
Abstract
Previous studies have suggested that massive small bowel resection (mSBR) compromises the normal intestinal processes of digestion and absorption, and requires an adaptive response to regain full function and reinstate coordinated contractile activity of the circular smooth muscle. This study was designed to investigate spontaneous contractile activity of circular smooth muscle using the mSBR rat model and to determine the functional role of M2 and M3 muscarinic acetylcholine receptors (mAChR) in this process. Male Sprague–Dawley rats underwent an 80% proximal SBR or sham operation. Markers of adaptation, including villus and microvillus height, were analyzed by hematoxylin and eosin staining and transmission electron microscopy. Contractility was measured by attaching the distal ileum strips to strain gauge transducers and exposing the tissue to varying doses of the cholinergic agonist carbachol. Protein expressions of M2- and M3-mAChR in intestinal smooth muscle (ISM) were detected by Western blot. Following mSBR, the ISM showed perturbed spontaneous rhythmic contraction, irregular amplitude and slow frequency by muscle strip test. However, by two weeks after mSBR, the contractile function of circular smooth muscle was found to have returned to normal levels. Protein expression of M2-mAChR was down-regulated following mSBR but up-regulated during the adaptive process when contractile activity of circular smooth muscle was regained. These results indicate that smooth muscle contractility was spontaneously restored in rats following mSBR, and involved the acetylcholine receptors M2 and M3. Thus, the disrupted contractile response of smooth muscle in short bowel syndrome may be corrected by therapeutic intervention to restore the expressions of M2- and M3-mAChR to pre-mSBR levels.
Keywords
Introduction
Short bowel syndrome (SBS), also known as short gut syndrome, is a malabsorption disorder caused by the surgical removal of the small intestine to treat common gut-related afflictions, including Crohn's disease, tumors or obesity. 1,2 In rare cases, SBS can be due to complete dysfunction of a large segment of bowel as a result of severely restricted blood supply caused by hernia, embolus, thrombosis or low blood pressure. When over two-thirds of the intestine is absent or non-functional, the remaining surface area of small intestinal mucosa is insufficient to absorb enough nutrients or water to sustain life. Even though most cases of SBS are acquired and not congenital, children still represent a significant portion of patients undergoing massive small bowel resection (mSBR), which is required to treat life-threatening cases of necrotizing enterocolitis, midgut volvulus, multiple atresias and gastroschisis. 3
After mSBR, the remaining bowel undergoes significant physiological changes to compensate for the increased functional load it must manage. This process is known as intestinal adaptation and is accompanied in the clinic by enteral nutrition therapy to ensure that the SBS patient will meet minimum fluid and nutritional requirements. As intestinal adaptation progresses to a life-sustaining level, the patient is weaned off the enteral therapy. If the adaptation response is incomplete, the patient will be committed to a lifetime of total parenteral nutritional (TPN) support, which involves direct injection of liquid nutrients into the bloodstream via catheter in a large vein, bypassing the gastrointestinal tract completely; this treatment is not only physically limiting and inconvenient but is also associated with significant risk of infection, liver damage and hyperglycemia. 4 Thus, understanding the complex process of resection-induced adaptation is necessary to identify molecular targets that can be clinically modulated to magnify this response.
Several prior studies have investigated resection-induced adaptation in the intestinal mucosa. Indeed, distinctive morphological changes have been characterized in crypt and villus units during successful adaptation of nutrient absorption. In addition to the mucosal layer, adaptation involves all layers of the bowel wall, including the underlying intestinal smooth muscle (ISM). 5 The contractile function of the ISM plays a central role in the regulation of intestinal transit. In contrast to what is known regarding mucosal adaptation, resection-induced changes in morphology, proliferation and contractility of the ISM, and its contribution to the adaptive response, are largely unknown.
The purpose of the present study was two-fold. We first aimed to elucidate the temporal profile of resection-induced changes in the ISM. Specifically, we assessed the temporal contractile properties of the ISM after resection in response to contractile agonists. We next sought to determine the mSBR-associated alterations of M2 muscarinic acetylcholine receptors (M2-mAChR) and M3-mAChR protein expression in the muscle layer of the rat ileum, which may represent future therapeutic targets of molecular therapies to alleviate SBS by promoting intestinal adaptation.
