
Abstract
Inflammatory bowel disease (IBD) comprises primarily the chronic relapsing inflammatory disorders, Crohn's disease and ulcerative colitis, with the former affecting any part of the gastrointestinal tract and the latter mainly afflicting the colon. The precise etiology of IBD remains unclear, and it is thought that interactions among various factors, including genetic factors, the host immune system and environmental factors, cause disruption of intestinal homeostasis, leading to dysregulated inflammatory responses of the gut. As inflammation is intimately related to formation of reactive intermediates, including reactive oxygen and nitrogen species (ROS/RNS), oxidative stress has been proposed as a mechanism underlying the pathophysiology of IBD. This review is intended to summarize succinctly recent new experimental and clinical evidence supporting oxidative stress as a pathophysiological component of IBD and point to the potential of using antioxidant compounds as promising therapeutic modalities of human IBD. The sources of ROS/RNS and the redox signaling mechanism underlying oxidative stress and inflammation in IBD are discussed to provide insight into the molecular basis of oxidative stress as a pathophysiological factor in IBD.
Introduction
Inflammatory bowel disease (IBD), including primarily ulcerative colitis and Crohn's disease, is a chronic, relapsing and remitting inflammatory condition of the intestine, which is characterized by rectal bleeding, severe diarrhea and weight loss. IBD afflicts individuals of both sexes throughout life, and is also a risk factor for colorectal carcinoma. 1 Currently, up to 1.4 million Americans have IBD, with ∼30,000 new cases being diagnosed each year. While IBD has, in the past, been most evident in the developed countries, its prevalence in the developing countries has been gradually increasing in recent decades. 2–4 At present, there is no cure for IBD. Current therapies involve treatment with 5-aminosalicylic acid (also known as mesalamine) and its analogs, as well as glucocorticosteroids; however, these drugs display limited beneficial actions. Immunosuppressive drugs and the recently developed anti-tumor necrosis factor (TNF-α) biological agents have also been used to control severe cases of IBD, regardless of the more serious complications and side-effects associated with them. 5 Over the long term, up to 75% of patients with Crohn's disease and 25–33% of those with ulcerative colitis will require surgery. 6 Hence, there is a great need to develop more effective therapies for IBD. In this context, advances in the understanding of the pathophysiology of IBD will certainly facilitate the development of novel and more effective mechanistically-based therapeutic modalities.
Although collectively called IBD, ulcerative colitis and Crohn's disease present clinically distinct conditions with distinguishing clinical, anatomical and histological findings. Ulcerative colitis is characterized by confluent inflammation of the colonic mucosa, extending to a variable extent from the rectum to the proximal colon. Crohn's disease, on the other hand, is characterized by discontinuous transmural inflammation, involving any portion of the gastrointestinal tract, but most commonly the terminal ileum. 7 The exact pathophysiological mechanisms of IBD remain to be fully elucidated. Recent experimental and clinical studies suggest that the initiation and progression of the disease are multifactorial, involving interactions among genetic, environmental and immune factors. 8–10 The interactions among these factors lead to uncontrolled, abnormal immune responses in the intestinal mucosa. 11 The mechanisms by which dysregulated immune responses initiate and perpetuate inflammatory intestinal damage are not entirely clear. However, substantial evidence suggests that chronic intestinal inflammation is associated with enhanced production of reactive oxygen and nitrogen species (ROS/RNS). Such an overproduction of ROS/RNS and the consequent oxidative stress and redox modulation have been demonstrated to play a critical role in the pathophysiology of IBD in both experimental animals and human subjects. 12–14 This review is intended to summarize the major recent findings on the oxidative stress and redox signaling mechanisms of IBD in both experimental models and clinical studies, and discuss the feasibility of using antioxidants as a promising approach to the intervention of the disease process.
Role of oxidative stress and redox signaling in IBD in experimental animals
Studies using various animal models have contributed greatly to our current understanding of the molecular and cellular pathophysiology of human IBD. The commonly used experimental models include chemically induced colitis (e.g. dextran sodium sulfate [DDS] and 2,4,6-trinitrobenzenesulfonic acid) and spontaneous development of intestinal inflammation in genetically engineered mice (e.g. interleukin-10 knockout mice). 15,16 Studies using these animal models have also contributed to our understanding of the oxidative stress and redox signaling mechanisms of IBD, as well as the development of antioxidant-based strategies for disease intervention. However, it should be borne in mind that animal models of IBD, especially the chemically elicited colitis in rodents, have limitations. In view of this, studies directly in human subjects are indispensible for delineating the potential role of oxidative stress and redox signaling in the pathophysiology of IBD (see later sections for discussion of human evidence).
