
Abstract
Human idiopathic pulmonary fibrosis (IPF) is a disease with unknown etiology and poor prognosis in which patients present a decrease in functional exercise tolerance and quality of life. At present, no treatment which can improve the prognosis of this disease is available. Many biomarkers of pulmonary fibrosis have been studied, and surfactant protein A (SP-A) expression is considered a specific marker of lung disease. This study aimed to investigate the influence of exercise training on exercise endurance capacity and murine-lung lesions induced by bleomycin (BLM). Thirty-four male Balb/c mice were subdivided into four groups: control sedentary (C-SED), bleomycin-treated sedentary (BLM-SED), control exercised (C-EXE) and bleomycin-treated exercised (BLM-EXE). Mice received 6.25 U/kg of BLM or saline via intratracheal instillation. After adaptation in a swimming pool, the animals started training one hour per day, with 60% of maximum load obtained in exercise endurance capacity assessment, five days/week for four weeks. The lungs were collected 48 h after the second endurance capacity assessment, fixed in buffered formalin and embedded in paraffin. Sections were analyzed using histochemical and immunohistochemical reactions for digital morphometry of pulmonary fibrosis, type I collagen, SP-A and type II pneumocytes (PII). The exercise endurance capacity of groups C-EXE (9.20 ± 0.81 min) and BLM-EXE (8.40 ± 0.82 min) increased significantly when compared with groups C-SED (5.84 ± 0.4 min) and BLM-SED (5.67 ± 0.60 min). The amounts of connective tissue, type I collagen, PII and SP-A increased significantly in the BLM-SED group. Exercise training significantly attenuated this response as observed in the BLM-EXE group. The present study shows that exercise training can prevent the decline of exercise endurance capacity and attenuate the progression of IPF.
Keywords
Introduction
Human idiopathic pulmonary fibrosis (IPF) is a disease with unknown etiology and poor prognosis. 1 The diagnosis of IPF is generally based on high-resolution computerized tomography scanning and invasive procedures. 2 Its incidence has been reported as 6–42 per 100,000 persons, but increases with age to more than 200 per 100,000 persons in those above 75 y of age.
Several factors have been associated with initiation and/or progression of IPF. Oxidative stress contributes to the pathology of bleomycin (BLM)-treated mice, a well-described model of pulmonary fibrosis. 3 The use of this model is based on the fact that fibrosis is one of the major adverse drug effects of BLM in human cancer therapy. Despite the fact that the BLM model has limitations in regard to understanding the progressive nature of human IPF, 4 the undisputed qualities and similarities in histological alterations observed in this model make it relevant and highly informative.
Many biomarkers of pulmonary fibrosis have been studied, with surfactant protein A (SP-A) being one of the most promising. In IPF patients, its concentration is decreased in bronchoalveolar lavage (BAL) and increased in plasma when compared with healthy subjects. 5,6
At present, no available treatment can improve the prognosis of this disease. 1 In addition to identifying additional agents and key dysregulated pathways that interfere in disease progression, we should also be attempting to develop and/or apply targeted therapies to treat or prevent this severe disease. 7 IPF patients present poor functional exercise tolerance and quality of life. Pulmonary re-habilitation has been applied successfully to patients with other chronic lung diseases such as interstitial diseases. However, the number of articles concerning the effects of pulmonary re-habilitation is highly limited in patients with lung fibrosis, 8 and they usually evaluate only functional changes.
Recently, we evaluated the aerobic performance of rats with lung fibrosis induced by paraquat. 9 We observed that the paraquat group presented a decrease of aerobic performance, mechanical efficiency and metabolic cost during acute progressive exercise on a treadmill compared with the control group. Additionally, a close negative correlation was observed between physical performance and the fractional area of collapsed alveoli. Until now, no study has evaluated the effects of aerobic exercise on the histopathological changes of lung parenchyma in a murine model of IPF. Thus, the present study investigates the influence of exercise training on exercise endurance capacity in mice treated with BLM. Additionally, in order to investigate the effects of aerobic exercise on pulmonary injury induced by BLM, immunohistochemical and morphometric analyses of pulmonary fibrosis, type I collagen and SP-A were performed in lung parenchyma.
Materials and methods
Animals
Thirty-four, 20-week-old, male Balb/c mice were subdivided into four groups: control sedentary (C-SED, n = 6), bleomycin-treated sedentary (BLM-SED, n = 8), control exercised (C-EXE, n = 6) and bleomycin-treated exercised (BLM-EXE, n = 8). The study design is described in Figure 1.
Study design. C-SED, control sedentary group (n = 6); C-EXE, control exercised group (n = 6); BLM-SED, bleomycin-treated sedentary group (n = 8); BLM-EXE, bleomycin-treated exercised group (n = 8)
Pulmonary fibrosis induction
After anesthesia (ketamine 5%, 200 mg/kg and xylazine 2%, 10 mg/kg dissolved in 2 mL of saline solution 0.9%, intraperitonial), 6.25 U/kg of BLM dissolved in saline was delivered via an intratracheal route (Bleomycin Sulfate, Meizle Biopharma, Barueri, SP, Brazil). The control groups received the same volume of saline (40 μL).
Exercise endurance capacity evaluation
The assessment of the exercise endurance capacity was carried out according to Almeida et al. 10 , before and after the physical training. Fourteen days after the intratracheal instillation, all animals were adapted to the pool water (20 cm deep and temperature kept at 31 ± 1°C), for three minutes per day, for five days, with no overload. Mice were then submitted to a progressive load test, which consisted of an increasing workload corresponding to 2% of body weight added every three minutes until exhaustion. Exhaustion was defined when mice failed to rise to the surface of the water to breathe within a seven-second period. 11
Adaptation protocol to physical training in swimming pool
The animals swam for 10, 20 and 40 min/day for five days in the first, second and third weeks, respectively, from 23 to 44 days after BLM instillation. 12
Physical training protocol in swimming pool
Animals from the C-EXE and BLM-EXE groups were trained for 60 min/day, five days/week, during four weeks (44–72 days), at 60% of the maximum load, performing a moderate-intensity physical activity. 12
Necropsy and histopathological analysis of lungs
Removal of lungs was performed 74 days after BLM instillation as described in our previous work. 9 After transection of abdominal aorta, the lungs were immediately collected, washed with NaCl 0.9% and fixed in 10% buffered formalin. Lung fragments were processed to obtain 4 μm thick sections, which were then stained with hematoxylin and eosin and Gomori's trichrome for histopathological and morphometric analyses of fibrosis. Other sections of these fragments were used to perform immunohistochemical localization of anti-type I collagen and anti-SP-A.
