
Abstract
Chronic low-grade inflammation is crucial for the development of insulin resistance and type 2 diabetes mellitus (T2DM), and immunocompetent cells, such as T-cells, B-cells, mast cells and macrophages, regulate the pathogenesis of T2DM. However, little is known about the role of natural killer (NK) and natural killer T (NKT) cells in the pathogenic process of T2DM. A total of 16 patients with new onset T2DM and nine healthy subjects were recruited, and the frequency of peripheral blood activated and inhibitory NK and NKT cells in individual subjects was determined by flow cytometry. The frequency of spontaneous and inducible interferon gamma (IFN-γ) and CD107a+ NK cells was further examined, and the potential association of the frequency of NK cells with clinical measures was analyzed. While there was no significant difference in the frequency of peripheral blood NK and NKT cells between patients and controls, the frequency of NKG2D+ NK and NKT cells in patients was significantly higher than those in the controls (P = 0.011). In contrast, the frequency of NKG2A+ and KIR2DL3+ inhibitory NK and NKT cells in patients was significantly lower than those in the controls (P = 0.002, P < 0.0001, respectively). Furthermore, the frequencies of NKG2D+ NK cells were correlated significantly with the values of body mass index in patients. Moreover, the frequencies of spontaneous and inducible CD107a+, but not IFN-γ-secreting, NK cells in patients were significantly higher than those in the controls (P < 0.004, P < 0.0001). Our data indicated that a higher frequency of activated NK cells may participate in the obesity-related chronic inflammation involved in the pathogenesis of T2DM.
Keywords
Introduction
Type 2 diabetes mellitus (T2DM) is characterized by insulin resistance and accounts for the majority of patients with diabetes. 1 During the pathogenesis of T2DM, low-grade inflammation and oxidative stress can be toxic to the pancreatic beta cells, leading to insufficiency of insulin production and exacerbating insulin resistance. However, the pathogenic process of T2DM is not fully understood. Recent studies have shown that immunocompetent cells, such as T-cells, B-cells, mast cells and macrophages, can infiltrate into adipose tissues and contribute to the process of adipose inflammation and insulin resistance. 2,3 In contrast, regulatory T-cells (Tregs) can negatively regulate the chronic inflammation and inhibit the development and progression of obesity and T2DM in mice. 4 However, little is known concerning the function and role of other types of immunocompetent cells in the development and progression of obesity and T2DM.
Natural killer (CD3−CD56+ NK) cells are important components of the innate system and they have the ability to both directly kill target cells and to interact with antigen-presenting cells as well as with T-cells. 5–7 Natural killer T (CD3+CD56+ NKT) cells are a subset of T-cells. NKT cells have limited types of T-cell receptor and recognize lipids and glycolipids in a CD1-dependent manner. NK and NKT cells can be activated and express activating receptors, such as NKG2D and natural cytotoxicity receptors NKp30, NKp44, NKp46 and others. 8 In addition, activated NK and NKT cells can produce inflammatory cytokines, such as interferon gamma (IFN-γ), and degranulate granzyme and other cytotoxic factors, mediating cytotoxicity against target cells. In contrast, NK and NKT cells can express inhibitory receptors, including the lectin-like CD94-NKG2A receptors and the killer cell immunoglobulin-like receptors (KIRs), such as KIR3DL1 and KIR2DL3. 9,10 Engagement of inhibitory receptors by human leukocyte antigen class I molecules initiates an inhibitory signal and inhibits the cytotoxicity of NK and NKT cells against the target cells, particularly for inhibitory NKT cells. Indeed, dysfunction or deficiency of NKT cells has been associated with the development of autoimmune diseases. A recent study shows that a higher frequency of NK cells and a higher NK activity were detected in patients with T2DM. 11 However, the different subsets of NK and NKT cells and what their activation status could be in patients with T2DM, and whether their frequency and function could be associated with the development and severity of T2DM have not yet been explored.
