
Abstract
A novel polypeptide (Mere15) was purified from Meretrix meretrix Linnaeus by ammonium sulfate fractionation, ion exchange, gel filtration and reversed phase chromatography. Mere15 exhibited selective cytotoxicity to several human cancer cells. In vivo study showed that Mere15 significantly suppressed the growth of human lung adenocarcinoma A549 xenograft in nude mice. The mechanism was associated with a G2/M phase arrest followed by apoptosis, including membrane blebbing, loss of mitochondrial membrane potential, externalization of phosphatidylserine, chromosome condensation and DNA fragmentation. Western blot analysis showed that the intrinsic pathway was involved in Mere15-induced apoptosis. These results suggest that Mere15 may have therapeutic potential for the treatment of non-small-cell lung carcinoma.
Introduction
Lung cancer continues to be the leading cause of cancer-related death in both men and women worldwide. 1 About 80% of lung cancer is made up of non-small-cell lung carcinoma (NSCLC), which includes adenocarcinoma, large-cell carcinoma, squamous-cell carcinoma and some rare subtypes. Unfortunately, more than 50% of patients diagnosed with NSCLC already possess locally advanced or metastatic disease and the median survival is only 4–5 months. 2 In the past decade, a number of new agents (gemcitabine, vinorelbine and taxanes) and the third-generation chemotherapy program constituted by platinum has been used to prolong survival and to improve quality of life of NSCLC patients. 3 However, the effects of these drugs have reached a therapeutic plateau and the overall five-year survival rate remains dismal at 15%. 1 Therefore, novel and more effective drugs are urgently required to improve the efficacy in the treatment of NSCLC and to prolong survival.
The diversity of the marine environment is immeasurable, and is a rich resource for discovering novel biologically active compounds, especially for anticancer drugs. 4 A series of new marine natural products such as ecteinascidin-743, aplidine and pachymatismin have, over recent years, shown high antitumor activity against NSCLC. 5–7 Among them, there are several peptides. For example, kahalalide F (KF), a complex depsipeptide isolated from the herbivorous marine mollusk Elysia rufescens, possesses a strong cytotoxic activity in NSCLC. 8
The marine mollusk Meretrix meretrix Linnaeus is a traditional Chinese medicine used for cancer and other diseases. Previous reports showed that several peptides and proteins derived from M. meretrix exhibited anticancer activities. Leng et al. 9 demonstrated that a peptide extracted from M. meretrix strongly inhibited the growth of human gastric gland carcinoma cells by affecting the activity of superoxide dismutase, alkaline phosphatase and tyrosinase. Previous studies from our laboratory have shown that a 40 kDa protein MML exhibited significant cytotoxicity to several cancer cell lines by changing the cell membrane permeability and inhibiting the tubulin polymerization. 10 However, the antitumor mechanisms of the peptides and proteins from M. meretrix have rarely been investigated in NSCLC cells. In the current study, another novel peptide (Mere15) with antitumor activity was extracted and purified from M. meretrix Linnaeus. Furthermore, the potential antitumor mechanism of the novel polypeptide against NSCLC was investigated.
Materials and methods
Materials
M. meretrix Linnaeus was purchased from the Fishery Company of Weifang (Weifang, China). CM-Sepharose Fast Flow, Superdex 75 and Resource 15Q pre-packed columns were obtained from GE Healthcare (Piscataway, NJ, USA). A C18 column was purchased from Agilent (Palo Alto, CA, USA). The bicinchoninic acid (BCA) protein assay kit was purchased from Pierce (Rockford, IL, USA). The Universal Genomic DNA extraction kit was from Takara (Dalian, China). The Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit was from Cusabio (Carlsbad, CA, USA). The JC-1 test kit was from Beyond (Nantong, China). The primary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The secondary antibodies were purchased from Bio-Rad (Hercules, CA, USA). The SuperSignal Substrate was from Pierce. The Coulter DNA Prep Reagents Kit was purchased from Beckman Coulter (Fullerton, CA, USA). Cyclophosphamide (CTX; 99%) was purchased from Hengrui Pharmaceuticals Co Ltd (Jiangsu, China).
Extraction and purification
The fresh M. meretrix Linnaeus was split and the fluids from the cavity were collected. Then, samples were centrifuged at 10,000 ×
The protein concentration was determined by measuring absorbance at 562 nm with a spectrophotometer DU-650 (Beckman, Fullerton, CA, USA) using the BCA protein assay kit. The purity was determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) as previously described. 11 The purified peptide (Mere15, 2 μg) was used to analyze with 15% polyacrylanmide gel. Its molecular mass was determined by comparison with the marker proteins.
Cell lines and cell culture
The human colorectal carcinoma RKO, human cervical carcinoma HeLa and human breast epithelial MCF-10A, as well as the murine fibroblast NIH 3T3 cell lines, were obtained from the American Type Culture Collection (Rockville, MD, USA). Human hepatocellular carcinoma BEL-7402, human lung adenocarcinoma A549, human breast carcinoma MCF-15 and human pancreatic cancer Aspc-1 cells were provided by the Cell Bank of Chinese Academy of Sciences (Shanghai, China). Human lung cancer A549 cells were cultured in F-12K medium supplemented with 10% heat-inactivated fetal bovine serum, 2 mmol/L glutamine, 100 U/mL of penicillin and 100 μg/mL streptomycin. HeLa, MCF-10A, Aspc-1, NIH 3T3 and MCF-15 cells were cultured in Dulbecco's Modified Eagle's Medium with complete supplement as above. The others were cultured in RPMI-1640 medium. Cells were grown in a 37°C incubator supplied with 95% air and 5% CO2.