Immunohistochemical reaction for type I collagen and SP-A
Lung sections were dewaxed, hydrated and washed in phosphate-buffered saline (PBS) pH 7.2. Endogenous peroxidase activity was eliminated using H2O2 30% at 3.5% in PBS and the non-specific binding sites were blocked with rabbit serum 1:40. The sections were incubated with anticollagen type I monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) 1:50, followed by incubation with biotinylated IgG (Bethyl Laboratories Inc., Montgomery, AL, USA) and streptavidin peroxidase conjugate (Zymed Laboratories Inc., San Francisco, CA, USA) 1:50. Color was detected with 0.05% diaminobenzidine solution in H2O2 30% at 0.2% and the sections were counterstained with Harris Hematoxylin. As a negative control, the primary antibody was omitted from the staining protocol. Histological sections of normal lungs of mice were used as positive controls. For immunohistochemical identification of SP-A, sections were dewaxed, hydrated and washed in PBS; antigenic retrieval was performed with the Dako Retrieval solution (Dako, Carpinteria, CA, USA) at 100°C for 20 min. Sections were then incubated with anti-SP-A monoclonal serum 1:50 (Santa Cruz Biotechnology), and the subsequent immunolabeling steps as outlined above were performed.
Morphometric analysis of pulmonary fibrosis, type I collagen, SP-A protein and type II pneumocytes
Histological sections of lung parenchyma were stained with Gomori's trichrome, visualized using a 40× objective and 30 random images were digitized through a JVC TK-1270/RGB (Tokyo, Japan) microcamera. The area of each image was 53,333.4 μm2, equaling a total area of 1.6 × 106 μm2 of lung parenchyma analyzed in each case. The fibrosed area was estimated by selecting the green-shaded pixels in the real image, and subsequently creating a binary image for digital processing. Measurements were obtained using the Carl Zeiss KS300 image analyzer software package (Oberkochen, Germany). 13 Using the same method, the area positively immunolabeled for type I collagen or SP-A was estimated by selecting the brown pixels. In the digitalized images of SP-A immunolabeling, a manual count of the type II pneumocytes (PII) was performed using the tools in the KS300 program.
Statistical analysis
The results are expressed as means ± SEM. Comparisons among different groups were assessed by one-way analysis of variance followed by Bonferroni test and two-tailed paired or no paired t-test as appropriate. Pearson's r was used to correlate the different variables. Statistical analyses were performed with the software GraphPad Prism 4 (GraphPad Software, Inc., San Diego, CA, USA). Statistical significance was set at P < 0.05.
Results
Exercise endurance capacity evaluation
Figure 2a shows the first and second evaluations of the exercise endurance capacity of groups C-SED and C-EXE. C-SED had its exercise endurance capacity significantly reduced over time (7.79 ± 0.53 versus 5.84 ± 0.4 min). In contrast, the C-EXE group increased significantly the exercise time between the first (7.19 ± 1.1 min) and the second (9.20 ± 0.81 min) evaluation (Figure 2a).
(a) Comparing the first (1st) and second (2nd) exercise endurance capacity evaluation (minutes) in sedentary (C-SED) and trained (C-EXE) mice. *P < 0.05 regarding the first evaluation. (b) Comparing the 1st and 2nd exercise endurance capacity evaluation (minutes) in sedentary (BLM-SED) and trained (BLM-EXE) mice with bleomycin-induced pulmonary fibrosis. *P < 0.05 regarding the second evaluation. (c) Comparing the second exercise endurance capacity evaluation (minutes) of sedentary and trained mice in the control group (C-SED and C-EXE) and in the bleomycin group (BLM-SED and BLM-EXE). *P < 0.05: regarding the sedentary group. C-SED, control sedentary; C-EXE, control exercised; BLM-SED, bleomycin-treated sedentary; BLM-EXE, bleomycin-treated exercised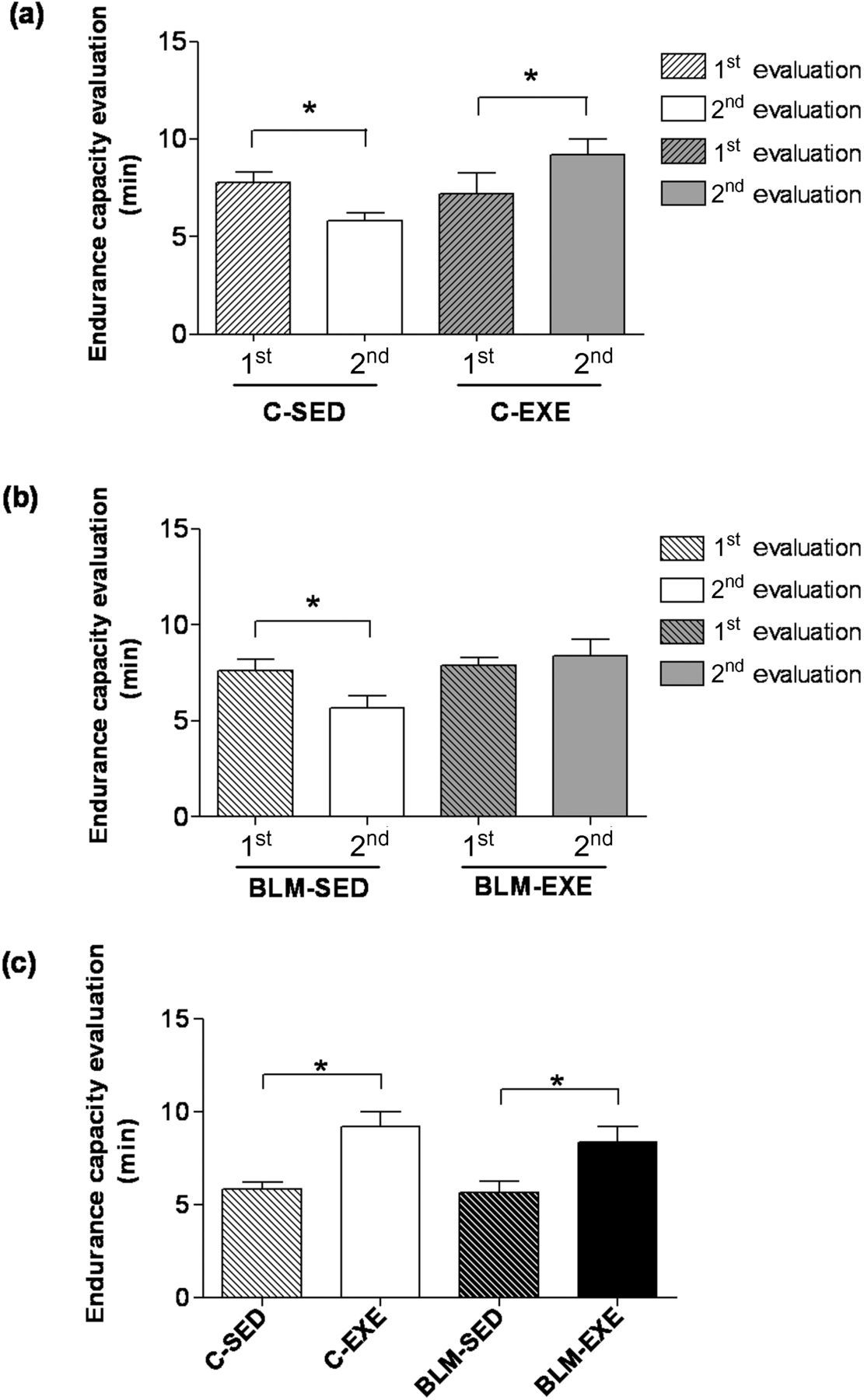
Similar to the C-SED group, sedentary animals with pulmonary fibrosis (BLM-SED) also presented a significant decrease in exercise endurance capacity in the second endurance capacity assessment (7.65 ± 0.57 min versus 5.67 ± 0.60 min) (Figure 2b). No difference was observed between the first and second evaluations, that is, before and after the training in the BLM-EXE group (7.89 ± 0.39 min versus 8.40 ± 0.82 min).