Materials and methods
Experimental design and animal model
Thirty rats were divided randomly into three experimental groups (n = 10, each). Group A rats underwent bowel transection and reanastomosis (sham). Groups B and C rats underwent an 80% SBR (SBS). Group A (sham) and group B (SBS1W) bowel tissues were harvested at day 7 postsurgery. Group C (SBS2W) bowel tissues were harvested at day 14 postsurgery.
Male Sprague–Dawley (SD) rats weighing 250–300 g were obtained from the Experimental Animal Center of Shanghai Jiaotong University School of Medicine (Shanghai, PR China). All experimental protocols included in this study were approved by the local Animal Care Committee and conformed to the Guide for the Care and Use of Laboratory Animals published by the Science and Technology Commission of the People's Republic of China (STCC Publication No. 2, revised 1988). All animals were housed in metabolic cages with free access to food and water, and acclimatized to their environment for five days before experimentation. Animals were maintained under standardized temperature, humidity and 12-h light–dark cycles. Animals were fasted for 16 h prior to laparotomy and intestinal surgery performed the following morning, as previously described. 6 All operative procedures were performed under anesthesia by intraperitoneal injection of pentobarbital sodium (30 mg/mL), which was administered at doses of 33–40 mg/kg body weight. During surgery, the abdomen was opened by a midline incision and the ligament of Treitz and ileal–cecal junction was identified and marked. For SBR rats, the enterectomy was performed by removing approximately 80% of the small intestine, leaving approximately 10 cm of the terminal ileum and 5 cm of the proximal jejunum, which were anastomosed. For sham-operated control rats, the laparotomy and all surgical manipulations were the same as above, but the resection procedure was not carried out. All animals received fluid resuscitation by intraperitoneal injection of saline (0.9% sodium chloride; 10 mL) before the abdominal wall was closed and surgery concluded. After recovery, rats were transferred back to individual cages and given water ad libitum overnight, after which a regular diet was reinstated.
Histological examination
Rats were euthanized at either day 7 or 14 postsurgery for tissue collection and analysis using standard procedures. 7 The 30-mm segment of mid-ileum was removed and opened along the mesenteric border. 8 Tissues were fixed in 4% paraformaldehyde and embedded in paraffin. Sections (5 μm) were stained with hematoxylin and eosin and examined using light microscopy. 9,10
Measurement of muscle contractile activity
To measure contractile activity, the intestine was opened to expose the mucosa and allow muscle strips to be excised. The length of muscle strips obtained was 15–20 mm, and the width was 2–3 mm. Circular muscle strips were then made from this tissue by suspending each sample along its circular axis in a bath of Krebs solution maintained at 37°C in an atmosphere of 95% O2 and 5% CO2. Muscle tension was measured isometrically under a resting tension of 1.5
Transmission electron microscopy
Full-thickness strips of the ileum from both control and mSBR rats were fixed with 2% paraformaldehyde, 2.5% glutaraldehyde, 3% sucrose and 1.25 mmol/L CaCl2 in 0.05 mol/L cacodylate buffer (pH 7.4) at 4°C for two hours. Tissues were subsequently washed in cacodylate buffer (4×15 min) prior to postfixation with 1% osmium tetroxide for one hour. After fixation, tissues were dehydrated through a graded series of alcohol, followed by immersion in propylene oxide for 10 min before being infiltrated and finally embedded in Eponate 12 resin (Ted Pella Inc, Redding, CA, USA). Semi- and ultra-thin sections were then cut parallel and transverse to the long axis of the circular muscle layer at different sites. Semi-thin sections were examined with a Leitz Diaplan microscope (Leitz, Wetzlar, Germany) using phase contrast. Ultra-thin sections were stained with uranyl acetate for 10 min and then counterstained with lead citrate for five minutes before being viewed with a Philips CM10 transmission electron microscope (Philips, Eindhoven, The Netherlands). 12
Western blotting
Muscle tissue samples were lysed in RIPA buffer (25 mmol/L Tris-HCl [pH 7.6], 150 mmol/L NaCl, 1% NP-40, 1% sodium deoxycholate and 0.1% sodium dodecyl sulfate). The protein concentration was then determined using the Bradford method and a commercially available kit (Pierce, Rockford, IL, USA). Tissue lysates (20 μg of total protein per lane) were separated by electrophoresis through a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis gel and transferred to nitrocellulose membranes (Hybond; Amersham, Piscataway, NJ, USA). The membranes were incubated with 5% milk to reduce non-specific antibody binding. Western blots were performed using antibodies directed against M2-mAChR (1:3000 dilution, mouse monoclonal M2-mAChR antibody; Thermo Scientific, Rockford, IL, USA), M3-mAChR (1:3000, rabbit polyclonal M3-mAChR antibody; Chemicon, Temecula, CA, USA) and glyceraldehyde-3-phosphate dehydrogenase (1:600, rabbit monoclonal antibody; CW Biotech Co, Shanghai, China). Alkaline phosphatase (AP)-conjugated secondary antibodies (CW Biotech Co) were used to detect protein bands.
