Histological grades of hepatocellular tumor were classified according to the General Rule for Clinical and Pathological Study of Primary Liver Cancer adopting the Tumor-Node-Metastasis (TNM) classification (International Union Against Cancer, 1987) as follows: well (n = 20), moderately (n = 39) and poorly differentiated (n = 61). For histological examination, all tissue portions of tumor and surrounding non-tumor were fixed in 10% formalin and embedded in paraffin for H&E staining.
Cell lines
The SK-Hep-1 HCC cell line and human hepatic cell line L02 were obtained from the Chinese Academy of Sciences, Shanghai Research Center (Shanghai, China). The Huh-7 HCC cell line was bestowed by Professor Niu JunQi (The First Affiliated Hospital of Bethune Medical College, Jilin University, Changchun, China). HepG2, BEL7402, SMMC7721 and Hep3B HCC cell lines were from our institute. All cell lines were cultured under recommended conditions.
Immunohistochemistry
Sections (4 μm), cut from the original paraffin blocks, were deparaffinized in xylene and re-hydrated in graded alcohols and distilled water. After inhibition of endogenous peroxidase activity for 10 min with 0.3% H2O2, the sections were incubated overnight with rabbit anti-human ING3 at 4°C. The sections were then incubated with biotinylated anti-rabbit IgG (ZhongShan Goldenbridge Biotechnology Co, Ltd, Beijing, China) for 30 min at 37°C and stained with 3,3-diaminobenzidine (Zhongshan Goldenbridge Biotechnology Co, Ltd). Finally, the sections were counterstained with hematoxylin. Normal goat serum and blocking with antibody-specific peptides were used as a negative control for the staining reactions. All sections were examined by the immunohistochemical assessment system, Motic Images Advanced 3.0 (Motic China Group Co Ltd, Xiamen, China). Two independent pathologists evaluated these sections in a double-blind manner. Both investigators re-assessed the sections under a multihead microscope, and in the case of disagreement, re-investigated the slides. The staining was semi-quantitatively evaluated by assigning a score to the proportion of the area positively stained and was classified into four groups: 23,24 +, 0–10%; ++, 10–25%; +++, 25–50%; and ++++, >50%; with the intensities ‘+’ and ‘++’ labeled as ‘low expression’ and the intensities ‘+++’ and ‘++++’ as ‘high expression.’
Western blot analysis
Protein extracts were prepared from 20 paired frozen tissues. The total protein concentrations of sample were measured based on the bicinchoninic acid (BCA) method (BCA protein essay kit; Beyotime Institute of Biotechnology, Jiangsu, China). Proteins were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels and electro-transferred onto nitrocellulose membranes. The blots were blocked in 5% non-fat dry milk in Tris-buffered saline containing 0.1% Tween 20, and incubated with primary ING3 antibody (1:100 dilution; Abcam Inc, Cambridge, UK) or glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA) overnight at 4°C. Blots were washed three times in Tris-buffered saline (TBS) containing 0.1% Tween 20 (TBST) and incubated with 700DX Conjugated Affinity Purified anti-Rabbit or anti-Mouse IgG (1:5000 dilution; Rockland Immunochemicals, Inc, Gilbertsville, PA, USA) at room temperature for one hour. Blots were then washed three times in TBS and the signal was recorded using the Odyssey Imaging System (Gene Company Limited, Hong Kong, China). The density of ING3 concentration was determined using Quant one analysis image software (Bio-Rad Laboratories, Hercules, CA, USA) and normalized with GAPDH.
RT-PCR
Total RNA was isolated from the fresh tumor and non-tumor tissues using Trizol reagent (Invitrogen, Life Technologies, Carlsbad, CA, USA). RNA was reverse-transcribed to cDNA with oligo dT. Subsequently, the resulting cDNA was subjected to PCR using a PCR kit (Takara, Dalian, China). PCR analysis was performed using the following sets of primers: for ING3: sense 5′ ATCTTCGGCACTTATGAC AAC 3′, antisense 5′ AGGGCAATCTTGGTTATCACA 3′; and for GAPDH: sense 5′ GGATTTGGTCGTATTGGG 3′; antisense 5′ GGAAGATGGTGATGGGATT 3′. The PCR profile was performed under the following conditions: for ING3, 95°C for five minutes, 95°C for 30 s, 52.8°C for 25 s, 72°C for 30 s for a total of 30 cycles and 72°C for eight minutes; for GAPDH, cycle conditions applied were: 95°C for five minutes, 30 cycles of 95°C for 30 s, 51.4°C for 25 s, 72°C for 30 s and a final extension at 72°C for eight minutes. The RT step was omitted for negative controls. The amplification products were separated by 1.5% agarose gels and stained with ethidium bromide. The density of ING3 concentration was performed using Quant one analysis software image software and normalized with GAPDH.