Evidence supporting a causative role for oxidative stress in experimental IBD
As discussed below, there are four lines of experimental evidence that support a causal involvement of oxidative stress in the development of IBD in animal models.
Increased formation of ROS/RNS
Multiple experimental studies demonstrated an increased formation of ROS/RNS, including superoxide, hydrogen peroxide, hypochlorous acid and peroxynitrite in colonic mucosa in animal models of IBD. 12,13,17 Formation of these reactive species was often observed at the early stage of disease process and correlated with disease severity and progression. 12,13,17
Depletion of antioxidants and increased biomarkers of oxidative damage
In line with the augmented formation of ROS/RNS, levels of endogenous antioxidants in colonic tissue, including glutathione and copper, zinc-superoxide dismutase (Cu, ZnSOD) are decreased in experimental IBD. 18 In contrast, heme oxygenase-1 expression in colonic tissue is upregulated in early IBD, which may represent a compensatory response to oxidative stress as well as inflammation. 19 Indeed, heme oxygenase-1 is highly inducible by oxidants and inflammatory cytokines. 20 Experimental IBD is also associated with increased biomarkers of oxidative stress, such as lipid peroxidation products in both colonic tissue and peripheral blood. 21 Although these findings alone do not allow the establishment of a causative role for ROS/RNS in the development of IBD, they are consistent with the notion that oxidative stress is an important pathophysiological component of IBD.
Protection by exogenous antioxidant compounds
A critical line of evidence supporting a causative role for oxidative stress in the pathophysiology of IBD is the demonstrated protective effects of diverse antioxidant compounds in animal models of IBD. The beneficial effects are manifested as prevention of disease genesis and/or retardation of disease development. The best protective effects are often observed when the antioxidant compounds are administered before the initiation of the experimental IBD. The protective effects by antioxidant compounds are usually associated with decreased biomarkers of inflammation and oxidative stress in colonic tissue and peripheral blood. 22 Notably, two recent studies showed that systemic administration of Cu, ZnSOD attenuated the peroxidation reactions in the inflamed colon and ameliorated inflammatory responses in experimental animals with chemically induced IBD, indicating a causal role for superoxide in the pathogenesis of IBD. 23,24
Protection by transgenic overexpression of endogenous antioxidant genes and sensitization by antioxidant gene knockout
The most compelling evidence for a cause and effect relationship between oxidative stress and IBD comes from studies using genetically manipulated animal models as well as chemical models to selectively modify endogenous colonic antioxidant enzymes. For example, either transgenic overexpression of Cu, ZnSOD or ex vivo extracellular SOD gene transfer resulted in attenuation of DDS-induced colitis in mice. 25,26 Interestingly, Watterlot et al. 27 recently reported that intragastric administration of a manganese superoxide dismutase (MnSOD)-producing recombinant Lactobacillus casei BL23 strain attenuated DSS-induced colitis in mice. Induction of colonic heme oxygenase-1 by chemical inducers also ameliorated colonic inflammation and oxidative stress in animal models of IBD. Conversely, depletion of glutathione by buthionine sulfoximine, a specific inhibitor of cellular glutathione biosynthesis, was demonstrated to cause the development of colitis in mice. 28 Mice with targeted disruption of both glutathione peroxidase-1 (GPx1) and GPx2 also spontaneously developed colitis, 29 suggesting an essential role for the glutathione system in maintaining the physiological barrier of the intestine and in protecting against the genesis of IBD.