Statistical analysis
The results are presented as means ± standard error (SE). Statistical differences between the groups were determined by using a one-way analysis of variance with the SigmaStat program (SPSS, San Diego, CA, USA). A P value less than 0.05 was considered statistically significant.
Results
Morphological changes in small intestine
In this study, we first examined the morphological changes that occurred in the small intestine following mSBR. The results indicated that villus height and crypt depth increased significantly in the remnant ileum of mSBR rats (Figure 1), as compared with that in control rats. In addition, at the ultrastructural morphological level, the height and density of mSBR-associated microvillus were also ameliorated (Figure 2). These observations confirmed the morphological changes in intestinal mucosa that had been observed previously.
5
In the present study, however, we also found a remarkable thickening of the smooth muscle layers of the remnant small intestine of mSBR rats, as compared with control rats (Figures 1b and c).
Morphological changes in normal and short bowel syndrome (SBS) ileum. Hematoxylin and eosin-stained cross-sections of the ileum from the sham-operated control group (a) and the SBS1W (b) and SBS2W (c) groups. In the SBS intestine, there was a marked increase in the thickness of both the longitudinal muscle (LM) and circular muscle (CM) layers (scale bar: 100um). *P < 0.05, as compared with controls (n = 7). (A color version of this figure is available in the online journal) Ultrastructure of intestinal epithelial cells and smooth muscle cells in short bowel syndrome (SBS). The height and density of massive small bowel resection-associated microvillus were ameliorated (a). Muscle cells in advanced states of hypertrophy showed myofilaments and organelles typical of smooth muscle cells, but had more prominent mitochondria and smooth sarcoplasmic reticulum (b)
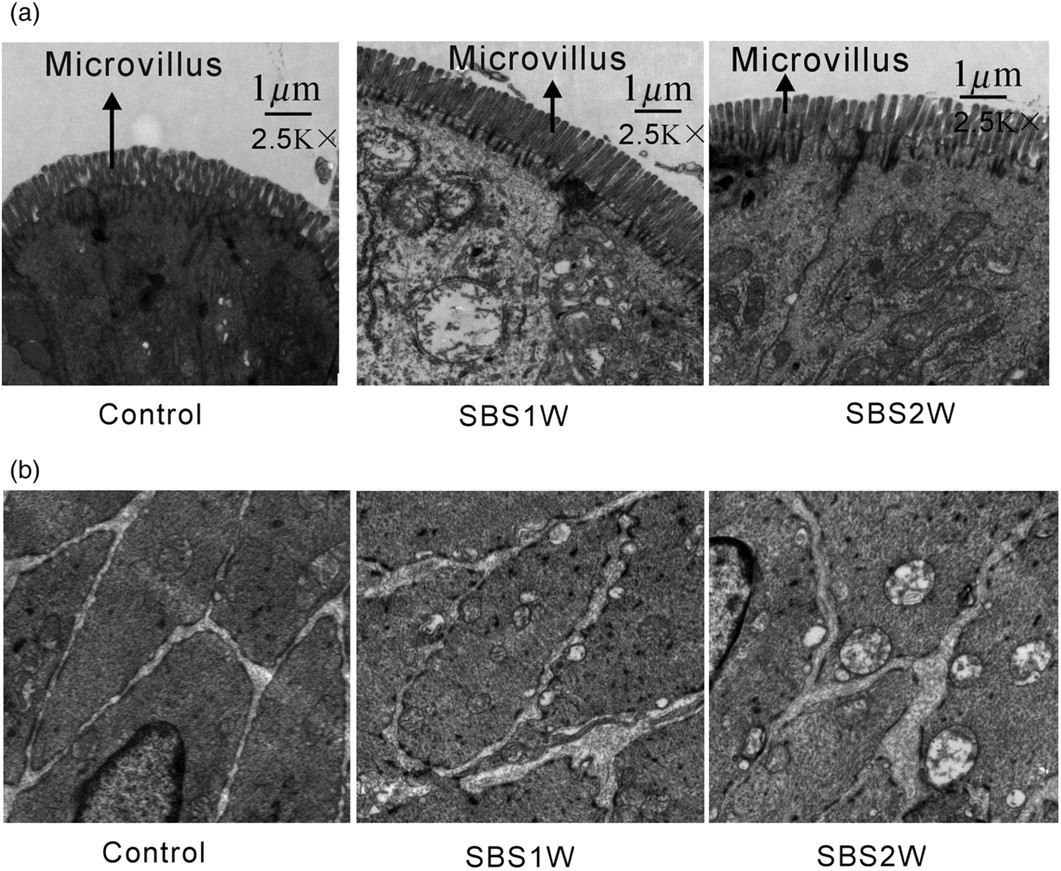
Morphological changes of the hypertrophic muscle cells