When comparing the groups (Figure 2c), considering only the last exercise endurance capacity evaluation, a significant increase (P < 0.05) was observed in physical conditioning in C-EXE (9.20 ± 0.81 min) and BLM-EXE (8.40 ± 0.82 min) in comparison to C-SED (5.84 ± 0.4 min) and BLM-SED (5.67 ± 0.60 min).
Macroscopic and histopathological analyses of lungs
The lungs of animals in C-SED and C-EXE showed normal macroscopic aspects (Figure 3a). Differently, in BLM-SED, the pleural surface of all animals presented dense, firm and reddish-white areas suggesting fibrosis (Figure 3d). Such areas were not observed in BLM-EXE, except for one animal in which small hemorrhagic foci and greater consistency were found (Figure 3g).
Lungs of mice treated with or without bleomycin, sedentary or submitted to swimming. (a) C-SED group: normal appearance; (b) C-SED group: alveolar septa with normal thickness (arrowheads), respiratory bronchioles (B) and aerated alveoli (A). Bar = 100 μm; (c) Higher magnification of the previous figure showing a detail of the green-stained interalveolar connective tissue (arrowheads). Bar = 30 μm; (d) BLM-SED group: dense, reddish-white areas, suggesting fibrosis (arrows); (e) BLM-SED group: great thickening of the interalveolar septa (arrowheads). Hyperemia (*). Bar = 100 μm; (f) BLM-SED group: strong thickening of the interalveolar interstitium resulting from the accumulation of cells and richly vascularized connective tissue (*). Bar = 30 μm; (g) BLM-EXE group: hemorrhagic areas (arrows); (h) BLM-EXE group: thickening of interalveolar septa (arrows). Aerated respiratory bronchioles (B). Bar = 100 μm; (i) BLM-EXE group: thickening of interalveolar septa (arrowheads). Bar = 30 μm. Gomori's trichrome. C-SED, control sedentary; BLM-SED, bleomycin-treated sedentary; BLM-EXE, bleomycin-treated exercised. (A color version of this figure is available in the online journal)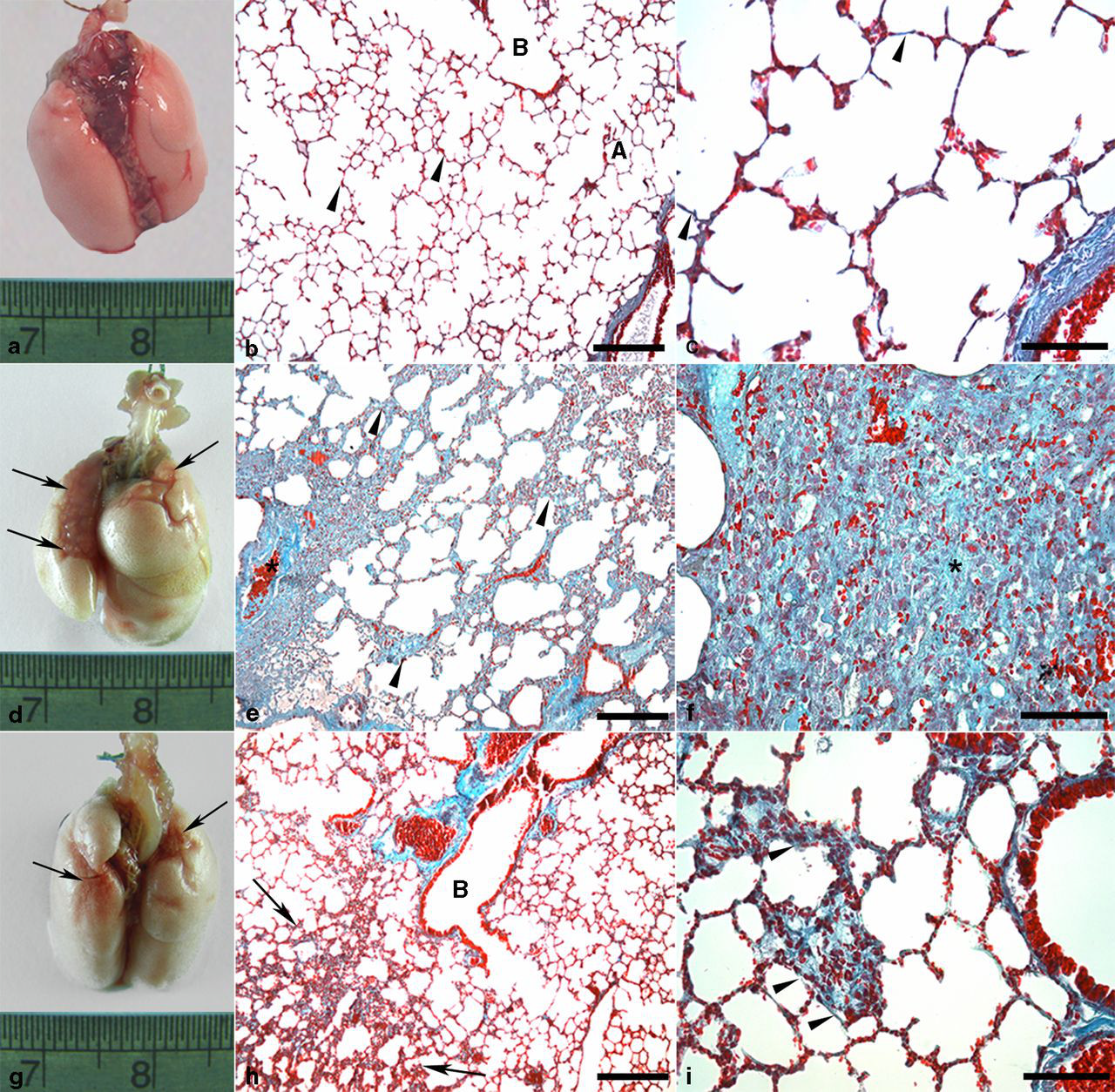
The lung parenchyma of C-SED and C-EXE presented a normal histological aspect (Figures 3b and c). Sedentary animals receiving BLM showed a fibrosing interstitial pneumonitis (Figures 3e, f and 4). The accumulation of inflammatory cells produced a thickening of the alveolar septa and, though less frequently, of interlobular septa. In BLM-EXE, a visible reduction of fibrosis in all areas and the restoration of much of the alveolar air space were observed, when compared with BLM-SED (Figures 3h and i).