Quantitative realtime PCR
Quantitative realtime PCR was performed using the LightCycler system together with the LightCycler DNA Master SYBR Green I Kit (Bioer Technology Co, Ltd, Hangzhou, China). Gene expression was determined using the Roter Gene 3000A systerm (Corbett Life Science Inc, San Francisco, CA, USA). For primer sequences for GAPDH and ING3 amplification, please see the previous ‘RT-PCR’ section. The comparative CT method, normalizing CT values to the housekeeping gene GAPDH and calculating relative expression values, quantified gene expression. Postamplification melting curve analyses were performed to assure product specificity. Relative ING3 expression levels were standardized in comparison with the expression level of matched normal hepatic tissue samples. All reactions were performed in triplicates.
Statistical analysis
Statistical analysis was performed using SPSS version 13.0 (SPSS Inc, Chicago, IL, USA). Differences in ING3 expression between hepatic cancer tissues and adjacent tissues were analyzed by non-parametric Mann–Whitney U test. Kruskal–Wallis and non-parametric Mann–Whitney U tests were employed for analysis of the association between ING3 expression and clinico-pathological parameters. Disease-free survival (DFS), OS and their statistical significance were analyzed by the log-rank test. In the above analyses, P < 0.05 was considered statistically significant. The prognosis meaning of ING3 expression was assessed by the method of Cox regression, and significance was affirmed when P≈0.1.
Results
Expression of ING3 in HCC and adjacent normal tissues by IHC
To investigate whether ING3 protein was dysregulated in HCC, the expression of ING3 in HCC (n = 122) and adjacent non-tumor tissue (n = 47; 29 normal hepatic tissues and 18 hepatic cirrhosis tissues) specimens was analyzed by IHC. The expression of ING3 protein was located in the cytoplasm. In the non-tumor tissues, ING3 expression was mostly in the cytoplasm; however, weak nuclear staining was also observed (Figures 1a and b). The expression of ING3 was significantly decreased in HCC when compared with the adjacent non-tumor tissues (P < 0.001) (Figure 1c).
Expression of ING3 protein in human hepatocellular carcinoma (HCC) and adjacent non-malignant hepatic tissues. (a) Immunohistochemical staining of ING3 protein in hepatic tissues. Normal epithelium tissues showed moderate to strong ING3 immunosignals. (b) Hepatic cancer exhibited negative or weak ING3 immunostaining. (a, b) ×200 magnification. (c) Summary results of ING3 protein immunohistochemical staining in hepatic carcinoma and adjacent non-malignant hepatic tissues. The distribution of ING3 expression in each histopathological group of hepatic carcinoma and non-cancerous hepatic tissues is indicated as follows: black, intensity – ++++; dark gray, intensity – +++; light gray, intensity – ++; and white, intensity – +. The levels of staining are significantly decreased for ING3 expression in hepatic carcinoma (P < 0.001). These results suggest that ING3 plays a negative role in neoplastic transformation of the hepatocellular epithelials
Western blot analysis for ING3 protein
To confirm the specificity of ING3 protein and to check the level of ING3 expression, 20 paired tumor and non-tumor liver tissues were analyzed by Western blot analysis (Figure 2). Most non-tumor tissues (16/20, 80%) of chronic hepatitis or liver cirrhosis had a clear expression of ING3 protein. However, HCC tissues did not express ING3 or a slight band appeared for ING3. Thus, our results demonstrated that there was a low level of ING3 protein in HCC.