At day 7 after mSBR, hypertrophic muscle cells exhibited the normal structure of smooth muscle cells, including myofilaments and organelle composition. However, mitochondria and smooth sarcoplasmic reticulum appeared to be more prominent in the mSBR muscle cells than in the control muscle cells. Similar to the control muscle cells, mSBR muscle cells not only exhibited cisternae of reticulum beneath the cell surface and between the caveolae (Figure 2b), but also exhibited complex clusters of sarcoplasmic reticulum sacs near the cell surface (Figure 2b). In addition, the hypertrophic muscle cells showed large numbers of small sacs of sarcoplasmic reticulum scattered throughout the cytoplasm. Gap junctions were consistently observed in the circular muscle of the ileum from control rats. In the hypertrophic muscle cells, nexuses were numerous, possibly more numerous than in control muscle cells, and abutted neighboring muscle cells.
Adaptive changes of smooth muscle function
The function of smooth muscle was first detected for the control group for subsequent comparative analysis with mSBR-treated rats to determine functional changes in response to mSBR. It was found that circular muscles generated large and relatively regular spontaneous monophasical contractions with a frequency of 29.73 ± 2.67 events/min (n = 7). However, in mSBR rats, the spontaneous activity was found to be significantly altered at seven days after mSBR; the contractions showed an irregular pattern, were of low amplitude (Figure 3a), and had a frequency of 23.53 ± 7.83 events/min (n = 7; P < 0.05) (Figure 3b). By two weeks after mSBR, the functional activity of muscle cells was ameliorated significantly, and produced similar activity measurements as the normally functioning tissue from control rats with a frequency of 27.27 ± 4.18 events/min (n = 7; P > 0.05). Thus, mSBR-impaired muscular function was almost completely restored to normal after two weeks of recovery without intervention (Figure 3d).