In this study, a group of patients with new onset of T2DM and a group of healthy control subjects were recruited. The frequency and function of peripheral NK and NKT cells in individual subjects were examined to determine the potential association of the frequency of different subsets of NK and NKT cells with the development of T2DM in Chinese patients. Our data indicated that, although there was no significant difference in the frequency of peripheral NK and NKT cells between T2DM patients and controls, a higher frequency of activating NK and NKT cells, but a lower frequency of inhibitory NK and NKT cells, was detected in patients with T2DM. Furthermore, the percentages of NKG2D+ NK cells were positively correlated with values of body mass index (BMI). More importantly, stimulation with K562 cells in vitro induced the degranulation of a significantly higher frequency of NK cells from patients with T2DM. These novel findings suggest that a higher frequency of activating NK cells may participate in the pathogenic process of T2DM in Chinese patients.
Materials and methods
Study subjects
A total of 16 patients with newly diagnosed T2DM were recruited to the inpatient service of the First Hospital of Jilin University in Changchun, China. Individual patients with T2DM were diagnosed, according to the diagnosis standards (fasting plasma glucose [FPG]>120 mg/dL and hemoglobin A1C (HbA1c) > 7%) of T2DM of the American Diabetes Association (ADA) 2011. 12 These patients met the criteria: (i) male or female patients between 20 and 70 years of age; (ii) diagnosis of T2DM within one month without diabetic ketoacidosis; (iii) BMI ≥ 25 kg/m2; (iv) Pc-peptide (mmol/L) post-Sustacal (60 min) >0.4 mmol/L, Pc-peptide (mmol/L) post-Sustacal (120 min) >0.8 mmol/L; (v) having no formal treatment for diabetes; and (vi) having no indications of other chronic diseases. Furthermore, individual participants with chronic respiratory disease, smoking habit, clinical history of neuromuscular disease, narcolepsy, stroke, transient ischemic attack, chronic heart failure, craniofacial abnormalities, alcoholic abuse or use of sedative drugs, current pregnancy, other endocrinological diseases and recent infection were excluded. There were nine healthy volunteers participating in this study. Written informed consent was obtained from individual participants and the experimental protocol was approved by the Institute Review Board of the First Hospital of Jilin University.
Clinical measurements
Individuals were subjected to physical examination and the age, BMI and waist circumference (WC) of each participant were measured. Individuals fasted overnight and their venous blood samples were obtained. The concentrations of FPG and serum insulin C-peptide and HbA1c were measured by the glucose oxidase method using a Hitachi 7600-020 automatic analyzer (Hitachi High-Technologies, Tokyo, Japan), radioimmunology (SR 300; STRATEC, Birkenfeld, Germany) and by immunoturbidimetry, respectively. Furthermore, their diabetic status, fasting lipids, and hematological and biochemical parameters were examined. In addition, individual subjects were subjected to a standard oral glucose tolerance test for the concentrations of C-peptide 30, 60, 120 and 180 min post-Sustacal stimulation. Insulin resistance of individuals was assessed by the homeostatic model assessment (HOMA; insulin resistance index = [FPG (mmol/L) × insulin (μU/mL)]/22.5).
Blood samples
Venous blood was collected after an overnight fast and analyzed on the day of collection. Routine laboratory analyses were performed by standard techniques. Ten milliliters of venous blood were collected in heparinized tubes for lymphocyte profiling.
Flow cytometry analysis
Individual venous samples were subjected to flow cytometry analysis of peripheral lymphocytes using specific antibodies. Briefly, 100 μL of blood samples were incubated in a mixture of fluorescein isothiocyanate (FITC)-conjugated anti-CD3, allophycocyanin (APC)-conjugated anti-CD56, peridinin chlorophyll protein (PerCP)-conjugated anti-CD16 and phycoerythrin (PE)-conjugated anti-NKG2D, anti-NKG2C, anti-NKp30, anti-NKp44, anti-NKG2A, anti-KIR2DL3, anti-CD158a and anti-CD158b (BD Biosciences, San Diego, CA, USA) for 20–30 min at room temperature, respectively. The mouse IgG1 and IgG2a were used as negative controls. The remaining erythrocytes were lysed using BD FACS lysing Solution 2 (BD Biosciences), and the frequency of the different subsets of NK and NKT cells was determined by flow cytometry using a FACSCalibur (BD Biosciences) and FlowJo software (v5.7.2; TreeStar, Ashland, OR, USA).