The expression of ING3 protein in the hepatic cancer and matched paracancerous tissues. Of 20 cancerous samples from hepatic cancer patients, 16 showed low expression of ING3 protein compared with the matched paracancerous tissues (T: cancerous samples; N: matched paracancerous tissues)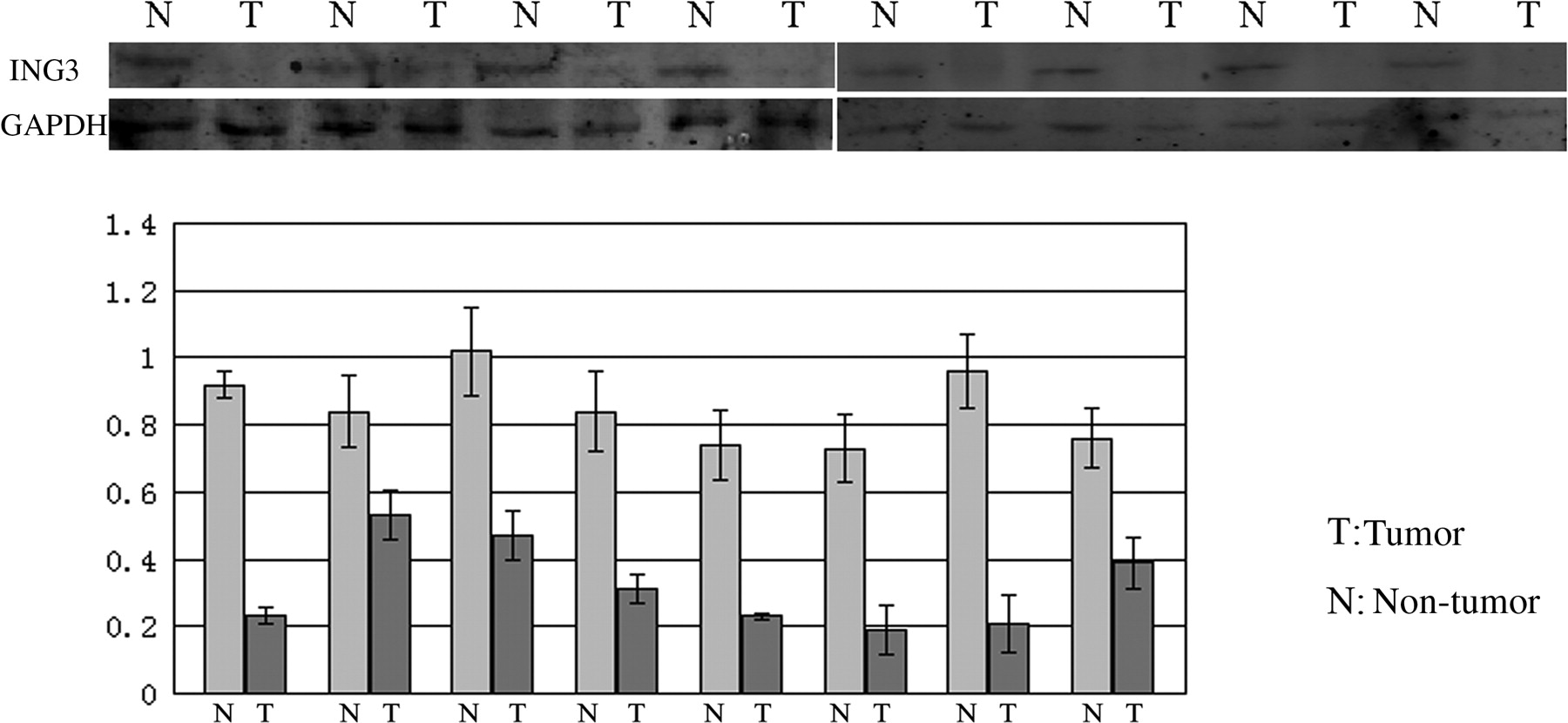
Differential expression of ING3 mRNA in HCC and non-tumor hepatic tissues
In this study, our results have shown that the expression of ING3 protein was lower in HCC tissues than that in non-tumor hepatic tissues at the protein concentration (Figure 3). In order to investigate whether mRNA expression of ING3 was deregulated in HCC, we analyzed the ING3 mRNA expression in the 20 fresh HCC tissue samples and their matched non-tumor tissues. Lower expressions of ING3 mRNA were found in 16 of the HCC tissue samples compared with the matched non-tumor hepatic tissues (Figures 3a and b). To confirm the RT-PCR results, we further investigated ING3 mRNA expression in the above 20 fresh HCC tissues and their matched non-tumor tissues by realtime PCR. We found that the ING3 mRNA level was lower in 16 of the HCC tissues than that in the matched non-tumor hepatic tissues (Figure 3c).