Cell viability assessment
The effect of Mere15 on viability of tumor cell lines as well as benign cell lines NIH 3T3 and MCF-10A was assessed in vitro by MTT assay. 12 In brief, A549 cells (4 × 103) were plated onto 96-well plates and cultured in 180 μL of F-12K medium for 24 h. Then, Mere15 with certain concentrations (0, 15, 30, 45, 60 and 75 μg/mL) was added to the medium. After incubation for 48 h, MTT dye (100 μL, 0.5 mg/mL) was added to the culture medium and the cells were cultured for another four hours at 37°C. The medium was removed and 150 μL dimethyl sulfoxide was added to dissolve the formazan crystals. The OD570 was measured by an ELx800 microplate reader (Bio-Tek, Winooski, VT, USA) with subtraction of background absorbance. The cytotoxicity of Mere15 was expressed as an IC50, defined as the concentration causing a 50% reduction of cell growth compared with untreated cells.
Cell morphological changes
Morphological changes of A549 cells were observed by a scanning electron microscope (SEM). Briefly, A549 cells were grown on poly-
Nuclear staining with Hoechst 33342
To evaluate chromosome condensation, Hoechst 33342 staining was performed as described previously. 13 Briefly, A549 cells were seeded in six-well plates at a density of 1 × 105 cells/well and cultured for 24 h. Mere15 (0, 15, 30 and 45 μg/mL) was added and cultured for another two hours. The cells were washed with PBS twice and stained with Hoechst 33342 (5 μg/mL) in the dark for 20 min at room temperature. Then, cells were rinsed in PBS twice and visualized with a fluorescence microscopy (Zeiss, Oberkochen, Germany). Apoptotic cells were defined by the condensation and fragmentation of their nuclei.
DNA ladder assay
A549 cells were seeded in six-well plates at a density of 1 × 105 cells/well and incubated for 24 h. Then, certain concentrations (0, 15, 30 and 45 μg/mL) of Mere15 were added and cultured for another 24 h. The cells were collected and total genomic DNA was extracted using the Universal Genomic DNA extraction kit following the manufacturer's protocol. DNA fragmentation was analyzed on 1.5% agarose gel and stained with ethidium bromide.
Annexin V-FITC/propidium iodine assay of apoptotic cells
The externalization of phosphatidylserine during the progression of apoptosis was detected using the Annexin V-FITC Apoptosis Detection Kit according to the manufacturer's protocol. Briefly, A549 cells (1 × 105) were plated in six-well plates for 24 h. Mere15 (0, 15, 30 and 45 μg/mL) was added and incubated for additional 24 h. Then, cells were harvested, washed with ice-cold PBS twice and resuspended in 400 μL binding buffer. Then, Annexin V-FITC (5 μL) and propidium iodide (PI) (5 μL) were added. Cells were gently vortexed, incubated in the dark for 10 min at room temperature and immediately analyzed using an EPICS XL-MCL flow cytometry system (Beckman Coulter, Miami, FL, USA).
Mitochondrial membrane potential assay
The reduction of mitochondrial membrane potential during apoptosis was measured by employing the JC-1 (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide) test kit following the manufacturer's instructions. Briefly, cells were seeded in six-well plates with 1 × 105 per well and allowed to grow for 24 h. Mere15 with certain concentrations (0, 15, 30 and 45 μg/mL) was added and incubated for one hour. Cells were collected by centrifugation at 3000 ×
Western blot analysis
The A549 cells were seeded in six-well plates, untreated or treated with Mere15 (15, 30 and 45 μg/mL). Then, the cells were collected by centrifugation at 1000 ×
Cell cycle analysis
A549 cells were seeded in six-well plates for 24 h as described above. Supernatants from the cultured plates were aspirated out and replaced with fresh medium containing Mere15 at certain concentrations (0, 15, 30 and 45 μg/mL). After incubation for 24 h, cells were collected by centrifugation at 1000 ×
In vivo antitumor activity
Antitumor activity in vivo was carried out using A549 xenograft model. The 6- to 8-week-old athymic nude mice (BALB/c, nu/nu) were obtained from the Institute of Experimental Animals, Chinese Academy of Medical Sciences & Peking Union Medical College (Beijing, China). Exponentially growing cells were harvested, adjusted to 4 × 106 per 200 μL and injected subcutaneously into the right axilla of each nude mouse. After postimplantation seven days, the nude mice were randomly divided into five groups (six mice per group) as follows:
Group A received normal saline; Group B received CTX 50 mg/kg as positive drug; Groups C, D and E received Mere15 with 12.5, 25.0 and 50.0 mg/kg, respectively.