Lung parenchyma of sedentary mice treated with bleomycin. (a) Large amount of alveolar macrophages (arrowheads) in the alveolar lumen adjacent to the bronchial wall (arrows). Bronchiolar lumen (L). Bar = 30 μm. (b) Thickening of the alveolar (arrowheads) and interlobular septa (arrows). Aerated alveoli (A). Bar = 100 μm. (c) Necrosis of bronchiolar epithelium (arrows) associated with the presence of alveolar macrophages in contact with the epithelium (arrowheads). Thickening of interalveolar septa (*). Bronchiolar lumen (L). Bar = 30 μm; (d) Bronchial epithelial papillae projected into the lumen resulting in regenerative hyperplasia (arrows). Thickening of the septum (*). Bronchiolar lumen (L). Bar = 30 μm. (e) Areas with collapsed alveoli showing the presence of atelectasis (arrows). Bar = 100 μm. (f) Detail of previous figure showing collapsed alveoli (arrows), alveolar macrophages (arrowheads) and interalveolar thickening mainly due to fibrosis (*). Bar = 30 μm; (a, b, e, f): Gomori's trichrome. (c, d): Hematoxylin and eosin. (A color version of this figure is available in the online journal)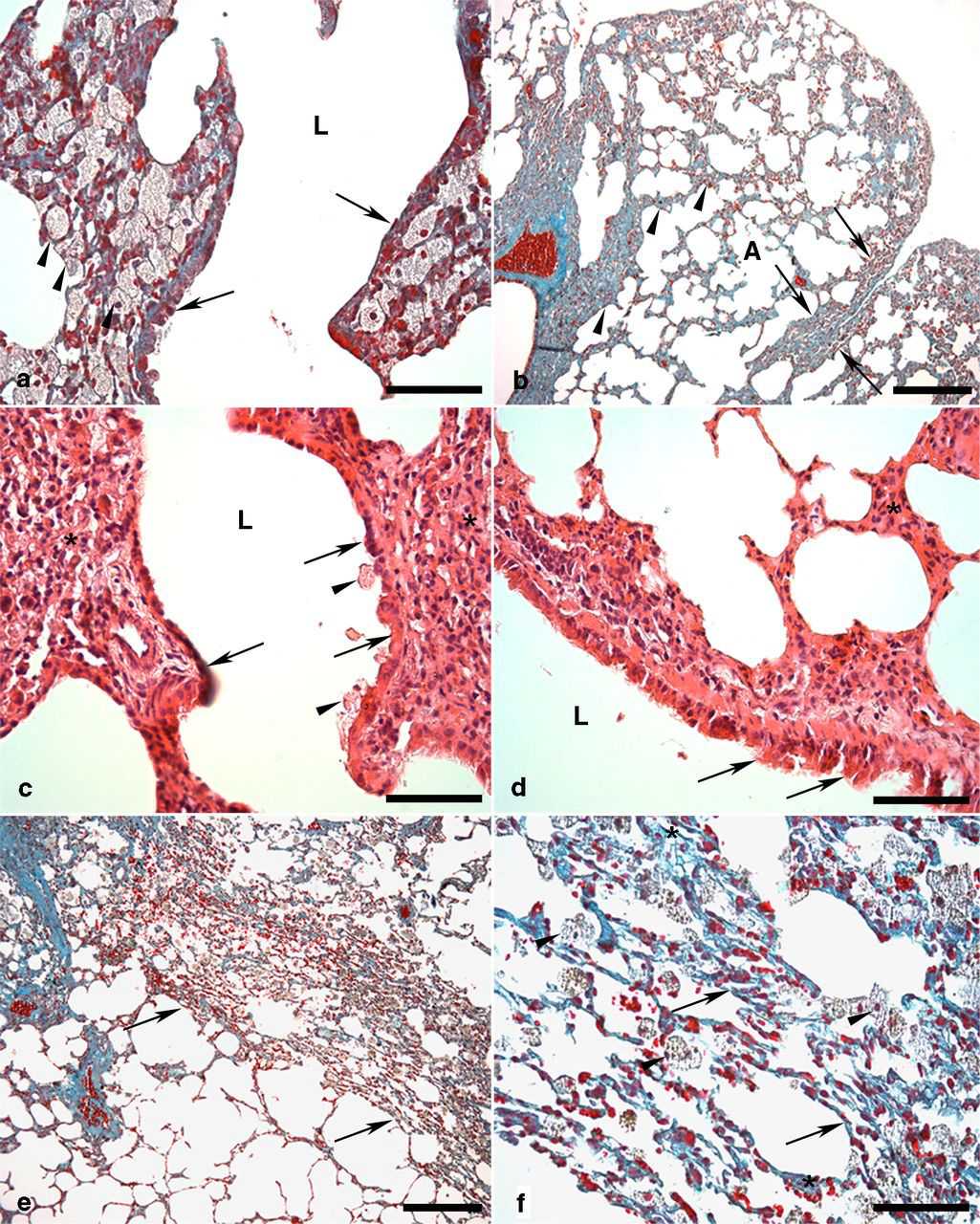
Morphometric analyses of fibrous connective tissue and type I collagen
The area of pulmonary fibrous tissue in BLM-SED (4,647.61 ± 939.69 μm2) was significantly larger than C-SED (179.83 ± 51.27 μm2), C-EXE (262.29 ± 54.81 μm2) and BLM-EXE (649.88 ± 98.35 μm2). Although training attenuated the lung fibrous tissue in the BLM-EXE group, the difference was still increased when compared with groups C-SED and C-EXE (Figure 5a).