The mRNA expression of ING3 in hepatic tumor and adjacent non-malignant tissues. (a, b) ING3 mRNA expression in hepatic tumor and matched adjacent non-malignant tissues was examined by reverse transcription polymerase chain reaction (PCR). Of 20 hepatic cancer tissues from cancer patients, 16 showed a low expression of ING3 mRNA when compared with the matched paracancerous tissues. (c) The expression of ING3 mRNA in hepatic tumor and matched adjacent non-malignant tissues was determined by realtime PCR. Of 20 hepatic cancer tissues from patients, 16 showed lower expression of ING3 mRNA in comparison with the matched paracancerous tissues (T: cancerous samples; N: matched paracancerous tissues)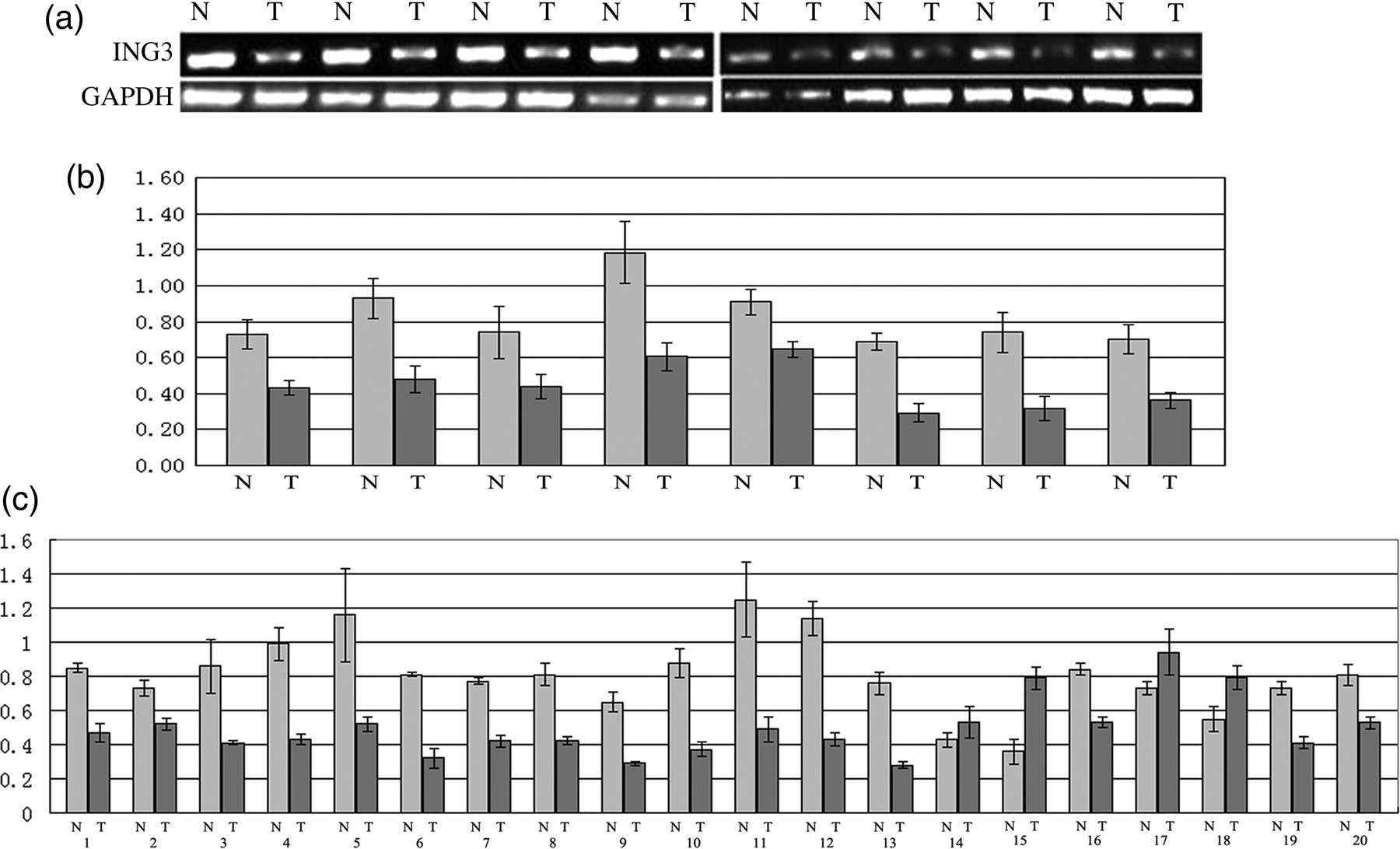
Expressions of ING3 protein and mRNA in HCC cell lines
To investigate the expression of ING3 protein in hepatic cancer cell lines, six hepatic cancer cell lines, SK-Hep-1, Huh-7, HepG2, BEL7402, SMMC7721 and Hep3B HCC cell lines and immortalized human embryo hepatic cell line L02, were analyzed by Western blot analysis. The ING3 protein expression was lower in highly malignant hepatic carcinoma cell lines Sk-hep-1 and SMMC7721 than that in moderately malignant hepatic carcinoma cell lines HepG2 and Hep3B, the well-differentiated cell lines Huh-7, and the immortalized human embryo hepatic cell line L02. However, there was no marked difference in ING3 expression between the well-differentiated hepatic carcinoma cell lines and the immortalized human embryo hepatic cell line L02. Of these six HCC cell lines, Sk-hep-1 expressed the lowest ING3 protein and Huh-7 expressed the highest ING3 protein (Figures 4a and b).