All mice were maintained in a pathogen-free isolator on hardwood bedding with a 12-h dark/light cycle, and with ad libitum access to the same standard chow. Each group was treated by intraperitoneal injection every day for 10 days.
After being grown for another four days, all mice were euthanized. Then, the final body and tumor weight were recorded and the mean weights were calculated. The tumor growth inhibition (TGI) for each group was calculated using the following formula: TGI = (1−mean treated tumor weight/mean control tumor weight) × 100%. The tumor tissues were snap-frozen in liquid nitrogen and stored at −80°C for Western blot analysis. The animal experiments were approved by the Experimental Animal Center of Shandong province, China, and the mice were treated in accordance with international animal ethics guidelines.
Statistical analysis
Statistical analyses were performed using the software SPSS for Windows 13.0 (SPSS Inc, Chicago, IL, USA). Values were expressed as mean ± SD. To analyze the data statistically, we performed Student's t-test for the analysis. Statistical significance was set at P < 0.05.
Results
Extraction and purification of the antitumor polypeptide from M. meretrix Linnaeus
In our initial step, we used ammonium sulfate to isolate the components. The crude extracts obtained from the portion of 30–70% ammonium sulfate saturation were ultrafiltered and applied to a CM-Separose Fast Flow column. As shown in Figure 1a, there are three protein peaks. The MTT assay showed that peak C2 eluted with 0.1 mol/L sodium chloride solution displayed cytotoxicity on A549 cells. Therefore, the active fraction was pooled and further purified using a Superdex 75 column. Three peaks were obtained, of which the last one (S3) showed strong activity (Figure 1b). The active fraction (S3) was purified on a Resource 15Q pre-packed column and the second fraction (R2) (Figure 1c) was found to exhibit cytotoxicity. This fraction was further purified to homogeneity by reversed-phase high-performance liquid chromatography using a C18 column (Figure 1d). This resulted in a single band with a molecular mass of 15 kDa as analyzed by SDS-PAGE analysis (Figure 1e), and this peptide was then designated Mere15.
Purification and sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) pattern of Mere15. (a) The 5–50 kDa portion was applied to a CM-Sepharose Fast Flow column. The column was washed with sodium acetate buffer (50 mmol/L, pH 5.5) and eluted with the same buffer containing 0.1 and 0.5mol/L sodium chloride at 10 mL/min. (b) The active fraction C2 was applied to a Superdex 75 column and eluted with sodium acetate buffer (50 mmol/L, pH 5.5) containing 0.15 mol/L sodium chloride at 0.8 mL/min. (c) The active fraction S3 was loaded for fast protein liquid