The crucial role of endogenous antioxidants in controlling intestinal inflammatory stress is also supported by the observation that Nrf2-deficient mice exhibited markedly increased susceptibility to chemically induced experimental IBD as compared with wild-type mice. 30 Nrf2 knockout also aggravated colitis-associated colorectal cancer in experimental animals. 31 As Nrf2 is a central regulator of tissue antioxidant defenses, 32 Nrf2 signaling has been proposed to be a unique target for developing antioxidant-based modalities for the intervention of IBD and its complications, such as colorectal carcinogenesis. 33
Sources of ROS/RNS in experimental IBD
A number of different cellular sources of ROS/RNS have been identified in experimental IBD. It is obvious that ROS/RNS are generated following the activation of the inflammatory cells during the disease process. 7 It is also possible that increased formation of ROS/RNS occurs during early stage of disease process and may happen even before the genesis of IBD. In this context, intestinal epithelial cells were shown to produce elevated levels of ROS/RNS after exposure to IBD-inducing insults, such as 2,4,6-trinitrobenzenesulfonic acid. 34
Inflammatory cells
Infiltration of inflammatory cells, including neutrophils and macrophages, is a hallmark of the disease pathophysiology of IBD. 11 Activation of the inflammatory cells results in the production of large amounts of superoxide and nitric oxide from NAD(P)H oxidase (NOX2) and inducible nitric oxide synthase (iNOS), respectively. Reaction of superoxide and nitric oxide generates the potent oxidant peroxynitrite, which participates in the pathogenesis of IBD. 35 Indeed, intrarectal administration of peroxynitrite was found to cause colonic inflammation and pathological changes characteristic of IBD in experimental animals. 36
Epithelial cells
NOX enzymes are expressed not only in inflammatory cells, but also in intestinal epithelial cells. 37 The notable one is NOX1, which is inducible by inflammatory cytokines, including TNF-α and interferon-γ. 38 Activation of the epithelial NOX enzymes may occur during the early stage of disease process and may even happen before the appearance of pathological changes of IBD. As these enzymes generate large amounts of ROS, the resulting oxidative stress may play a role in the genesis of IBD. This notion is supported by the demonstrated effects of exogenous antioxidant compounds and endogenously overexpressed antioxidant enzymes in retarding the disease development of experimental IBD. In addition to NOX enzymes, iNOS is also expressed in intestinal epithelial cells, whose expression is inducible by inflammatory cytokines. 39 Hence, epithelial cells contribute to the production of ROS/RNS via activation of NOX enzymes and iNOS. Exposure of intestinal epithelial cells to chemical inducers of IBD, such as 2,4,6-trinitrobenzenesulfonic acid, also led to the generation of free radical species via chemical metabolism or bioactivation. 40
NOX enzymes and iNOS not only act as major sources of ROS/RNS, but they also causally contribute to the disease development. This is evidenced by the observations that pharmacological inhibition or genetic deletion of these enzymes resulted in attenuation of the development of IBD in experimental animals. 41
Other sources
There is also evidence suggesting the involvement of other enzymes or pathways in the formation of ROS in IBD. These include xanthine oxidase, 5-lipoxygenase, cytochrome P450 enzymes and myeloperoxidase (MPO). Notably, MPO is a hallmark of gut inflammation and oxidative stress in experimental IBD. 42 Indeed, MPO activity in the inflamed intestinal tissue is a widely used biomarker in experimental IBD to assess disease pathophysiology as well as antioxidant intervention. MPO is primarily derived from infiltrated neutrophils and is involved in the generation of hypochlorous acid, a potent oxidant capable of eliciting tissue injury. 43
Molecular mechanisms of oxidative stress injury in experimental IBD
Direct oxidative damage to intestinal mucosal cells and aggravation of inflammatory stress are two important mechanistic aspects of oxidative stress in IBD. As depicted in Figure 1, these two intertwined molecular mechanistic events may contribute to both disease progress and disease genesis.
Schematic illustration of the role of oxidative stress and inflammation in the pathophysiology of IBD. As depicted, inflammation augments oxidative stress by stimulating ROS/RNS-generating systems, such as NAD(P)H oxidases (NOXs) and inducible nitric oxide synthase (iNOS), as well as the release of myeloperoxidase (MPO) from inflammatory cells. On the other hand, oxidative stress and redox signaling are intimately involved in the upregulation of inflammatory cytokines as well as the increased infiltration of inflammatory cells via stimulating signaling pathways, especially the redox-sensitive transcription factor, nuclear factor kappa B (NF-κB). IBD, inflammatory bowel disease; ROS/RNS, reactive oxygen and nitrogen species. (A color version of this figure is available in the online journal)
Oxidative damage contributing to disease progression
Overproduction of ROS/RNS via various sources, especially the activated inflammatory cells during the disease process of IBD, causes oxidative damage to cellular constituents, including lipids, leading to additional mucosal injury, dysfunction and inflammation, which further exacerbate the pathological process of IBD. In this context, lipid peroxidation products, such as reactive aldehydes, were shown to induce infiltration and activation of inflammatory cells. 44,45
Oxidative damage contributing to disease genesis
As intestinal epithelial cells can be activated by dysregulated inflammatory cytokines, augmented formation of ROS/RNS may occur at the very early stage of disease process or even before the disease development. Under this scenario, oxidative stress may play a causative role in the genesis of IBD. This notion is strengthened by the finding that depletion of intestinal epithelial antioxidants resulted in spontaneous development of IBD in animal models. 28,29 Direct inoculation of ROS/RNS to the intestine of experimental animals also caused inflammation and colitis, further supporting the concept that ROS/RNS can directly participate in the initiation of IBD. 36 Notably, the levels of endogenous antioxidants in intestinal epithelial cells are relatively low as compared with those in many other tissues (authors’ unpublished observations). The relatively deficient endogenous antioxidant defenses may render intestinal epithelia more susceptible to oxidative stress injury.