The expression of ING3 protein and mRNA in several hepatic cancer cell lines. Expression of ING3 protein in the cell lines was determined by Western blotting, reverse transcription polymerase chain reaction (RT-PCR) and realtime PCR. (a, b) Expression of ING3 protein in the normal hepatic and HCC cells; (c, d) expression of ING3 mRNA in the normal hepatic and HCC cells by RT-PCR; and (e) expression of ING3 mRNA in the normal hepatic and HCC cells by realtime PCR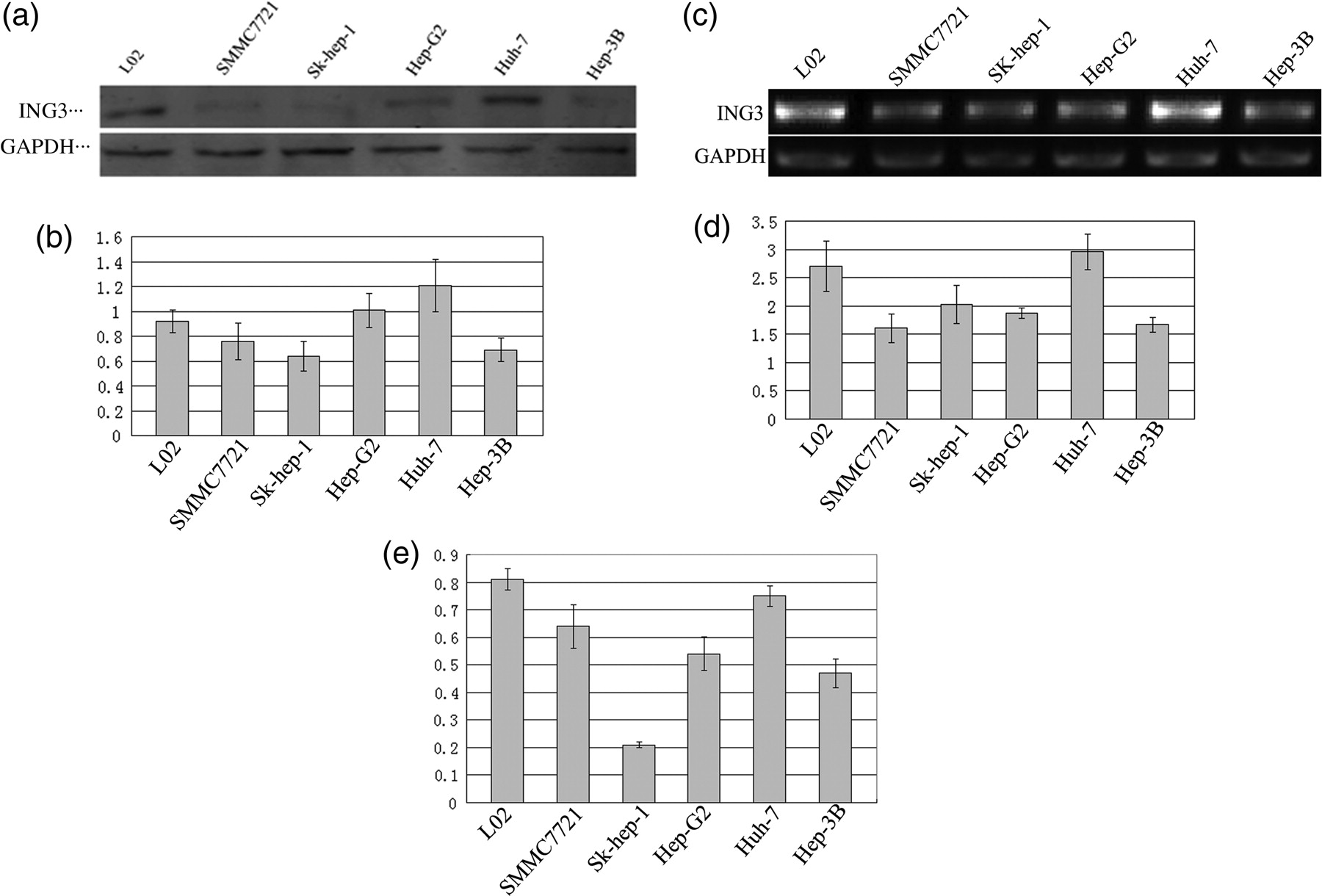
We further measured the mRNA expression of ING3 mRNA in these six HCC cell lines by RT-PCR analysis. Huh-7 cells showed the highest expression levels of ING3 mRNA among all six HCC cell lines. Contrary to Huh-7, SK-hep-1, with the high metastatic potential, had the lowest levels of ING3 mRNA expression. Hep3B, a non-metastatic cell line has a higher expression level of ING3 mRNA than the other three cell lines (SMMC7721, BEL7402 and HepG2), which has moderate metastatic potential (Figures 4c and d).