chromatography with a Resource 15Q pre-packed column. The column was washed with Tris-HCl buffer (20 mmol/L, pH 9.0) and then eluted with a linear gradient of 0.5 mol/L sodium chloride in the same buffer from 0% to 50% at 1 mL/min. (d) The active fraction R2 was subjected to reversed-phase high-performance liquid chromatography on a C18 column pre-equilibrated with 0.1% (v/v) trifluoroacetic acid in water. The column was eluted with a linear gradient of acetonitrile in the same buffer from 0% to 100% at 0.5 mL/min. (e) Mere15 was analyzed by SDS-PAGE and stained with Coomassie brilliant blue R250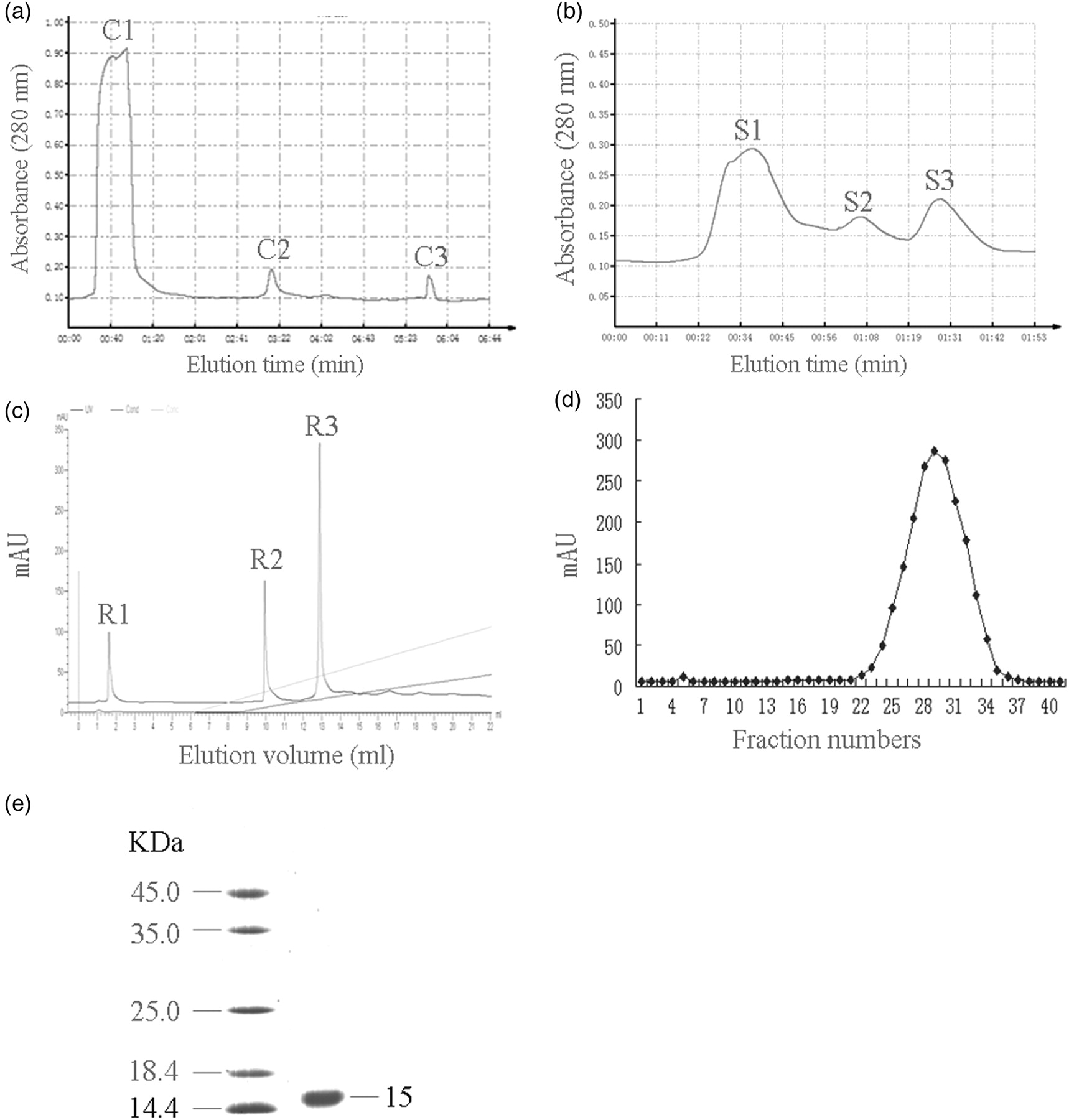
Mere15 inhibited the viability of human tumor cell lines
In the next step, we studied the cytotoxicity of Mere15 towards several human cancer cells. Our results showed that Mere15 exhibited cytotoxicity to a wide variety of human cancer cell lines in a dose-dependent manner (Figure 2a). The IC50 values of Mere15 on the cancer cell lines are shown in Table 1. However, Mere15 did not significantly decrease the viability of benign cells, such as NIH 3T3 and MCF-10A (Figure 2b). These results demonstrated that Mere15 exhibited selective cytotoxicity to human cancer cell lines. Among these cells lines, A549 cells were much more sensitive than other cells. Therefore, A549 cells were chosen in our subsequent experiments.
Effects of Mere15 on cell viability. The tumor cell lines (HeLa, Aspc-1, A549, RKO, MCF-15 and BEL-7402) (a) as well as benign cell lines (NIH 3T3, MCF-10A) (b) were treated with indicated concentrations of Mere15 for 48 h, and then cell viability was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Data are presented as the mean ± SD of three independent experiments IC50 for Mere15
#Normal cell lines IC50 (%) was assessed by MTT assay and calculated by linear regression. Data are presented as the mean ± SD of three independent experiments MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
Mere15 induced the morphological changes of A549 cells
Morphological description using electron microscopy remains one of the best ways to separate apoptosis from necrosis.
15
In the current study, SEM was applied to study the surface ultrastructural morphology of A549 cells. As shown in Figure 3, treatment of A549 cells with Mere15 resulted in distinct morphological changes with apoptotic characteristics, including cell shrinkage, surface membrane blebbing and microvilli reduction. Moreover, the number of blebs (cytoplasmic extension), an indicator of cell apoptosis, was found to be increased with the increasing concentrations of Mere15. In contrast, the A549 cells without Mere15 treatment showed normal morphological features. These results suggested that Mere15 inhibited the viability of A549 cells via inducing apoptosis.