Dysregulated redox signaling contributing to perpetuation of gut inflammation
Oxidative stress not only induces direct damage to intestinal cells, but also causes dysregulated redox signaling, leading to activation of NF-κB and the subsequent overexpression of proinflammatory cytokines and adhesion molecules. 11,46 Hence, oxidative stress may contribute to the perpetuation of intestinal inflammation in IBD. 11,47 Such interactive mechanisms between oxidative stress and inflammation provide a strong basis for the development of the combination therapies of both antioxidants and anti-inflammatory drugs in the management of IBD. 47 In this context, a recent study by Palffy et al. 48 demonstrated that intragastric administration of attenuated bacteria Salmonella typhimurium SL7207 carrying a plasmid encoding either an monocyte chemoattractant protein-1 (MCP-1; a proinflammatory molecule) receptor inhibitor or Cu, ZnSOD led to amelioration of DSS-induced colitis in mice. Notably, the MCP-1 receptor inhibitor gene therapy also improved the total antioxidant capacity of the colon tissue, highlighting the intimate relationship between anti-inflammatory and antioxidative activities. 48 However, this study by Palffy et al. did not investigate the combined effects of both Cu, ZnSOD and MCP-1 receptor inhibitor gene therapies on experimental colitis. Such studies would provide important insights into the potential additive or synergistic effects of antioxidative and anti-inflammatory combination modalities for treating IBD.
Summary of the oxidative stress and redox signaling mechanisms in experimental IBD
There is substantial evidence showing sustained over-production of ROS/RNS as well as other reactive species in IBD. Oxidative stress and redox modulation not only contribute to the disease progress, but also likely participate in the early stage of the disease process. The intimate involvement of oxidative stress in the development of IBD is evidenced by the well-demonstrated effects of both exogenous and endogenous antioxidants in protecting against experimental IBD in various animal models. Depletion of intestinal epithelial antioxidant defenses by either pharmacological or genetic approaches also leads to the spontaneous development of colitis, further strengthening the concept that oxidative stress contributes to both the genesis and progression of IBD in experimental animals. Some of the above key experimental findings are also replicated in studies involving human subjects, which are discussed subsequently.
Role of oxidative stress and redox signaling in IBD in humans
Evidence for a potential causative role of oxidative stress in human IBD
Increased ROS/RNS formation and elevated biomarkers of oxidative stress
Augmented formation of ROS/RNS and biomarkers of oxidative injury, including lipid peroxidation products (e.g. reactive aldehydes, F2-isoprostanes) and protein modifications (e.g. protein carbonyls) have been demonstrated in intestinal mucosa of patients with either ulcerative colitis or Crohn's disease. 49,50 Conversely, the levels of intestinal mucosal antioxidants, including glutathione, coenzyme Q10, glutathione S-transferase (GST), superoxide dismutase, catalase, paraoxonase-1 and metallothionein, are reduced in IBD patients as compared with control subjects. 22,51,52 The levels of antioxidants in peripheral red blood cells were also shown to be decreased in patients with active IBD. 53 Of note, the antioxidant levels and the oxidative stress biomarkers are usually correlated with the disease severity and the extent of intestinal inflammation in the IBD patients.
Antioxidant gene polymorphisms
Genetic variations in antioxidant enzymes are associated with altered enzyme activity and risk of developing IBD. For example, the null genotype of GST M1 or T1 was found to be associated with the development of IBD, and the combination of the two GST genotypes further increased the risk.
54
In patients with ulcerative colitis, lack of GST M activity as measured in whole blood was correlated with an early age of onset and a more severe clinical course leading to colectomy.