(a) Comparing the areas of pulmonary fibrous connective tissue (μm2) of sedentary and trained mice in the control groups (C-SED and C-EXE) and in the bleomycin groups (BLM-SED and BLM-EXE). (b) Comparing the stained areas of type I collagen (μm2) of groups C-SED, C-EXE, BLM-SED and BLM-EXE. (c) Comparing the stained areas of surfactant protein A (SP-A, μm2) of C-SED, C-EXE, BLM-SED and BLM-EXE groups. (d) Comparing the number of pneumocytes II (PII) of C-SED, C-EXE, BLM-SED groups and BLM-EXE. *P < 0.05 in relation to other groups, $
P < 0.05 in relation to C-SED and C-EXE groups, #
P < 0.05 in relation to C-SED group. C-SED, control sedentary; C-EXE, control exercised; BLM-SED, bleomycin-treated sedentary; BLM-EXE, bleomycin-treated exercised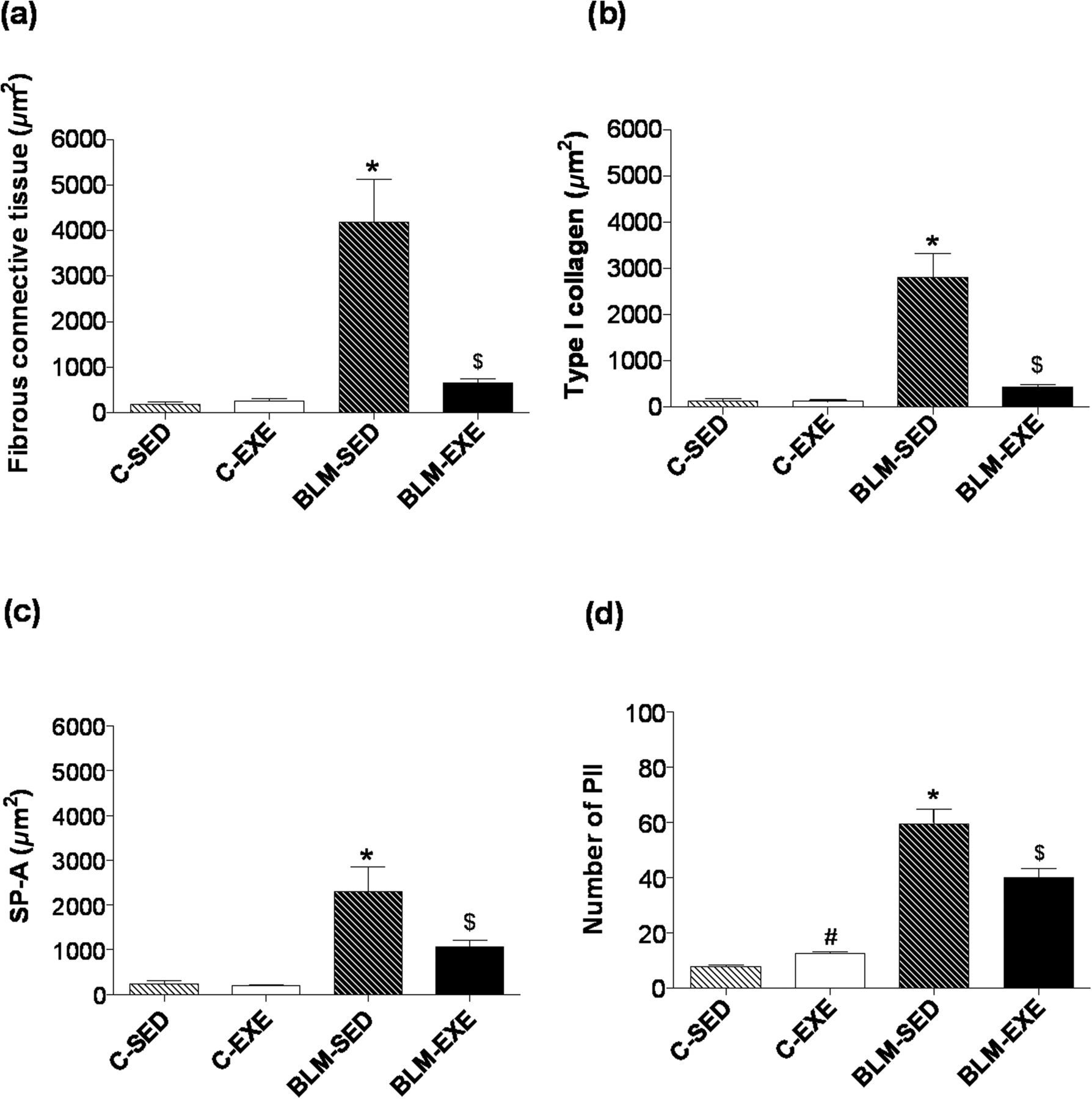
In histological sections where the anticollagen type I antibody was replaced by PBS, no positive reaction was observed in the pulmonary interstitium or in other areas (Figure 6a). In C-SED and C-EXE, levels of immunolabeling were within the normal range (Figure 6b). Differently, in BLM-SED, the interstitium presented thick areas of positive labeling for collagen I in the alveolar septa, peribronchiolar and perivascular stroma and in interlobular and subpleural spaces (Figures 6c and d). Animals in the BLM-EXE group presented a visible decrease in collagen I-positive areas in all interstitial spaces, when compared with BLM-SED (Figures 6e and f).
Immunohistochemical reaction for type I collagen in lung parenchyma of mice treated with or without bleomycin-sedentary or submitted to swimming. (a) Negative control for type I collagen. Note the absence of a brown coloration. (b) C-SED group: brown coloration in alveolar interstitium showing positive reaction for type I collagen (arrowheads). (c) BLM-SED group: positive areas for type I collagen in a region of great interalveolar (arrowheads) and subpleural (arrows) septal thickening. (d) BLM-SED group: thickening of interalveolar septa mainly due to deposition of type I collagen (arrowheads). (e) BLM-EXE group: Type I+ collagen areas in interalveolar septa with moderate thickening (arrowheads). (f) BLM-EXE group: interalveolar septa thickening associated with type I+ collagen areas (arrowheads). Counterstaining with Harris Hematoxylin. Bar = 30 μm. C-SED, control sedentary; BLM-SED, bleomycin-treated sedentary; BLM-EXE, bleomycin-treated exercised. (A color version of this figure is available in the online journal)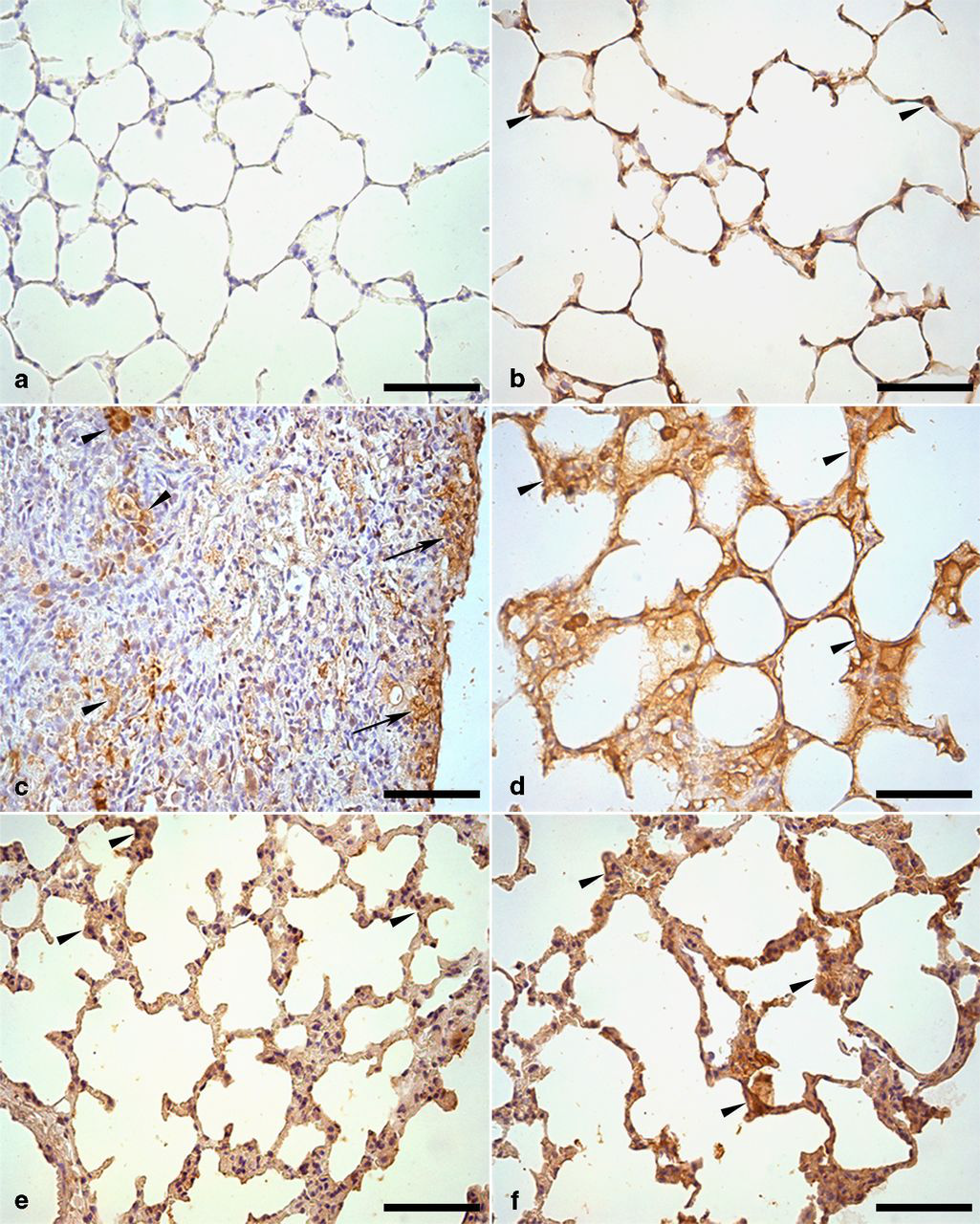
The collagen I-positive areas in BLM-SED (2,814.24 ± 505.73 μm2) were significantly higher than those of groups C-SED (143.65 ± 40.75 μm2), C-EXE (130.65 ± 20.79 μm2) and BLM-EXE (427.20 ± 66.12 μm2). Physical training attenuated the area immunolabeled for collagen type I (BLM-EXE), but remained enlarged as compared with C-SED and C-EXE. The results were expressed as average of the area in 30 analyzed images ± standard error (Figure 5b).