To confirm the results of RT-PCR, quantitative realtime PCR was performed in the above-mentioned six HCC cell lines and L02, and similar results were observed (Figure 4e).
Relationship between ING3 protein expression and clinico-pathological characters
ING3 protein expression was analyzed with pathological features
†1987 International Union Against Cancer Tumor-Node-Metastasis (TNM) stage
*P < 0.05; **P < 0.01
Prognostic significance of ING3 protein
We investigated whether the expression of ING3 protein in HCC can serve as a prognostic biomarker. We followed 122 patients who underwent curative surgical operations, and used the Kaplan–Meier method and log-rank test to analyze their DFS and OS rates (Table 2). We followed up with the patients for a period of 11–124 months (median, 35 months). During this period, 58 (58/120, 48.3%) patients died; of these 58 patients, 34 (34/58, 58.6%) patients died from recurrence or metastasis and 24 (24/58, 41.4%) patients died from other diseases. Of the 62 (62/120, 51.7%) patients who survived, three patients developed a recurrence or distal metastasis and the other 59 patients survived with no tumor recurrence or metastasis. Kaplan–Meier survival curves (Figure 5) showed that expression of ING3 was positively correlated with DFS and OS. Patients with high ING3 expression had a significantly longer DFS and OS time than those with low expression (P < 0.01) (Table 2).
Kaplan–Meier survival curves for patients with high and low expression of ING3 protein. Patients were followed up for a period of 11–124 months (median, 35 months). Kaplan–Meier survival curves showed that the expression of ING3 was positively correlated with disease-free survival (DFS) and overall survival (OS). (a) DFS curve of ING3 and (b) OS curve of ING3 OS and DFS in 120 patients with hepatic cancer (Cox regression) Factors affecting the patient's prognosis through Cox regression analysis. ING3 can be a positive independent factor contrary to clinical stage and lymph node metastasis. Intrahepatic metastasis could be a negative independent factor of OS OS, overall survival; DFS, disease-free survival; TNM, Tumor-Node-Metastasis *P ≈0.1; **P < 0.01
We further investigated whether the expression of ING3 protein can serve as an independent prognostic biomarker in HCC and analyzed the DFS and OS rate of them using a Cox regression analysis (Table 2). Although the patient's age, gender, vascular invasion, capsular formation, intrahepatic metastasis, portal vein tumor thrombus, tumor size and cancer cell differentiation have no impact on DFS by COX regression analysis (P > 0.05), significant prognostic influences of lymph node metastasis, clinical stage, capsular invasion and ING3 expression were found. However, lymph node metastasis, clinical stage, intrahepatic metastasis and ING3 expression were found to have significant impact on OS by COX regression analysis (P≈0.1).
Discussion
HCC is a highly fatal cancer especially in East Asia and sub-Saharan Africa. Compared with research on activation of cellular oncogenes, more studies concentrate on inactivation of tumor suppressor genes such as p53. 21,22 Molecular genetic studies have shown that loss of heterozygosity at specific chromosomal loci of 7q31 occurs very frequently in HCC. 25–27 The tumor suppressor genes always locate on such loci and their inactivations are involved in the development of HCC. The human ING3 gene is located on deleted region 7q31. However, at present, few tumor suppressor genes located in this regions have been identified. Considering the structural homology, ING3 may share functional similarities with ING1 and ING2, and the reduced expression of ING1 and ING2 genes have been detected in HCC. 19,20 There have been a few other studies demonstrating the involvement of ING3 as a tumor suppressor in certain malignancies and including melanoma and head and neck cancers, 16–18 so we presumed that ING3 may play a role in tumor development and progression of HCC. To our knowledge, there are no reports about studies of ING3 in human HCC samples. In our study, to evaluate its potential clinical relevance, we investigated the expression pattern of ING3 in HCC.