16
Morphological observation by a scanning electron microscope. A549 cells were untreated (control) or treated with Mere15 at a concentration of 15, 30 and 45 μg/mL, respectively. Untreated A549 cell membrane showed a typical morphological feature of a cancer cell such as numerous microvilli (as shown in white arrow). Mere15-treated A549 cells showed distinct morphological changes associated with apoptosis, including membrane blebbing (b) and microvilli reduction (r)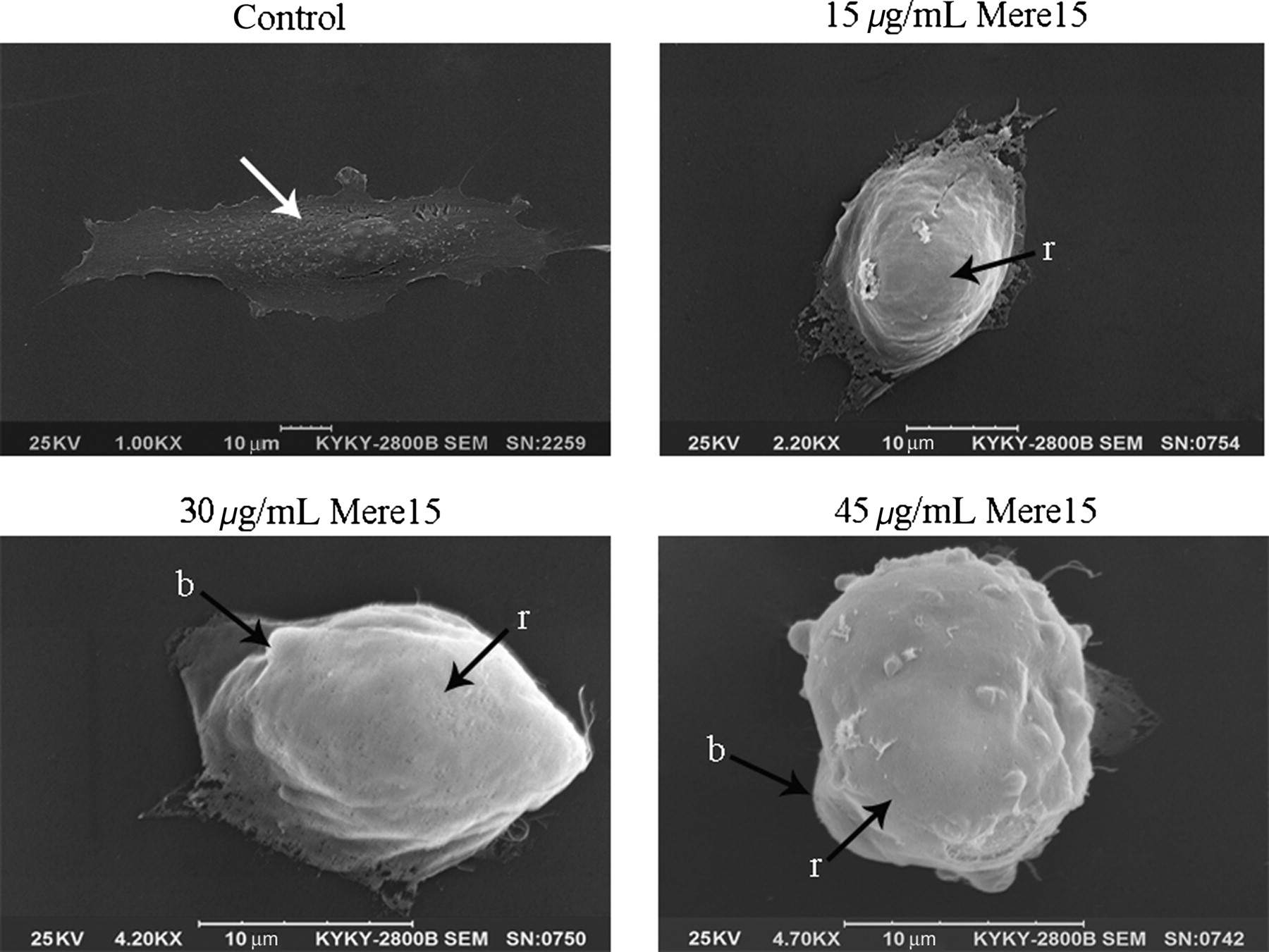
Mere15 induced apoptosis in A549 cells
In order to further study the apoptotic effects of Mere15 on A549 cells, the profile of chromosomal DNA was analyzed by Hoechst 33342 staining and DNA electrophoresis. As shown in Figure 4a, nuclear pyknosis, chromosome condensation and formation of apoptotic bodies were observed in cells treated with Mere15. Moreover, a ladder-like DNA fragmentation was observed on agarose gel with DNA extracted from the cells treated with Mere15 (Figure 4b).
Mere15 induced apoptosis in A549 cells. A549 cells were untreated (control) or treated with certain concentrations (15, 30 and 45 μg/mL) of Mere15. (a) Nuclear morphology was observed with Hoechst 33342 staining under a fluorescence microscope. The white arrows showed the condensed, fragmented nuclei. (b) Internucleosomal DNA fragmentation was analyzed by agarose gel electrophoresis with ethidium bromide staining. (c) The externalization of phosphatidylserine during the progression of apoptosis was detected. Early apoptotic cells (Annexin V+/PI−) are in the right lower quadrant. (d) The percentage of early apoptotic cells was calculated. Data are representative of three different experiments. **
P < 0.01 compared with control. (A color version of this figure is available in the online journal)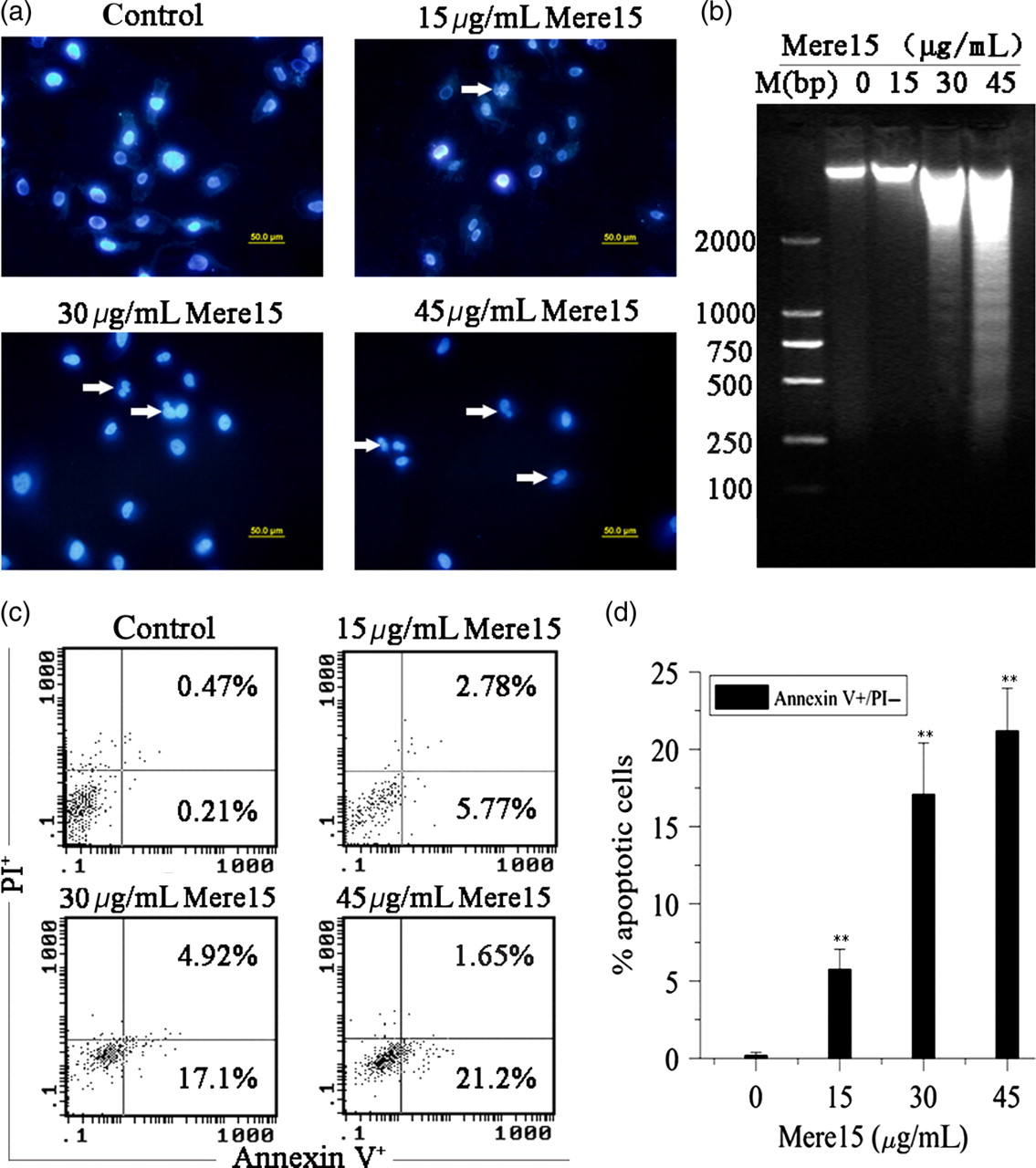
Annexin V-FITC/PI assay was performed to show the degree of apoptosis in cells treated with Mere15. As shown in Figure 4c and d, the percentage of early apoptotic cells (Annexin V-FITC-positive/PI-negative) was 0.21% in control cells. In contrast, the percentages of apoptotic cells were 5.77%, 17.1% and 21.20% when the cells were treated with Mere15 at 15, 30 and 45 μg/mL, respectively. These results further indicated that Mere15 inhibited the growth of cancer cells associated with the apoptotic pathway.