Area of SP-A and PII counting
In histological sections, the negative control (omission of primary antibody) did not show SP-A+ staining (Figure 7a). In C-SED and C-EXE, SP-A+ areas were observed on the alveolar walls and in the PII cytoplasm, as well as in the clear cells and on the epithelial surfaces of bronchioles (Figure 7b). In BLM-SED, very evident SP-A+ areas were visualized in lung interstitial spaces, especially interalveolar. A large amount of PII SP-A+ was often observed as cell clusters in all of these interstitial areas (Figures 7c and d). When compared with BLM-SED, the animals in BLM-EXE showed decreased amounts of SP-A+ areas in the same regions of lung parenchyma and, less often, the presence of PII SP-A+ clusters (Figures 7e and f).
Immunohistochemical reaction for surfactant protein A (SP-A) in the lung parenchyma of mice treated with or without bleomycin-sedentary or submitted to swimming. (a) Negative control for SP-A. Note the absence of the brown coloration. (b) C-SED group: brown coloration on the alveolar walls (arrowheads), showing positive reaction for SP-A. (c) BLM-SED group: SP-A+ areas in the thickened interalveolar interstitium (arrowheads) and in a large amount of type II pneumocytes (PII) (arrows). (d) BLM-SED group: positive stain for SP-A in bronchial epithelium (arrows), in the thickened interalveolar interstitium and PII (arrowheads). (e) BLM-EXE group: PII positive for SP-A (arrowheads) in interalveolar septa with thickness similar to normal. (f) Similar aspects to those of the previous figure, showing a positive reaction for SP-A in bronchial epithelium (arrow) and in PII (arrowheads). Counterstaining with Harris Hematoxylin. Bar = 30 μm. C-SED, control sedentary; BLM-SED, bleomycin-treated sedentary; BLM-EXE, bleomycin-treated exercised. (A color version of this figure is available in the online journal)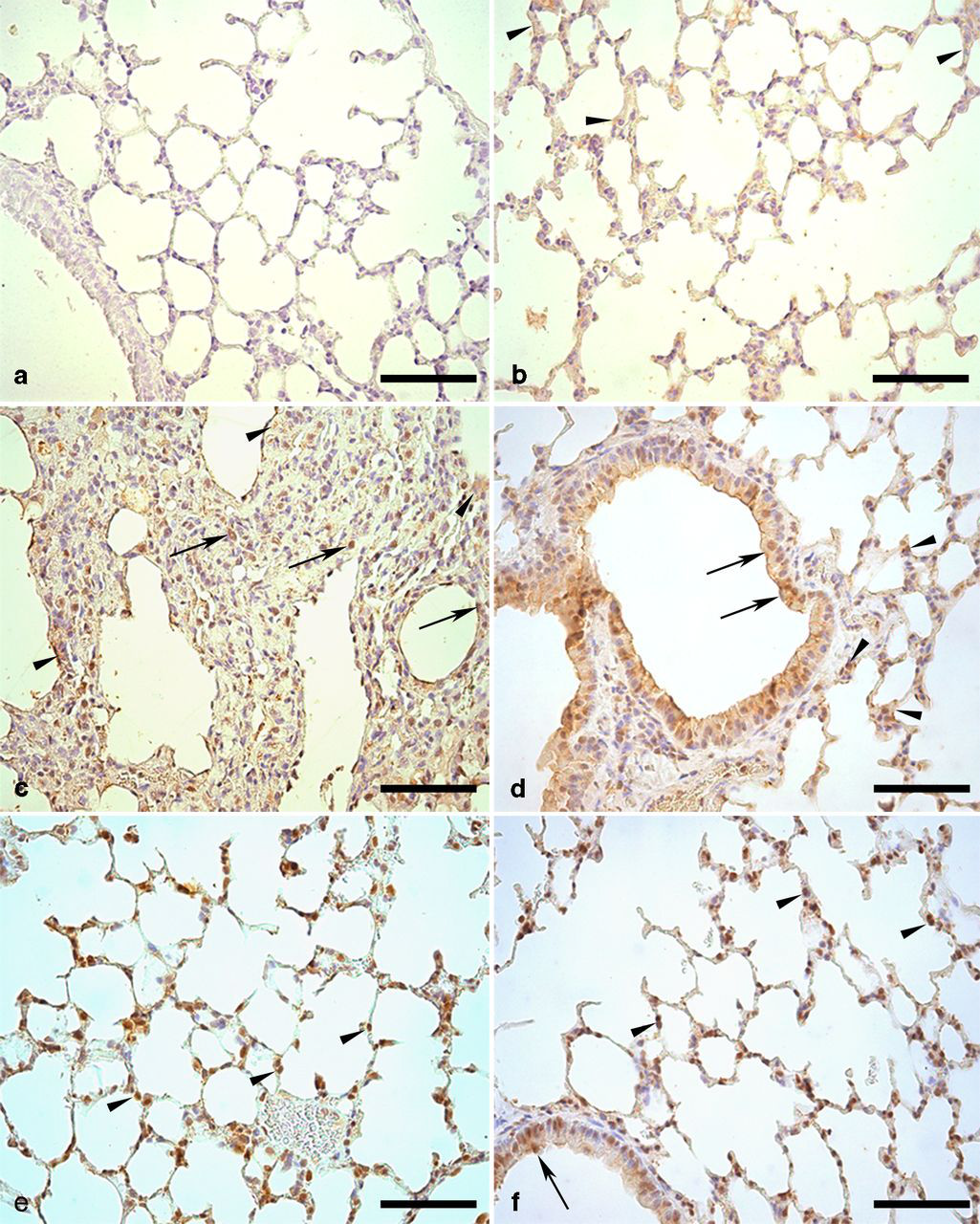
The SP-A+ area was significantly enlarged in BLM-SED (2,304.21 ± 546.51 μm2; P < 0.05) when compared with the other groups. Physical exercise attenuated such response, as observed in BLM-EXE (1,066.32 ± 155.82 μm2), but it remained elevated in relation to C-SED (245.01 ± 61.35 μm2) and C-EXE (197.35 ± 19.82 μm2) (Figure 5c). In C-EXE (12.39 ± 0.59; P < 0.05), the number of PII was higher when compared with C-SED (7.76 ± 0.50). In BLM-SED (59.41 ± 5.26; P < 0.05), the number of PII was significantly higher than control groups. BLM-EXE (40.02 ± 3.17) showed a significant decrease in the number of PII and SP-A+ area when compared with BLM-SED (P < 0.05). However, BLM-EXE showed a higher number of PII than C-SED and C-EXE (P < 0.05) (Figure 5d).