IFN-γ production
For the detection of IFN-γ secreted by NK cells, peripheral blood mononuclear cells (PBMCs) were isolated from individual heparinized blood samples by density-gradient centrifugation using Ficoll-Paque Plus (Amersham Biosciences, Buckinghamshire, UK). PBMCs (106 cells/well) were stimulated with 50 ng/mL phorbol myristate acetate (Sigma-Aldrich, St Louis, MO, USA) and 1.0 μg/mL of ionomycin (Sigma-Aldrich) in complete RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) for two hours at 37°C in 5% CO2 and exposed to Brefeldin A (GolgiPlug; BD Biosciences) for an additional five hours. Subsequently, the unadherent cells were harvested, washed with cold phosphate-buffered saline (PBS) and stained with FITC-anti-CD56 and APC-anti-CD3, followed by being fixed with 4% paraformaldehyde (30 min/room temperature) and then permeabilized with 0.5% saponin in 10% fetal bovine serum in PBS (30 min/room temperature). After washes, the cells were stained with PE-anti-IFN-γ and subjected to flow cytometry analysis.
The degranulation of NK cells
The CD107a degranulation is widely used for evaluating the potentials of NK cell cytotoxicity. 13 The isolated PBMCs (106 cells/well) were co-cultured in duplicate with K562 cells at a ratio of 10:1 of effector to target (E:T) in RPMI-1640 medium for one hour in the presence of anti-CD107a or control IgG2a, and exposed to 2 μm monensin (GolgiStop; BD Biosciences) for an additional five hours. The PBMCs alone served as negative controls. Subsequently, the cells at 106/tube were stained in duplicate with FITC-anti-CD3 and APC-anti-CD56 at room temperature for 30 min, respectively. After washing with PBS (containing 1% FCS and 2.5% paraformaldehyde), the frequency of CD107a+ CD3−CD56+ NK cells was determined by flow cytometry analysis using a FACS Calibur, and at least 10,000 CD56+ events per sample were analyzed.
Statistical analysis
All data are expressed as individual values, median and range of each group of subjects. Multiple comparisons between two groups were analyzed by the Kruskal–Wallis H non-parametric test. Correlations between variables were evaluated by the Spearman's rank correlation test using SPSS 19.0 for Windows (SPSS, Inc, Chicago, IL, USA). A two-sided P value of <0.05 was considered statistically significant.
Results
Higher frequencies of activated NK and NKT cells, but lower frequencies of inhibitory NK and NKT cells, are associated with the development of T2DM
The demographic and characteristics of subjects
BMI, body mass index; WC, waist circumference; HbA1C, hemoglobin A1C; N/A, not applicable; T2DM, type 2 diabetes mellitus
Characterization of CD3−CD56+ NK and CD3+CD56+ NKT cells revealed that there was no significant difference in the frequency of peripheral NK, NKT, NKG2C+, NKp30+, NKp44+, CD158a+, CD158b+ NK and NKT cells between the patient and control groups (data not shown). There was no significant difference in the frequency of CD3−CD16+ NK and CD3+CD16+ NKT cells between these two groups of subjects (data not shown). The frequency of NK and NKT cells in patients was not significantly associated with the clinical characteristics (data not shown). However, we found that the frequency of activated NKG2D+ NK and NKT cells in the patients was significantly higher than that in the controls (Figures 1a–c). In contrast, the frequency of inhibitory KIR2DL3+ and NKG2A+ NK and NKT cells, but not other inhibitory NK and NKT cells, in patients was significantly lower than in the controls (Figures 1d–i). These data indicated that a higher frequency of activated NK and NKT cells, but a lower frequency of inhibitory NK and NKT cells, was associated with the development of T2DM in Chinese patients.