Mere15-induced apoptosis via the intrinsic pathway
Previous studies have shown that some antitumor peptides diffused into the cells to interact with the mitochondrial membrane.
17
In this study, the effect of Mere15 on mitochondrial membrane potential was evaluated using fluorescent dye JC-1 by a fluorescence plate reader. Treating A549 cells with Mere15 for one hour, the ratio of red to green fluorescence decreased significantly in a dose-dependent manner, indicating a collapse in Δψm (Figure 5a). The release of cytochrome c and the cleavage of procaspase-9, procaspase-3 and PARP (poly[ADP-ribose] polymerase) were studied by Western blot analysis. As shown in Figure 5b, Mere15 induced a dose-dependent release of mitochondrial cytochrome c into the cytosol. The cleavage of procaspase-9 and procaspase-3 were detected at a concentration of 15 μg/mL of Mere15, and gradually increased in a dose-dependent manner. Similar results were observed in PARP cleavage. These results suggested the intrinsic pathway was involved in the Mere15-induced apoptosis.
Mere15-induced apoptosis involved the intrinsic pathway. (a) Effect of Mere15 on mitochondrial membrane potential was examined using fluorescent dye JC-1. Fluorescence intensity was expressed as a ratio of 590:530 nm emission. Data are expressed as mean ± SD of three independent experiments. **P < 0.01 compared with control. (b) Mere15 induced cytochrome c release and activated caspase-9 and -3. Total cell lysates were prepared from the A549 cells treated with certain concentrations of Mere15. Western blot analysis was performed to analyze the release of cytochrome c and the cleavage of procaspase-9, procaspase-3 and poly (ADP-ribose) polymerase. (c) Mere15 alters expression of Bcl-2 to Bax and p53
To study the upstream factors resulting in the mitochondrial pathway, the Bcl-2 family proteins were assessed by Western blot analysis. As shown in Figure 5c, treatment of A549 cells with Mere15 resulted in the reduction of Bcl-2 levels in a dose-dependent manner, associated with a dose-dependent increase in the expression of Bax.