Correlation analysis
The exercise endurance capacity evaluation did not display any correlations among the evaluated parameters (connective tissue, type I collagen, PII and SP-A). As expected, the fibrous connective tissue presented a strong positive correlation with type I collagen (r = 0.9709 and P < 0.0001, Figure 8a). Similarly, the connective tissue presented a strong positive correlation with the number of PII (r = 0.7225 and P < 0.0001, Figure 8b), and the SP-A+ area (r = 0.7738 and P < 0.0001, Figure 8c). As in the case of the fibrous connective tissue, a strong positive correlation was observed between the type I collagen area and the number of PII (r = 0.7069 and P < 0.0001, Figure 8d), and for the SP-A+ area (r = 0.7462 and P < 0.0001, Figure 8e). A strong and positive correlation was observed between the SP-A+ area and the number of PII (r = 0.8167; P < 0.0001) (Figure 8f).
Correlation among the different variables evaluated, considering C-SED, C-EXE, BLM-SED and BLM-EXE groups. (a) Correlation between the area of fibrous connective tissue (μm2) and the area stained for type I collagen (μm2). (b) Correlation between the fibrous connective tissue area (μm2) and the number of type II pneumocytes (PII). (c) Correlation between the area of fibrous connective tissue (μm2) and the SP-A+ areas. (d) Correlation between the areas of type I collagen (μm2) and the number of PII; (e) Correlation between the area of type I collagen (μm2) and the SP-A+ areas. (f) Correlation between the SP-A+ areas and the number of PII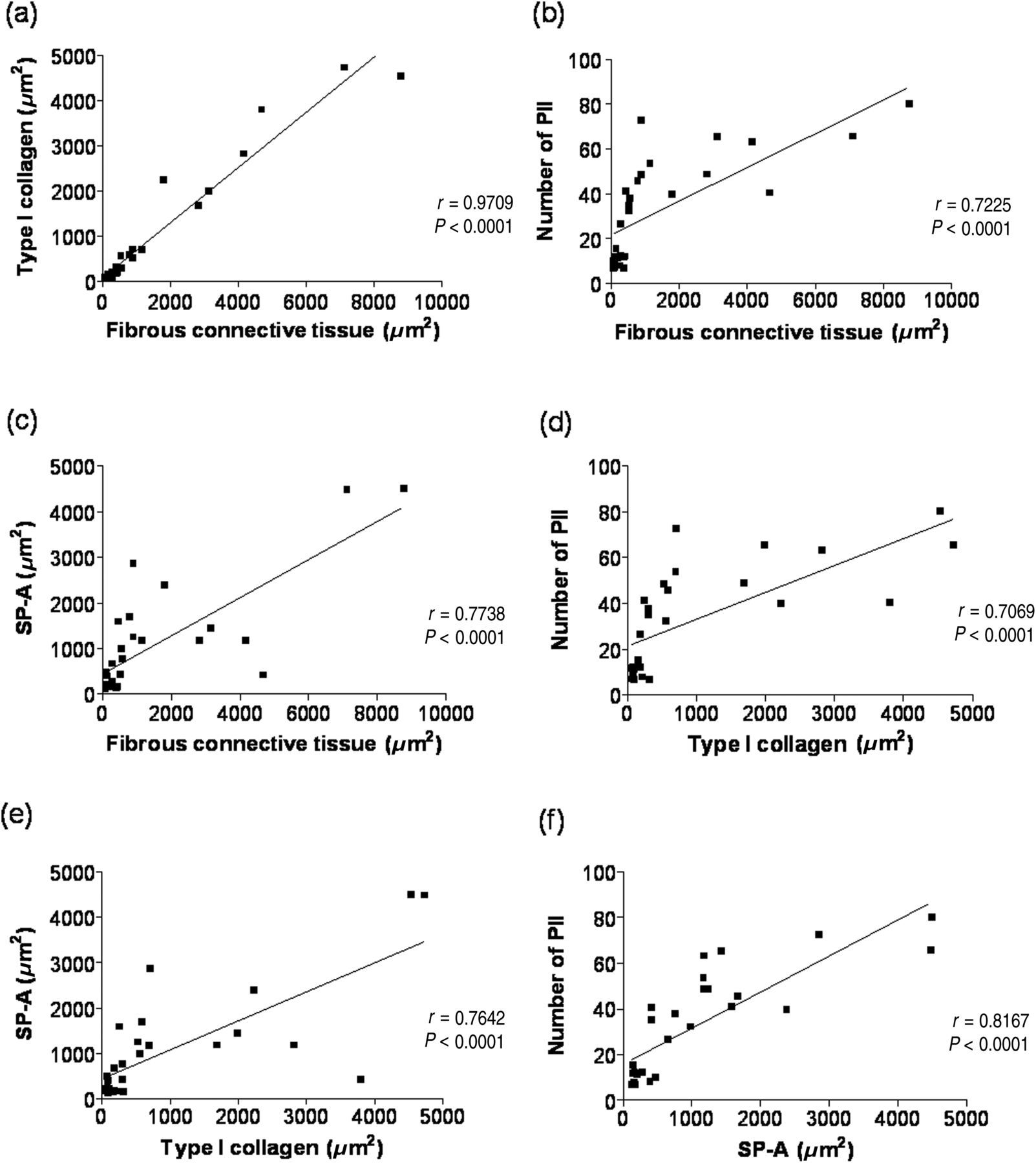
Discussion
The present study showed that aerobic exercise significantly attenuated the fibrous connective tissue, type I collagen, hyperplasia of PII and the release of alveolar SP-A in animals with BLM-induced pulmonary fibrosis. Additionally, it was observed that the exercise training program prevented functional decline in mice treated with BLM when compared with sedentary groups, suggesting that exercise training programs might play an important role as a non-pharmacological therapeutic strategy in the treatment of interstitial lung diseases like IPF. Previous observations described in humans have shown that exercise for respiratory muscles and physical training on an ergometric bicycle significantly reduced the dyspnea and improved life quality in IPF patients. 14
There are different routes of BLM administration. The BLM delivered intratracheally may induce fibrosis that progresses or persists for 60–90 days or can begin to resolve after this period. In this study, the animals were euthanized 74 days after BLM instillation and the presence of pulmonary fibrosis was confirmed macroscopically and by histopathology microscopically. As the exercise was applied 23 days after BLM administration, it was considered an antifibrotic treatment instead of an anti-inflammatory treatment that is administered during the early phase of this disease. 15 The BLM animal model is the most commonly used model of pulmonary fibrosis. However, this model has limitations with regard to understanding the progressive nature of human IPF. 4 BLM causes an inflammatory response and the subsequent development of fibrosis. However, unlike what is found in patients, the slow, irreversible progression of IPF is not reproduced in the BLM model. Studies with animal models are essential to understanding pathophysiology and the development of novel therapeutic strategies. Thus, efforts to develop even better disease models should be prioritized. 16
In order to evaluate the effect of exercise on the exercise endurance capacity in animals with pulmonary fibrosis, the animals were submitted to an adaptation program and later to an exercise training program in the swimming pool. Comparing the first and second evaluations, it was possible to observe that the control group had its exercise endurance capacity increased significantly after the exercise training program. When comparing the groups at the end of the exercise training program, considering only the last exercise endurance capacity evaluation, a significant difference between the groups BLM-EXE and BLM-SED was verified, which was due to preservation of the endurance in group BLM-EXE, beyond a significant fall in the endurance in group BLM-SED, demonstrating that the aerobic exercise in the swimming pool preserved the exercise endurance capacity in BLM-treated animals.