55
The paraoxonase (PON) gene family maps to chromosome 7q21.3–22.1, within loci that also showed evidence of susceptibility genes for both ulcerative colitis and Crohn's disease. A case–control study reported that carriage of the PON1 R192 allele (showing increased PON1 activity) may confer protection against the development of IBD in a Jewish population.
56
In addition to GST and PON, polymorphisms of epoxide hydrolase, NAD(P)H:quinone oxidoreductase 1 and MnSOD may also influence the age of onset of ulcerative colitis as well as the efficacy of drug therapy in patients with ulcerative colitis.
57,58
Collectively, these observations support the concept that alteration of endogenous antioxidant/biotransformation enzymes and disruption of tissue redox homeostasis may contribute to the development of IBD and influence the clinical feature of the disease. In line with this notion, a polymorphism that reduces the activity of Nrf2 has been recently shown to be associated with an increased risk of developing ulcerative colitis in a Japanese population.
59
As Nrf2 acts as a central regulator of intestinal antioxidant and phase 2 defenses, pharmacological activation of this redox-sensitive molecular target may represent an effective strategy for the intervention of human IBD (Figure 2).
Activation of Nrf2 signaling as a promising strategy for the intervention of IBD. As depicted, pharmacological agents and phytochemicals can be used to activate Nrf2 to augment intestinal antioxidant and anti-inflammatory defenses. In addition, gene therapy, such as gene delivery could also be used as a potential means of enhancing the functionality of Nrf2 signaling. Note: double arrows denote that the two events are intertwined. IBD, inflammatory bowel disease. (A color version of this figure is available in the online journal)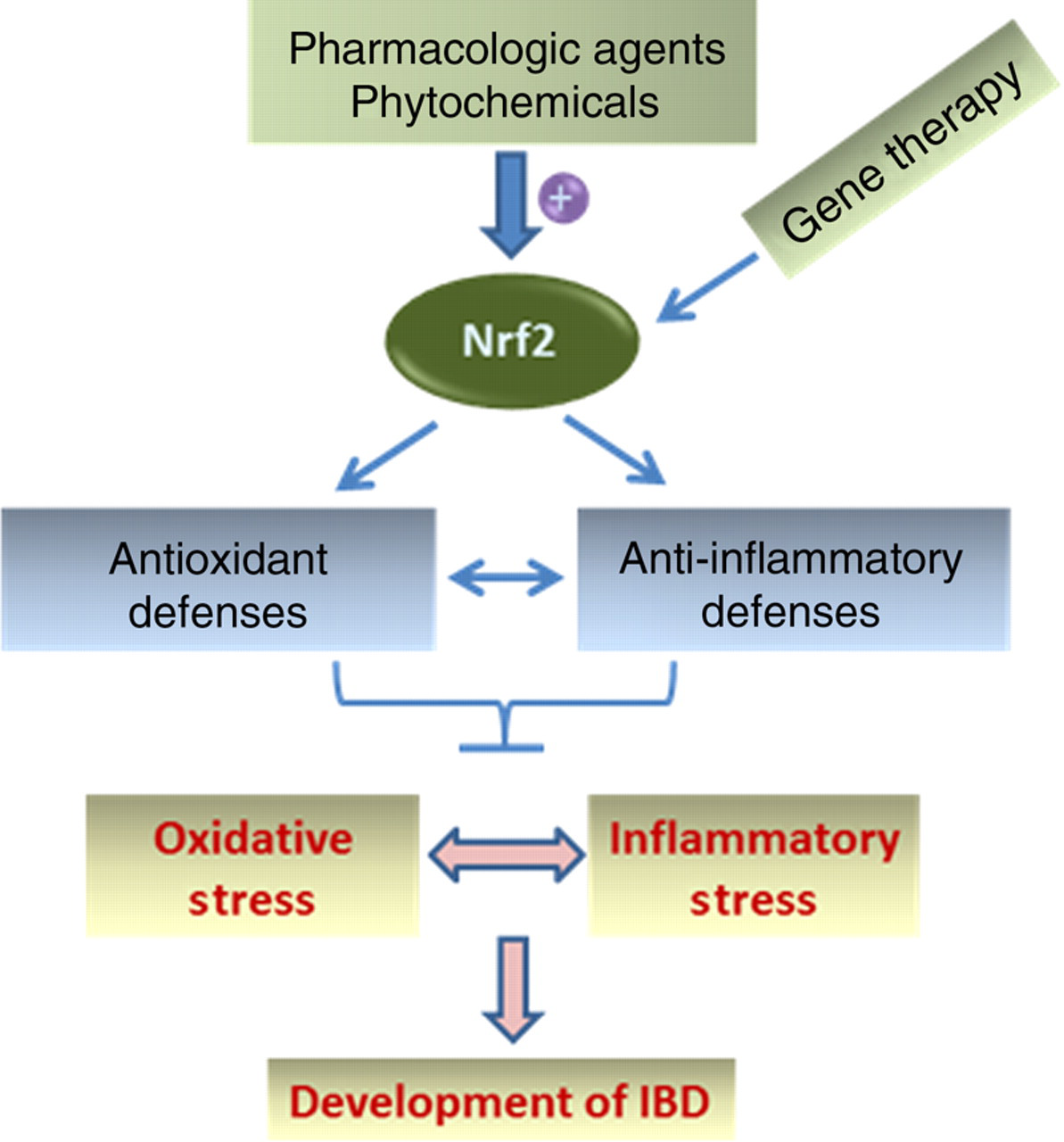
Antioxidant intervention
The involvement of oxidative stress and redox signaling in both experimental IBD and human IBD has spurred an increase of the interest in the development of antioxidant-based modalities for treating this inflammatory disorder in human subjects. In contrast to the large array of antioxidant compounds shown to be protective against IBD in experimental animals, antioxidant compounds shown to be effective in human IBD are relatively few. Nevertheless, multiple pilot studies showed an efficacy of antioxidant-based approaches in treating human IBD. For example, intravenous injection of lecithinized superoxide dismutase was found to be safe and effective in improving the clinical conditions in patients with active ulcerative colitis. 60 This observation is also consistent with the finding in experimental animals that administration of superoxide dismutase suppressed experimental colitis. 23,24 In a randomized, placebo-controlled pilot trial, addition of N-acetylcysteine to conventional mesalamine therapy was reported to increase the remission rate in patients with ulcerative colitis. 61 The dietary phenolic compound curcumin was also shown to ameliorate the disease severity in active ulcerative colitis and reduce clinical relapse in patients with quiescent ulcerative colitis. 62