Recovery of spontaneous rhythmic contractions of the intestinal circular muscle. The data illustrate typical traces of spontaneous contractions from individual non-resected (control) and massive small bowel resection (short bowel syndrome model) rats (a). Analytical data showing the frequency of spontaneous rhythmic contractions (b) and of contractions induced by different concentrations of carbachol in different rat model intestines (c). Concentration–response curves derived from contraction data of ileum strips from the control and SBS rat model in the presence of various concentrations of carbachol (d). *P < 0.05 (n = 7)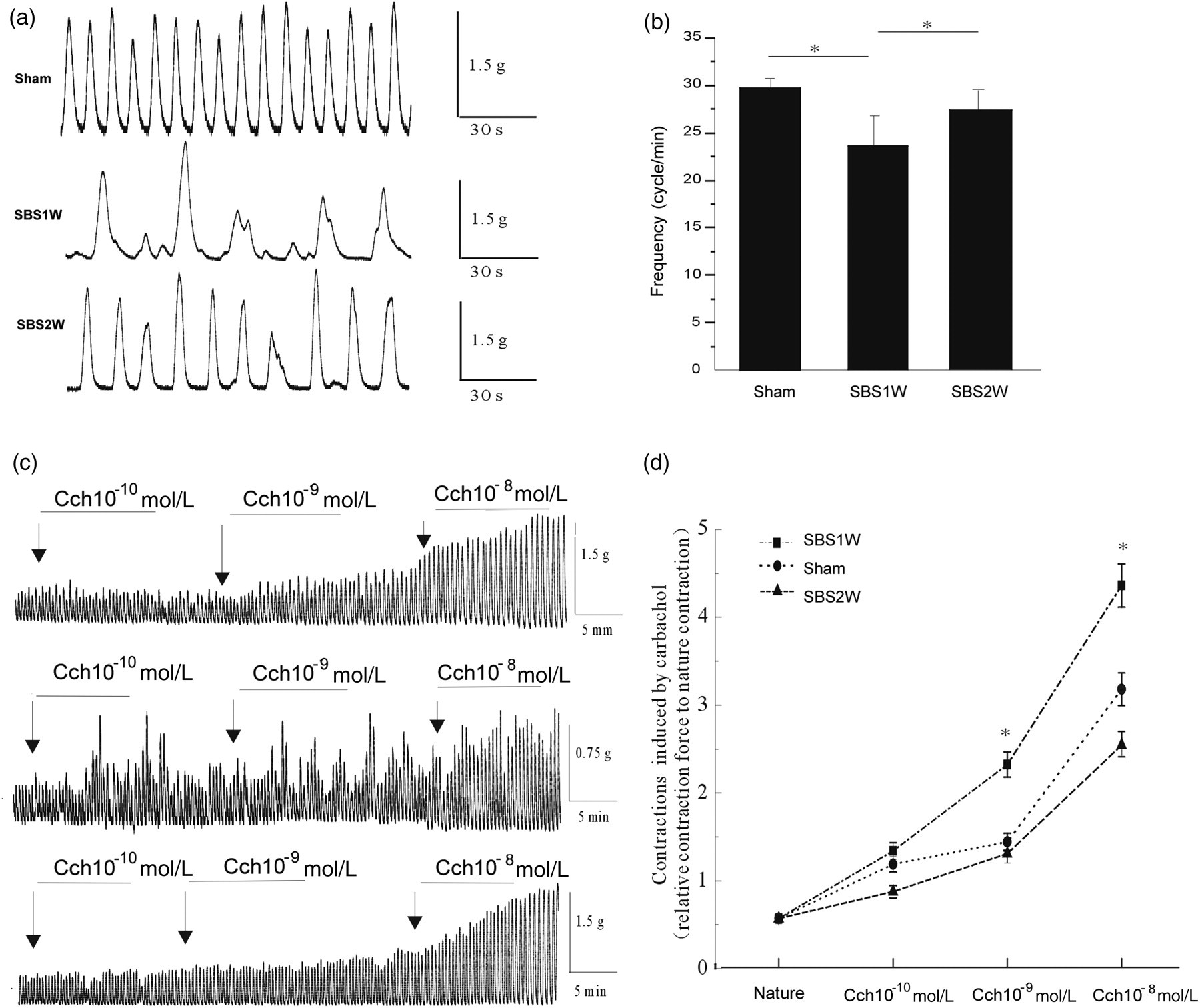
mAChR are involved in the recovery of contractile function of smooth muscle after mSRB
In order to investigate the roles of M2- and M3-mAChR in the recovery process of the contractile function of smooth muscle, we detected the protein expression of these two receptors in mSBR and sham rat ISM tissue by using Western blot (Figure 4a). Immunoreactivity for M2- and M3-mAChR were normalized to the internal control glyceraldehyde-3-phosphate dehydrogenase (Figure 4b) and compared between control and mSBR-treated groups. The expression of M2-mAChR was found to be significantly decreased by mSBR treatment, as evidenced by the low levels detected in the SBS1W group (n = 4; P < 0.05). The levels returned to baseline (as compared with control group) by day 14 after mSBR, when the contractile function had also been recovered. There were no statistical differences in the M3-mAChR levels in response to mSBR treatment or recovery of contractile ability. The results demonstrated that M2 and M3 muscarinic receptors can mediate neurogenic cholinergic contractions in rat ileum.