Characterization of different subsets of naturall killer (NK) and NKT cells. Peripheral blood samples (100 μL) of individual participants were stained in duplicate with FITC-anti-CD3, APC-anti-CD56, PerCP-anti-CD16 and PE-anti-NKG2D, anti-NKG2C, anti-NKp30, anti-NKp44, anti-NKG2A, anti-KIR2DL3, anti-CD158a and anti-CD158b or control mouse IgG1 and IgG2a for 20–30 min at room temperature, respectively. After lyzing the remaining erythrocytes, the frequency of different subsets of NK and NKT cells was determined by flow cytometry. The cells were gated on APC-anti-CD56+ and at least 10,000 CD56+ events were analyzed. Data shown are representative charts from different groups of subjects and the percentages of specific subsets of NK and NKT cells in individual subjects from five separate experiments. (a) Flow cytometry analysis of NKG2D+ NK and NKT cells; (b) quantitative analysis of the frequency of NKG2D+ NK cells; (c) quantitative analysis of the frequency of NKG2D+ NKT cells; (d) flow cytometry analysis of the frequency of NKG2A+ NK and NKT cells; (e) quantitative analysis of the frequency of NKG2A+ NK cells; (f) quantitative analysis of the frequency of NKG2A+ NKT cells; (g) flow cytometry analysis of the frequency of KIR2DL3+ NK and NKT cells; (h) quantitative analysis of the frequency of KIR2DL3+ NK cells; and (i) quantitative analysis of the frequency of KIR2DL3+ NKT cells. FITC, fluorescein isothiocyanate; APC, allophycocyanin; PerCP, peridinin chlorophyll protein; PE, phycoerythrin; HC, healthy controls; T2DM, Type 2 diabetes mellitus. (A color version of this figure is available in the online journal)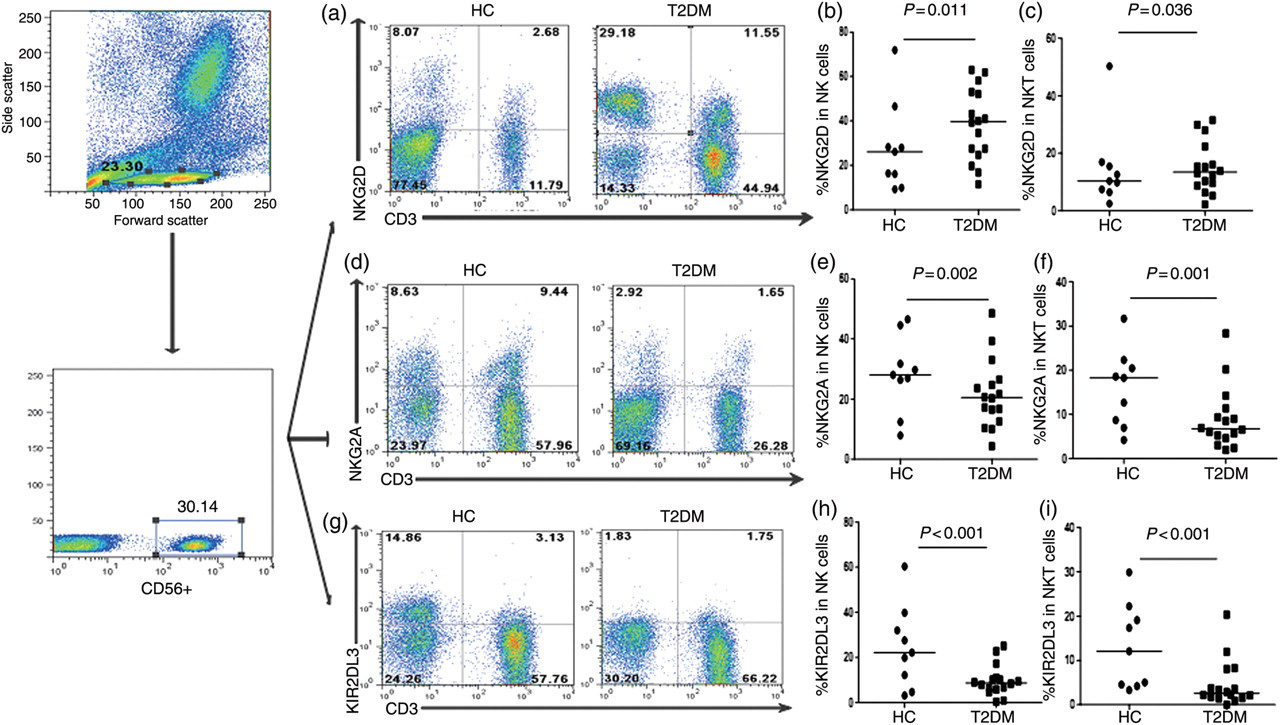
The frequencies of NKG2D+ NK cells are correlated with the values of BMI in patients with T2DM
Further analysis of the role of activated NKG2D+ NK and NKT cells in the pathogenesis of T2DM indicated that the frequencies of NKG2D+ NK cells, but not NKT cells, were significantly correlated with the values of BMI in patients with T2DM (Figure 2). However, the frequency of NKG2D+ NK cells was not associated with other measures tested in those patients. Interestingly, we did not observe any significant association of the frequency of inhibitory NK and NKT cells with any of the clinical measures tested in those patients.