It is well documented that Bax can be activated by p53 via a transcription-dependent and transcription-independent mechanism. 18 We next studied whether p53 was involved in the Mere15-induced apoptosis. The results showed that after treatment with increasing concentrations of Mere15, the expression of p53 protein increased significantly (Figure 5c), suggesting that p53 was involved in the Mere15-induced apoptosis of A549 cells.
Mere15 induced G2/M phase arrest
As shown in Figure 6a, treatment of A549 cells with Mere15 induced a G2/M phase arrest with a decrease in the S phase population in a dose-dependent manner. The amount of cells in the G2/M phase increased from 2.8% (untreated cells) to 18.4%, 25.9% and 27.1% for A549 cells treated with Mere15 at 15, 30 and 45 μg/mL, respectively. Interestingly, the sub-G1 DNA peak, which has been considered to be the hallmark of apoptosis, was observed with the apoptotic rates of 18.1% and 24.7% after treatment with Mere15 at 30 and 45 μg/mL, respectively. These results suggested that Mere15 induced the G2/M phase arrest followed by apoptosis in A549 cells. p21, which is the cyclin-dependent kinase inhibitor under the transcriptional control of p53, plays an essential role in the G2/M phase arrest.
19
To further investigate the underlying mechanism for the G2/M phase arrest induced by Mere15, the expression of p21 protein in A549 cells was measured. Results showed that the expression of p21 protein dose-dependently increased when A549 cells were treated with Mere15 (Figure 6b).
Mere15 induced cell cycle arrest associated with the expression of p21. (a) Cell cycle analysis of A549 cells untreated or treated with certain concentrations of Mere15 was performed by flow cytometry using propidium iodide staining. The areas M1, M2, M3 and M4 in panels represent the sub-G1, G0/G1, S and G2/M phases, respectively. The results were repeated at least three times. (b) The expression of p21 protein was determined using Western blot analysis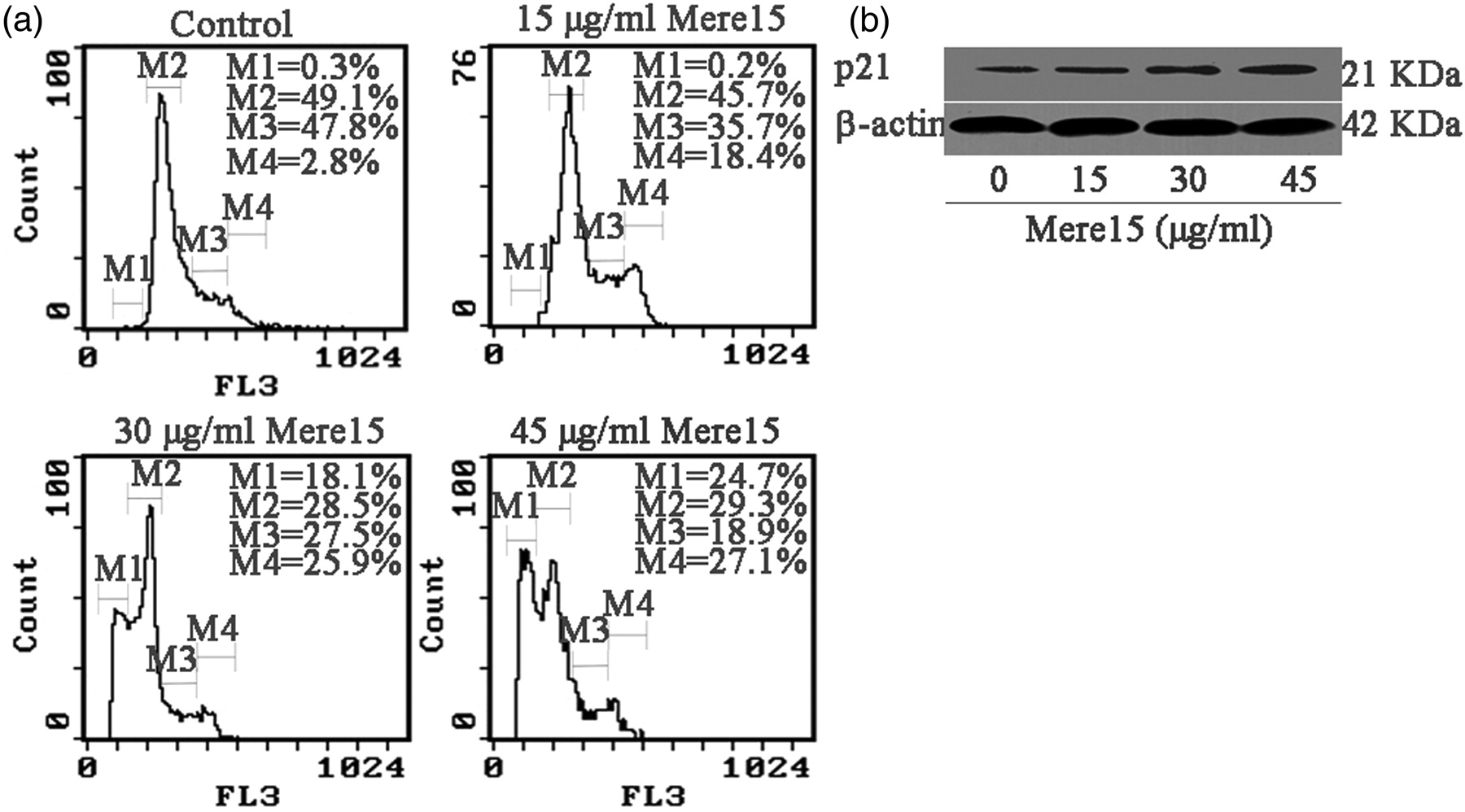
Mere15 inhibited the growth of A549 xenograft in athymic mice
Growth inhibition of A549 xenograft in athymic mice
Data are represented as the mean ± SD of three independent experiments
CTX, cyclophosphamide; TGI, tumor growth inhibition
**P < 0.01 compared with control. No mouse died during the experimental period
Discussion
M. meretrix is a popular seafood and traditionally used as a Chinese medicine for cancer. From previous studies, several bioactive peptides and proteins have been isolated from M. meretrix, and some of them have shown potent cytotoxicity to human cancer cells. 9,10 In the present study, we used a MTT assay to isolate the components with cytotoxicity, and a novel polypeptide with antitumor activity both in vitro and in vivo was obtained. This result suggests that M. meretrix contains diverse bioactive peptides, and that novel bioactive agents can be obtained using different approaches for isolation.