In this study, we investigated whether ING3 might play a role in the tumor progression of HCC. To our knowledge, there are no reports of ING3 in human HCC. In this study, to evaluate its potential clinical value, we investigated the expression pattern of ING3 in HCC. The present study aimed to examine the mRNA and protein expressions of ING3 in HCC and to address the relationship between its expression and clinico-pathological features. We also suggested that downregulation of ING3 might be a new negative prognostic marker in HCC. Finally, we demonstrated that the dysregulation of ING3 in HCC occurred at the levels of both transcription and translation.
The expression of ING3 protein in both liver tumor and non-tumor tissues was examined using immunohistochemistry assays. A decreased expression of ING3 was found in most liver cancer samples in comparison with adjacent non-tumor tissues. Interestingly, in contrast to ING3 which expresses as a nuclear protein in melanoma, 16 the expression of ING3 was located mostly in the cytoplasm rather than in the nucleus. In agreement with our findings, some other studies 27,28 also found that p33ING1b and ING2, homologs of ING3, were aberrantly localized in the cytoplasm in brain tumor and HCC, respectively. Recent studies have reported that mislocalization of p33ING1b contributes to the generation of specific tumors. In melanoma, papillary thyroid carcinoma, ductal breast carcinoma and acute lymphoblastic leukemia, increased levels of cytoplasm p33ING1b were reported, concomitant with loss of nuclear localization. 29,30 It is possible that mislocalization of ING2 from the nucleus to the cytoplasm could trigger degradation and loss of its function. It was demonstrated that ING3 was degraded by the SCF (Skp2)-mediated ubiquitin–proteasome system. 31 As we know, the ubiquitin–proteasome is located in the cytoplasm. All ING proteins contain a nuclear localization sequence (NLS), and some ING proteins appear to have multiple NLS. To date, the role of the NLS has been studied extensively only for ING1. 32,33 NLS deletion results in cytoplasm accumulation of the protein. Localization of ING proteins to the nucleus has been proposed to be critical to their function, as is evident by the observation of loss of nuclear ING1 staining in a number of cancers, 34 and because deleting the entire NLS of ING4 resulted in a protein that could no longer bind p53 in co-transfection experiments. 35 Additionally, there are two copies of a putative nucleolar translocation signal (NTS) contained within the NLS of ING1, and translocation of ING1 to the nucleolus following exposure to UV light appears to be required for ING1-associated apoptosis. 32 It is currently unknown whether ING3 protein has a similar NTS or can be detected in the nucleolus compartment. The precise mechanism of ING3 in our observations remains elusive and need further investigation.
In addition, there was a correlation of the downregulation of ING3 protein with tumor grades. The expression of ING3 was negatively related with tumor size (P < 0.05) and clinical stage (P < 0.05), which suggests that the deregulation of ING3 played a continued role throughout the processing of cancer advancement and development. Furthermore, the negative expression of ING3 is also associated with elevated AFP serum levels (P < 0.05). The aberrant expression of AFP is a characteristic of HCC and serves as a diagnostic and specific tumor marker. 36 It has been reported that p53 negatively regulates AFP gene expression through alteration of chromatin structure at the core promoter. 37,38 Some results 8,11 demonstrated that ING3 gene regulates p53. It is possible that ING3 regulates the AFP gene through modifying p53 function. In fact, it was reported that p33ING1b, ING2 strongly repressed AFP promoter activity. 20,39 It is concordant with our results, which showed that negative ING3 was also correlated with high levels of serum AFP. However, the mechanism still needs to be elucidated. In this study, we found no relationship between expression of ING3 and age, gender and capsular formation.
Prognostic molecular biomarkers are valuable for the clinician to evaluate patients and to aid in tumor control. We found that the decreased expression of ING3 protein was significantly correlated with poor patient outcomes. In this study, the assessment of the correlation between ING3 expression and the duration of patient survival (Figure 5) showed that the lower the ING3 expression level, the shorter the patient survived (DFS and OS), and conversely, the higher the ING3 expression level, the longer the period of patient survival. Furthermore, results from multivariate Cox regression analysis show that the final model of the patient's age, gender, differentiation, intrahepatic metastasis, capsular formation, portal vein tumor thrombus and vascular invasion had no effects on DFS (P > 0.15), whereas significant prognostic influences of tumor stage, capsular invation, lymph node metastasis and ING3 expression on DFS were found (P < 0.15). As independent prognostic factors of OS, tumor stage, lymph node metastasis, intrahepatic metastasis and ING3 expression may shorten the OS of patients with HCC after surgery. Our data indicate that the expression of ING3 is significantly correlated with OS and DFS, which is consistent with the former findings that reduced ING3 expression was significantly correlated with a poorer survival of patients with HCC.