The frequencies of NKG2D+ natural killer (NK) cells are significantly correlated with the values of body mass index (BMI) in patients with new onset of type 2 diabetes mellitus. The correlation between the frequencies of NK cells and NKT cells and the values of BMI in patients was determined by the Spearman's rank correlation test. Data shown are individual values of those patients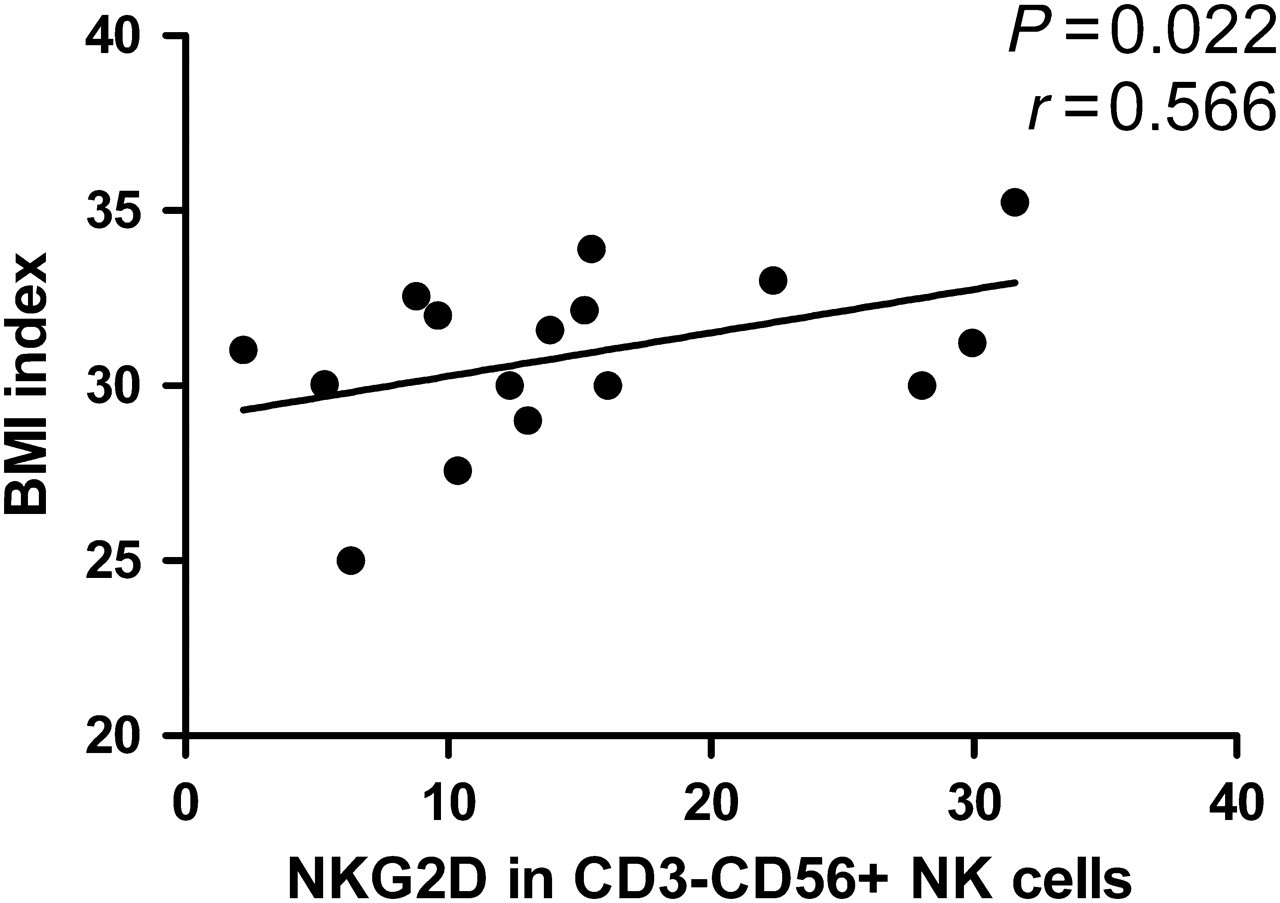
Functional analysis of activated NK cells in patients with T2DM
Activated NK and NKT cells can produce proinflammatory cytokines, such as IFN-γ, and have potent cytotoxicity against target cells. Next, we examined the sensitivity of NK cells to stimulation in patients with T2DM. As shown in Figures 3a and b, a few NK cells from both patients and controls spontaneously produced IFN-γ and a low frequency of NK cells from most patients produced IFN-γ, except that a high frequency of IFN-γ-secreting NK cells was detected in a few patients. As a result, there was no significant difference in the frequency of spontaneous and inducible IFN-γ-secreting NK cells between the patients and controls.
Characterization of spontaneous and inducible natural killer (NK) cell activation. The isolated peripheral blood mononuclear cells (PBMCs) were stimulated with, or without, phorbol myristate acetate (PMA)/lonomycin for seven hours in the presence of monensin for the last five hours and the cells were stained with FITC-anti-CD3 and APC-anti-CD56. After being fixed and permeabilized, the cells were stained with PE-anti-IFN-γ. The frequency of IFN-γ-secreting NK cells was determined by flow cytometry analysis and at least 10,000 events per sample were analyzed. In addition, the isolated PBMCs were cultured alone for testing spontaneous activation or co-cultured in duplicate with K562 cells in the presence of anti-CD107a or control