Our results demonstrated that Mere15 suppressed the growth of several cancer cells including human colon, breast, liver, lung, cervix and pancreas cells. Among them, Mere15 exhibited the greatest cytotoxic effect on lung cancer cells with the IC50 value of 31.8 μg/mL. However, Mere15 showed comparatively high IC50 values on normal cells (Table 1). The cytotoxicity of Mere15 can thus be characterized as having a tumor-selective manner. The exact mechanism underling the selectivity of Mere15 on cancer cells is still unknown. However, considering the apoptosis-inducing mechanism of Mere15, it is conceivable that the polypeptide may interact with specific signal molecules responsible for cancer cell apoptosis. Our laboratory has developed the monoclonal antibody of Mere15, and studies are in progress in our laboratory to identify the interacting molecules of Mere15 in cancer cells.
Studies have shown that most anticancer agents inhibit cell growth via the apoptotic pathway. 20,21 Mere15, as a novel polypeptide, also induced cell death via the intrinsic apoptotic pathway, characterized by increasing the expression of Bcl-2 and p53, resulting in the loss of mitochondrial membrane potential and subsequent release of cytochrome c, activating caspase-9 and caspase-3 (Figure 5b). These results suggested that Mere15, like most anticancer agents, induced cell death via the apoptotic pathway. However, since Mere15 belongs to a large, polar molecule, there is some difficulty for its permeating cell membrane. Further study is needed to address the mechanism.
Our present studies also showed that Mere15 significantly suppressed the growth of A549 xenograft in nude mice with an inhibition rate of 69%. Furthermore, administration of Mere15 did not alter the growth of nude mice, suggesting that there were no apparent signs of toxicity of Mere15 to nude mice (Table 2). Therefore, Mere15 might be useful as a novel, and low toxic anticancer agent for the treatment of cancer patients. However, since there is a very low content of Mere15 in M. meretrix, a novel biotechnical approach needs to be developed to obtain enough amount of the polypeptide. Initial studies have been done in our laboratory to try to dissect the amino acid sequence of Mere15. However, since some modifications existed on the N-terminal sequence of Mere15, we have not yet got the exact sequence of the polypeptide. We are now using enzymatic digestion and matrix-assisted laser desorption/ionisation-time of flight mass spectrometry approaches to sequence the amino acid of Mere15.
In conclusion, Mere15, a novel polypeptide extracted and purified from M. meretrix Linnaeus, was proven to display selective cytotoxicity in vitro against human cancer cells. Mere15 exhibited antitumor activity via the induction of the G2/M phase arrest and apoptosis, involved in the intrinsic apoptosis signaling pathway. However, the interaction of Mere15 with the cells leading to the G2/M phase arrest and apoptosis still requires further investigation. Additionally, Mere15 inhibited the growth of tumor in xenograft mice significantly. Considering its selective cytotoxicity to cancer cells and its potent antitumor activity in vivo, Mere15 may be a good candidate to be developed as a promising chemotherapeutic agent for NSCLC.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the National 863 High Technology Program of China (No. 2007AA091403 and No. 2007AA09Z408) and supported in part by the Innovative Drug Development Program in China (No. 2009ZX09103-661 and No. 2009ZX09102-240), and also by the National Natural Science Foundation of China (No. 81001396).