Induction of colitis by hydrogen peroxide enemas
Direct administration of ROS/RNS to the colon of experimental animals caused acute and chronic colitis. Similarly, decades ago, the use of hydrogen peroxide enemas for the evacuation of fecal impactions was found to be associated with the development of rectal bleeding and colitis in humans. 63 This finding further strengthens a causal role of oxidative stress in the genesis of human IBD.
Summary of the oxidative stress and redox signaling mechanisms in humans
Accumulating evidence from both observational studies and interventional trials suggests a potential causative involvement of oxidative stress in the development of human IBD. Although the preliminary findings with antioxidant intervention are promising and supportive of the oxidative stress mechanism of human IBD, the exact value of these antioxidant compounds, as well as other antioxidant-based modalities in treating human IBD, awaits further investigation in large-scale, well-designed clinical trials. Future clinical studies should not only assess the efficacy of the antioxidant compounds alone, but also determine the combined effects of antioxidants and conventional drugs (e.g. mesalamine, glucocorticosteroids and other immunosuppressive agents). In addition to the use of individual antioxidants, the clinical benefits of antioxidant cocktails should be evaluated, as different antioxidants may target different ROS/RNS pathways that together lead to oxidative gut injury. It is important to mention that future clinical studies should incorporate the indices of oxidative stress and endogenous antioxidant status and determine the correlation between antioxidant therapy, attenuation of oxidative stress and improvement of clinical conditions. Such studies will help establish the causative involvement of oxidative stress in disease pathophysiology and lead to the development of more effective antioxidant-based therapies for human IBD. Lastly, it is important to keep in mind that ROS/RNS are also useful species, playing important physiological roles, especially in innate immunity against pathogenic microorganisms encountered in the gut. In this regard, future antioxidant-based modalities should be designed to reduce the overt oxidative gut tissue injury without significantly compromising the physiological activities of ROS/RNS. Along this line of research, additional studies are warranted to further elucidate the potential differential roles of the individual ROS/RNS in both IBD and innate immunity in animal models as well as human subjects.
Footnotes
ACKNOWLEDGEMENTS
YRL is supported in part by a grant from AICR and NIH HL93557, and ZH is supported in part by NIH DK81905.