IgG2a for six hours. Subsequently, the cells were stained with FITC-anti-CD3 and APC-anti-CD56. Then, the frequency of CD107a+CD3−CD56+ NK cells was analyzed. Data shown are representative charts from healthy controls and T2DM patients and expressed as individual values of the percentage of activated NK cells from five separate experiments. (a) Flow cytometry analysis of the frequency of IFN-γ-secreting NK cells; (b) quantitative analysis of the percentage of IFN-γ-secreting NK cells; (c) flow cytometry analysis of the frequency of CD107a+ NK cells; and (d) quantitative analysis of the percentage of CD107a+ NK cells. FITC, fluorescein isothiocyanate; APC, allophycocyanin; PE, peridinin chlorophyll protein; IFN-γ, interferon gamma. (A color version of this figure is available in the online journal)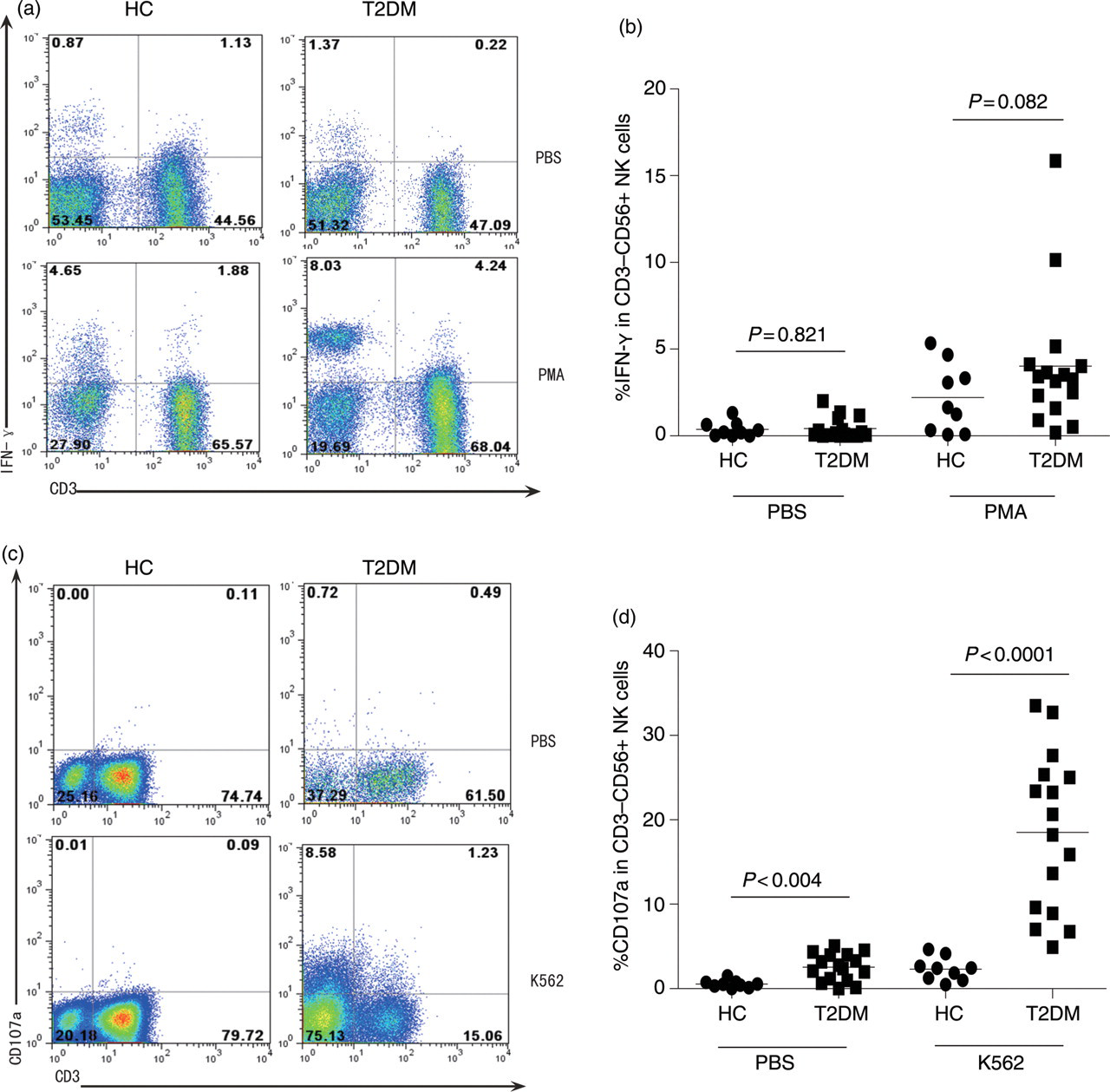
Characterization of NK cell degranulation indicated that the frequency of spontaneous CD107a+ NK cells in patients with T2DM was significantly higher than that in controls. Following stimulation with the target K562 cells, the frequency of CD107a+ NK cells in controls only slightly increased. In contrast, the frequency of CD107a+ NK cells after K562 stimulation dramatically increased in patients and was significantly higher in patients than those in the controls (Figures 3c and d). These data clearly indicated that spontaneously activated NK cells existed in patients with T2DM.
Discussion
Chronic inflammation has been associated with the development of insulin resistance and T2DM. 14–19 Aberration of innate immunity, such as NK and NKT cells, plays an important role in the development of inflammatory diseases, including coronary artery disease and chronic obstructive pulmonary disease. 