The expression of ING3 protein also showed a low level in HCC by Western blotting. We further investigated ING3 mRNA expression in tumor and non-tumor hepatic tissues. We found that the mRNA expression levels in HCC samples were lower than that in non-tumor tissues. These results indicated that the deregulation of ING3 in HCC occurred at both transcription and translation levels.
Immunoblot analysis of ING3 expression in HCC cell lines showed that the expression of ING3 protein was inversely correlated with the tumor malignant degree. Well-differentiated and low metastasis potential cell lines consistently expressed relatively high ING3 protein levels, whereas moderate to high metastasis potential cell lines showed lower ING3 expression levels. Using RT-PCR and realtime RT-PCR analysis, we found similar results, suggesting that the upregulation of ING3 occurred at the transcriptional level in HCC cells. Furthermore, a significant difference was found in ING3 mRNA expression between the HCC cell lines with different malignant degrees, which is positively correlated with the ING3 protein expression. Thus, transcriptional mechanisms may account for the dysregulation of ING3 protein expression in HCC cell lines.
Hepatocellular carcinogenesis is a multistep and complex process involving several etiological agents that exert differential effects on the molecular pathways involved. Mutations in the p53 gene found in HCCs have shown allelic loss at the 17p13.1 region of the chromosome. 40,41 It has been reported that ING3 cooperates with p53 to regulate apoptosis death receptor/extrinsic apoptotic pathways. 8
The ING proteins have multiple domains that are potentially capable of binding to many partners. It is conceivable, therefore, that such proteins could function similarly within protein complexes. In this case, within this family, each function could be attributed to a specific domain.
ING3 is a component of the NuA4/Tip60 HAT complex and is required for acetylation of chromatin substrates including the N-terminal tails of histones H4 and H2A. 42,43 This 12-component acetyltransferase complex is responsible for acetylation of the N-terminal histone H4 and H2A tails. In addition to Tip60, which contains the catalytic MYST domain, the hNuA4 complex includes Eaf6, the enhancer of polycomb homology domain 1 (EPC1), and the ING3 subunit among others. ING3 plays a key role in regulating the enzymatic activity of NuA4/Tip60. It bridges the HAT complex to chromatin through concurrently associating with the EPC1 subunit and H3K4me3 through its N-terminal region and the C-terminal PHD finger, respectively. Similarly to ING1 and ING2, the ING3 tumor suppressor is also thought to cooperate with p53 in transcription regulation, cell cycle control and apoptosis. 44
The leucine zipper-like (LZL) domain is found in the N-terminus of all ING proteins, except ING1. This domain consists of four to five conserved leucine or isoleucine residues spanning every seven amino acids (forming a hydrophobic patch near the N-terminus) with a similar leucine distribution for ING3 to ING5. 7,45,46 However, little is known about the function of LZL and it been has reported that this region is responsible for homo- and hetero-oligomerization between ING3 and the other members of the family. 45
It is possible that the ING3 protein transduces stress signals by its distinct structure subsequently contributes to regulate tumor development. However, these mechanisms cannot by themselves explain all the events in which ING proteins are involved.
The understanding of the molecular pathways leading to the development of HCC may provide important data to develop new therapies. Similarly to other tumor types, an imbalance between unrestrained cell proliferation and impaired apoptosis appears to be a major unfavorable feature of HCC. 47 Recent studies have documented that ING3 gene was upregulated after drug treatment, resulting in oxidative stress and endoplasmic reticulum stress in various cancer cell lines. 48 These results showed that ING3 expression might also correlate with the response of therapeutic interventions.
Conclusively, our findings suggested that the downregulation of ING3 frequently occurred in HCC and decreased expression of ING3 in HCC may lead to a poor prognosis. Deregulation of ING3 occurs in HCC at both transcription and translation levels. Our findings suggest that ING3 may be a candidate tumor suppressor gene for HCC and may have diagnostic and therapeutic potential for patients with HCC. However, more studies should be done to elucidate the role and molecular mechanism of ING3 in hepatic carcinogenesis and the interaction of ING3 and AFP expression.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the Program for Innovative Research Team (in Science and Technology) in Higher Educational Institutions of Heilongjiang Province (No. 2009td06), Heilongjiang Provincial Department of Education Funding (No. 11541198 and No. 12521276), First Affiliated Hospital of Harbin Medical University Fund (No. 2011BS08) and the Science and Technology Department of Heilongjiang Provincial Laboratory Animal Special Funds (No. PC09S03). We thank Dr Nishant Bhatta from the Department of Hepatic Surgery of the First Affiliated Hospital of Harbin Medical University for reading and revising this manuscript.