Changes in the expression profile of M2 muscarinic acetylcholine receptors (M2-mAChR) and M3-mAChR in response to short bowel syndrome (SBS) and intestinal adaptation. The expression of M2-mAChR was down-regulated in the first week after massive small bowel resection but increased by the following week (a, b). SBS did not produce a significant change in the expression of M3-mAChR at any time point examined (c, d). Relative expression was determined by normalization to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). *P < 0.05 (n = 4)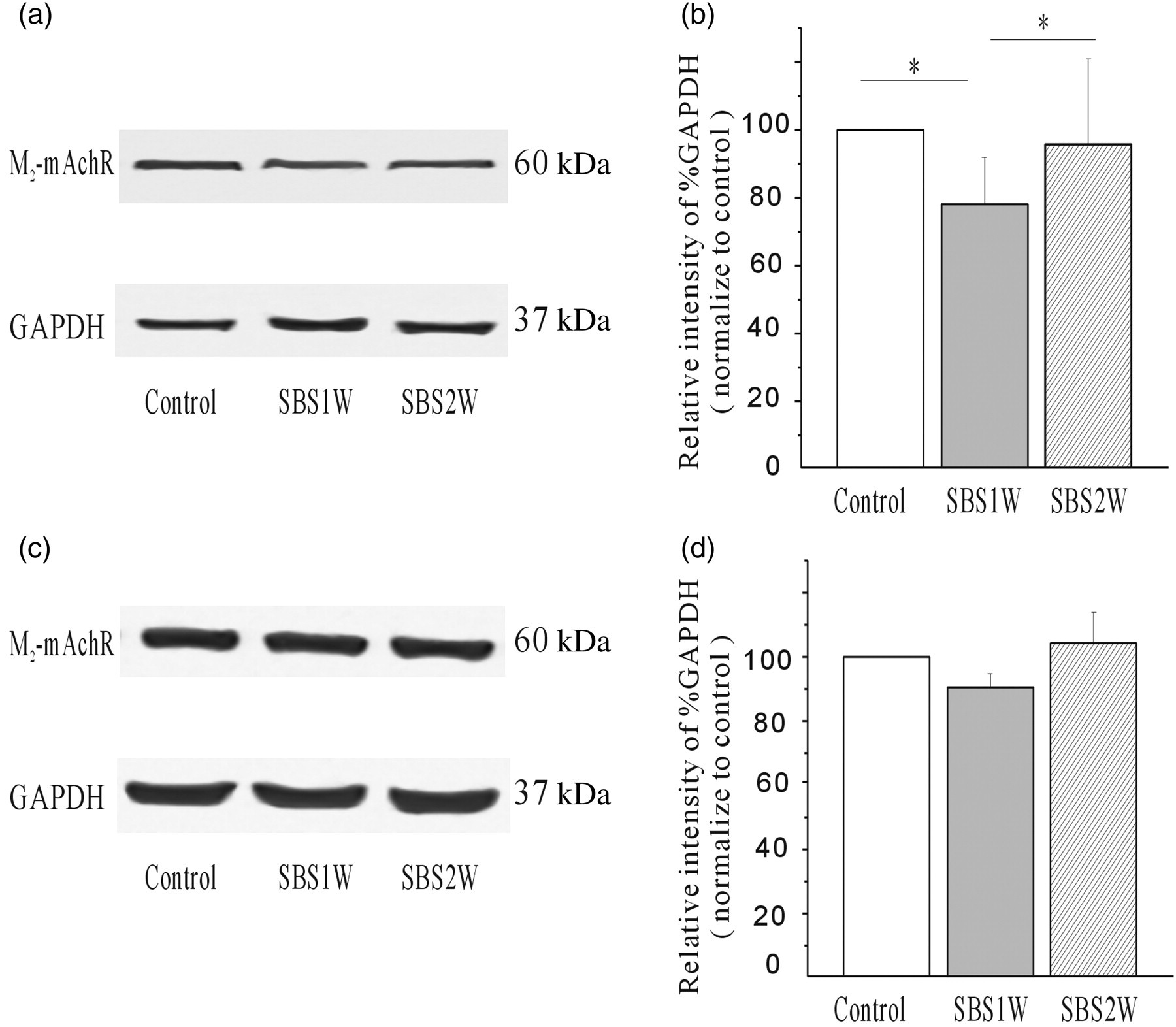
Discussion
Small bowel resection of more than two-thirds of the organ causes a pronounced and persistent transmural growth response in the remaining small bowel as the organ adjusts to meet the increased functional demands. The physiological adaptation process in both humans and rodents is characterized by dilation of the intestinal lumen, increased villus height, increased crypt depth and increased number of goblet cells. 13,14 In the current study, we confirmed that the resection-induced intestinal adaptation in a rat SBR model involves both the mucosal and ISM layers (Figures 1b and c). However, there is inconsistency in the literature over whether an adaptive increase in thickness of the smooth muscle also occurs after resection. Nguyen et al. 15 reported that ISM thickness significantly increased only after 50–75% mSBR. Some other reports demonstrated that intestinal resection caused no significant increases in the thickness of the smooth muscle layers in the remaining small bowel 16 after 75% mSBR in rats. Thus, it is import to investigate the adaptive function of muscle in the mSBR model. Currently, it is generally believed that all layers of the intestinal wall participate in the adaptive response. Martin et al. 5 measured changes in mucosal growth and intestinal caliber and length after mSBR in mice at postsurgery times similar to those used in our study. Their data indicated that ISM proliferation could possibly represent a passive response to mucosal growth. In our study, however, it was impossible to distinguish the primary event under the experimental conditions used. Nonetheless, we did observe that mSBR-treated rats exhibited a significant increase in the thickness of the smooth muscle layers in the remaining small bowel, as compared with controls (Figures 1b and c).