20–22 However, the role of NK and NKT cells in the development of insulin resistance and T2DM is still unclear. In this study, we first characterized the frequency of different subsets of peripheral blood NK and NKT cells in patients with new onset T2DM and healthy control subjects. We found that there was no significant difference in the frequency of NK and NKT cells between T2DM patients and control subjects. However, the frequency of NKG2D+ activated NK and NKT cells in T2DM patients was significantly higher than those in healthy subjects, while the frequency of NKG2A+ and KIR2DL3+ inhibitory NK and NKT cells in T2DM patients was significantly lower than those in healthy subjects. These data indicated that a higher frequency of activated NK and NKT cells and a lower frequency of inhibitory NK and NKT cells were associated with the development of T2DM in this population. The low frequency of inhibitory KIR2DL3+ and NKG2A+ NK and NKT cells in patients with T2DM may reduce the threshold for the activation of NK and NKT cells contributing to the inflammatory process. Hence, the balance of activated and inhibitory NK and NKT cells is crucial for regulating chronic inflammation involved in the pathogenic process of T2DM. Our data are partially consistent with a previous observation of higher NK and NKT cell activity in T2DM patients. 11 However, our data were not consistent with the previous findings of increased frequency of NK cells in T2DM patients. 11 The difference may stem from variable populations of patients as they studied patients with a mean of 7.4 years of T2DM rather than the new onset T2DM patients we studied. Previous studies have indicated that NK cells regulate peripheral tolerance and inflammatory responses as well as the pathogenesis of T1DM. 23–28 However, to the best of our knowledge, this is the first report on higher frequency of activated NK and NKT cells and lower frequency of inhibitory NK and NKT cells in T2DM patients.