The model used in our study did not produce significant difference in the endurance capacity between the groups C-SED and BLM-SED. A possible explanation for these findings would be that reduced exercise tolerance in IPF is multifactorial. Gas exchange and ventilatory impairments, increased dead space ventilation, respiratory and peripheral muscle dysfunction and increased elastic inspiratory load might be involved in this response. Moreover, cardiovascular and nutritional abnormalities may also be involved. 17 Moreover, the test used in the present study may not have provided a regular pattern of muscle response to the load and oxygen uptake (VO2) that could allow the detection of respiratory dysfunction in the BLM-SED group. We believe that the severity of the damage observed in the study led to significant pulmonary dysfunction. This assertion is based on in previous studies having used BLM as a pulmonary fibrosis model. 18–21 However, in studies of this nature, this is one of the models suggested for the evaluation of physical capacity. 10,11 Despite these limitations, the present study demonstrated that physical activity reversed the damage caused by BLM in the BLM-EXE group, which may explain the preservation of endurance capacity in this group. This is a new and important finding in an experimental model aimed at evaluating both the evolution of lung damage and the performance of animals.
Lung macroscopic changes characteristic of fibrosis were observed in sedentary animals treated with BLM, similar to those observed in rats receiving BLM. 22 Such changes were observed discreetly in only one of the trained animals, treated with BLM. Microscopically, fibrosing interstitial pneumonitis similar to that observed in humans was observed in the lung parenchyma of animals which received BLM. 23,24 In BLM-EXE group, a visible reduction of fibrosis in all areas and the restoration of a large part of the alveolar air space were observed, when compared with the BLM-SED group.
Similar results were observed in the quantitative analysis of type I collagen, the main constituent of the pulmonary connective tissue. 25 In animals in the BLM-SED group, thick collagen I-positive areas were observed, especially in the alveolar septa and peribronchiolar and perivascular interstices, associated or not to clusters of alveolar epithelial and interstitial cells. Its strong correlation with the area of fibrous connective tissue shows that the increase in fibrosis is due mainly to the deposition of type I collagen. Type I collagen is produced in large amounts during fibrotic reactions and its deposition causes a loss in the normal alveolar architecture and contributes to a decrease of pulmonary complacency, a reduction of gas exchanges and other functional changes in the lungs. 26 Based on this reasoning, it is likely that the decline in exercise endurance capacity is associated with the high degree of fibrosis observed in BLM-SED animals.
The effects and mechanisms of aerobic exercise on different pulmonary pathological conditions, as well as on the immune system, are not yet fully understood. It is suggested that low and moderate intensity aerobic exercises can exert a protective effect against pulmonary allergic inflammation and airways modeling in experimentally-induced asthma in mice. 27 It has been demonstrated that physical activity induces an increase in the systemic levels of cytokines with anti-inflammatory properties from the skeletal muscles. 28
The lung epithelium, particularly the PII, performs a critical role in initiating and perpetuating pulmonary fibrosis in the presence of different endogenous and exogenous attacks. 29 In lung fibrosis, the regenerative process response is altered and the alveolar type II cells, progenitor cells of the pulmonary epithelium, usually proliferate after epithelial cell injury. In the current study, the number of PII dramatically increased after 74 days of BLM-induced lung fibrosis (BLM-SED group). We observed that the exercise modulated the number of PII differently. In healthy animals (C-EXE), the exercise slightly increased the PII number whereas in animals with BLM-induced lung fibrosis, the PII number were strongly attenuated. Such results suggest a dual role of exercise on PII, with a slight increase in the physiological conditions and an important reduction in hyperplasia in pathological conditions. In addition, the reduction of PII presented a strong positive correlation with the decrease of type I collagen and the area of fibrous connective tissue, suggesting that PII reduction may be favorable in order to re-establish the physiological lung balance in a mice model of fibrosis. In other words, an effective experimental therapeutic procedure is expected to lead to a decrease in PII after an initial phase of hyperplasia due to the capacity of differentiation to PI. Indeed, the increase of PII could exacerbate the intensity of lesions due to a probable epithelial–mesenchymal transition. Some studies have demonstrated the up-regulation of alpha-smooth muscle actin and vimentin in E-cadherin and surfactant protein C-expressing cells. 30,31 Considering this phenotypic change, the accumulation of PII in a given phase of lung damage could be directly involved in the excessive deposition of extracellular matrix.
The expression of the pulmonary surfactant's SP-A is considered a specific marker of pulmonary disease. In IPF, its concentration appears decreased in BAL and increased in the plasma as compared with healthy individuals. The present study shows, for the first time, that in pulmonary fibrosis, there was an expressive increase of SP-A in the lung parenchyma and that exercise attenuated such a response. Serum SP-A levels may be useful in predicting the course of IPF patients. Patients with normal serum SP-A levels had a better prognosis than those with elevated serum levels. 32
Considering the significant reduction of lesions in group BLM-EXE (fibrous connective tissue and collagen type I, hyperplasia of PII and area of SP-A) compared with group BLM-SED, it is possible that the preserved endurance capacity could be associated with the improvement in lung pathology.
In conclusion, the present study showed that physical exercise preserved the exercise endurance capacity and significantly attenuates the lesions in mice with BLM-induced pulmonary fibrosis. Such data suggest that physical exercise modulates pathophysiology and attenuates progression of pulmonary lesions, and may constitute a non-pharmacological therapeutic option for this illness.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by FAPEMIG (Fundação de Amparo à Pesquisa do Estado de Minas Gerais) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico). The authors are grateful to Professor Gregory T Kitten for the English review of this manuscript, and Mirna MD Souza and Gislene A Silva for the technical assistance.