Extensive distal resection of the small intestine is known to produce distinct motility disorders in the intestinal remnant and to contribute to malabsorption and loss of nutrients by diarrhea. Short bowel syndrome is also characterized by intestinal inflammation that activates an inhibitory neuronal reflex pathway resulting in decreased motility early after surgery. 17,18 Martin et al. 4 were the first to recognize a temporal physiological response in contractility; specifically, increased contractile response was observed at the seventh day after SBR in mice. In our study, we extended the investigation of contractile function of smooth muscle tissues to determine the extent of functional recovery over time. Like the results from Martin et al., our mSBR rat model demonstrated significantly decreased frequency of contractility at day 7 after surgery, but we also found that the function was almost completely restored to normal after two weeks of recovery with no intervention.
In most species, cholinergic nerves play a crucial role in stimulatory regulation of smooth muscle activity and peristaltic movement. As cholinergic nerves are activated, the transmitter molecule acetylcholine (ACh) is released from their terminals. ACh then acts on smooth muscle cells to activate cell surface muscarinic receptors that mediate various intracellular signaling pathways leading to a rise in cytosolic Ca2+ concentrations. Ultimately, smooth muscle contraction is produced in the gastrointestinal smooth muscles. Intestinal smooth muscle cells are known to express five isoforms of the muscarinic receptor (M1–M5). 19 Of those, M2 and M3 are thought to be involved in contractile activity. 20,21 However, the role of muscarinic receptors in the pathogenesis of motility disorders of the ileum, especially after mSBR, has seldom been reported. 13,22
In this study, we examined the functional roles of M2 and M3 muscarinic receptors involved in ileal smooth muscle contractions. Our results showed that expression of M2-mAChR apparently decreased by approximately 24% at day 7, but M3 receptor expression had no significant change. It can be concluded that the shifting balance of M2- and M3-mAChR to M3-mAChR predominance may partially contribute to the increased contractile response. Previous studies have implicated a number of factors contributing to the pathogenesis of intestinal dysfunction. Although we found that muscarinic receptors and the ACh signaling pathway were involved in the recovery of contractile function, the precise mechanisms by which this occurs remains to be determined. It is possible that this effect is mediated by changes in the response of the smooth muscle itself, including modulation of receptor density or the signal transduction mechanism, or of particular related factors. Ongoing work in our laboratory is addressing this issue.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by grants from the Program for Innovative Research Team of Shanghai Municipal Education Commission and Special Foundation of Shanghai Municipal Public Health Bureau (LJ06021), the National Natural Science Foundation of China (30772270, 81000242) and the Scientific Foundation of Nantong University (10Z046).