Obesity is a risk factor for the development of metabolic syndrome-related T2DM, hypertension and non-alcoholic fat liver diseases, as well as other macrovascular and microvascular diseases. Interestingly, we found that the frequency of NKG2D+ NK cells in patients with new onset T2DM was correlated positively with the values of BMI. Notably, chronic inflammation in adipose tissues is critical for the development of obesity and T2DM 29 and a previous study has shown a higher frequency of NKG2D+ NK cells in patients with latent autoimmune diabetes in adults (LADA). 27 Given that both T2DM and LADA share similar clinical features at the early stage of the pathogenic process, it is possible that NKG2D+ NK cells participate in the inflammatory process in the adipose tissues, contributing to the pathogenesis of both diseases. Apparently, measurement of NKG2D+ NK cells may be valuable for the early detection of obesity-related inflammation in those at high risk of developing T2DM.
Interestingly, we found that the frequency of spontaneous and inducible IFN-γ-secreting NK cells in patients with T2DM was not significantly different from those in the healthy controls. It is possible that NK cells participate in the pathogenic process of T2DM by secreting other proinflammatory cytokines, such as IL-15 and IL-12. 30,31 Alternatively, NK cells may predominately degranulate, contributing to the pathogenic process of T2DM. Indeed, we found a high frequency of CD107a+ NK cells in patients with T2DM. Notably, activated NK cells can secrete high levels of proinflammatory cytokines, such as IL-12, IL-15, IL-18, IL-2 and CCL5 and release perforin and proteases as well as other enzymes by degranulation. Conceivably, activated NK cells may, through these proinflammatory factors, contribute to the pathogenesis of T2DM.
In conclusion, our data indicated a higher frequency of NKG2D+ NK and NKT cells, but a lower frequency of NKG2A+ and KIR2DL3+ NK and NKT cells in patients with new onset T2DM, and the frequency of NKG2D+ NK cells was correlated with the values of BMI in T2DM patients. Furthermore, the higher frequency of activated NK cells was associated with higher levels of NK cell degranulation in patients with T2DM. Apparently, our findings suggest that activated NK cells may participate in the obesity-related chronic inflammation involved in the pathogenic process of T2DM. We recognize that our studies had limitations of a small sample size, one time of measurement and a lack of longitudinal follow-up, as well as only a few proinflammatory factors being tested. We are interested in further investigating the role of NK and NKT cells in the pathogenic process of T2DM and the potential diagnostic and prognostic values of detecting activated and inhibitory NK and NKT cells in patients with T2DM. If confirmed, our findings may provide new insights into understanding the pathogenesis of T2DM and aid in developing new measures for early diagnosis and prognosis of obesity-related chronic inflammation in humans.
Footnotes
ACKNOWLEDGEMENTS
We thank Medjaden Bioscience Limited for assisting in the preparation of this manuscript. This work is supported by grants from the National Natural Science Foundation of China (No. 30972610), Jilin Province Science and Technology Agency (No. 200705128), the Chinese Medical Science and Technology Projects of Jilin Province (08sys-086), the Health Department Research Projects in Jilin Province (2009Z054) and Cutting-edge Science and Interdisciplinary Innovation Projects of Jilin University.