
Abstract
Streptococcus pneumoniae resides on the mucosal surface of the upper respiratory tract and is ready to spread and trigger clinical diseases. Hence the vaccine that can eliminate the nasopharyngeal colonization was thought to be an ideal protective strategy against pneumococcal invasive diseases. Caseinolytic protease X (ClpX), a pneumococcal caseinolytic protease ATPase subunit, was shown to be a non-transmembrane protein by bioinformatics analysis. Consistent with the in silico prediction, the secretory expression of ClpX, instead of surface exposure, was further confirmed by flow cytometry and Western blot. Furthermore, ClpX was highly conserved in nine different serotypes of S. pneumoniae at both gene and protein concentrations. In addition, the anti-ClpX IgG antibody levels in human serum samples were much higher in healthy children, compared with pediatric patients, and displayed an age-related increase. Finally, ClpX protein antigen was introduced to BALB/c mice through a mucosal route, and protection against nasopharyngeal colonization and lethal infection caused by different S. pneumoniae serotypes was successfully elicited. Our findings suggest that ClpX is a potential candidate antigen that could be incorporated in pneumococcal protein vaccines.
Introduction
Streptococcus pneumoniae (pneumococcus) is a Gram-positive human pathogen which colonizes the mucosal surfaces of the host upper respiratory tract. It can be readily extended from carrier state to diverse, life-threatening, invasive infections such as bacteremia, pneumonia and meningitis for all age ranges, with children and the elderly at the highest risk. 1 This pathogen is responsible for an estimated 1.6 million deaths, including at least 1 million children under five years of age worldwide per year, causing great morbidity and mortality, and this situation is much worse in developing countries. 2,3 Moreover, as the aging population increases and the antibiotic resistance of S. pneumoniae develops, the need for a promising preventive strategy becomes a global problem of great urgency. 4
Currently available pneumococcal vaccines are all based on the serotype-specific capsular polysaccharides (CPS). The 23-valent polysaccharide pneumococcal vaccine (PPV23) contains free polysaccharides only and induces a T-cell-independent immune response, which is poorly immunogenic in children under two years old. Instead, the pneumococcal conjugate vaccines (PCV), including PCV7, PCV10 and PCV13, comprise covalently linked polysaccharide and protein and then turn to be T-cell-dependent antigens. 3,5 However, 93 distinct capsular serotypes have been identified until now. 6 Although these vaccines have greatly decreased the burden of pneumococcal disease, the limited serotype coverage can be an issue, making it essential to develop alternative strategies for better protection. 7
Pneumococcal protein-based vaccines that will protect against pneumococcal diseases have recently being investigated. Compared with the CPS-based vaccines, protein antigens can be produced in high quantity at low cost, and can induce T-cell memory responses. Besides, the elicited protections are well-conserved among diverse serotypes of pneumococcal strains. 8 Over the past years, considerable numbers of protein antigens have been well-studied in mice models, such as pneumolysin (Ply), 9 pneumococcal surface protein A, C (PspA, PspC) 10–12 and pneumococcal surface adhesion A (PsaA). 13 In these studies, protein antigens displayed an improved protection against nasopharyngeal colonization and lethal infection with one or more pneumococcal strains. However, many protein antigens exhibited polymorphism due to genetic diversity. 11,14 Thus, protein antigens that are well-conserved and could elicit effective protection are greatly needed.
Heat shock proteins (HSPs) are highly conserved proteins that can be elicited to protect eukaryotic and prokaryotic cells in stress condition. 15 Moreover, HSPs could stimulate both humoral and cellular immune responses and have manifested themselves to be potential vaccine candidates in various microbes. 16–18 Pneumococcal Clp (caseinolytic protease) proteins are members of the HSP100 family, and consist of a proteolytic subunit, ClpP, along with ATPase subunits (ClpX, ClpE, ClpL or ClpC), forming a functional Clp proteolytic complex. 19,20 It has been revealed that the ClpX subunit was an important virulence factor in the respect that the depletion of ClpX, instead of ClpP, ClpE, ClpL or ClpC, resulted in rapid cell death without overt cell morphology changes. 21 Although a previous study has reported the potential of ClpX as a subunit vaccine component of Brachyspira spp. for its conservation and immunogenicity, 22 no work has been done to characterize the immunogenic property and protective efficacy of ClpX in S. pneumoniae.
Recently, an ANTIGENome approach was prompted to screen pneumococcal vaccine candidates. It was suggested that a promising antigen should be surface-exposed and/or secreted. 2 Most currently, reported antigens were surface-exposed. Bioinformatic analysis implied the possibility of ClpX to be a secretory protein. In the present study, we verified the secretory expression of ClpX and then evaluated the capacity of intranasal immunization with recombinant ClpX (rClpX) protein to elicit cross-protection against nasopharyngeal colonization and lethal infection caused by different S. pneumoniae serotypes in mice.
Materials and methods
Mice
BALB/c female mice (6–8 weeks old) used in this study were purchased from, and raised at, Chongqing Medical University, Chongqing, China. All animal experiments were performed in accordance with the guidelines of the respective ethics committees of Chongqing Medical University.
Bacterial strains and culture
S. pneumoniae standard strains D39 (NCTC 7466, serotype 2) and R6 were obtained from the National Collection of Type Cultures (NCTC, London, UK), while S. pneumoniae clinical isolates CMCC 31109 (serotype 1), CMCC 31203 (serotype 3), CMCC 31446 (serotype 4), CMCC 31011 (serotype 5), CMCC 31207 (serotype 6B), CMCC 31507 (serotype 7F), CMCC 31614 (serotype 14) and CMCC 31693 (serotype 19F) were obtained from the National Center for Medical Culture Collections (CMCC, Beijing, China). Bacteria were grown on Columbia sheep blood agar or in C + Y medium. Escherichia coli stains DH5α and BL21 (DE3) (Novagen, Madison, WI, USA) were used for cloning and expression, respectively, and cultured in Luria–Bertani (LB) medium. Recombinants with pET-28a(+) expression plasmid were grown in LB medium supplemented with 50 μg/mL kanamycin.
Cloning, expression and purification of rClpX
ClpX primers (ClpX forward: 5′ GCCGC
Pneumococci surface staining
The surface staining of anti-ClpX antibodies to pneumococci was investigated by flow cytometry as previously described. 23,24 Briefly, S. pneumoniae strain R6 (non-encapsulated variant of D39 25 ) were collected and incubated with mice anti-ClpX antisera for one hour at 4°C, with mice anti-DnaJ antisera and preimmune serum samples as positive and negative controls, respectively. 23 Following this, the cells were washed twice by PBS and then stained with fluorescein isothiocyanate-conjugated goat anti-mouse IgG antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 30 min at room temperature in the dark. The cells were washed again, followed by flow cytometry analysis (Beckman Coulter, Brea, CA, USA).
Fractionation of S. pneumoniae
Fractionation was performed to analyze the localization of ClpX in S. pneumoniae. 26 S. pneumoniae D39 were cultured to OD600 of 0.5 in C + Y medium. After centrifugation, the culture supernatant was filtered and stored at −80°C in aliquots. The pellet was suspended in the protoplast buffer mixed with 25 μg mutanolysin (Sigma-Aldrich, St Louis, MO, USA) per mL and then incubated at 37°C overnight with shaking. The supernatant containing the cell wall proteins was collected, while the protoplast pellets were lysed in H2O, followed by centrifugation. The supernatant containing the cytoplasmic proteins were harvested, and the membrane proteins were solubilized in 0.5% Triton X-100. All the fractions were analyzed by Western blotting with anti-ClpX antibodies. The quality of fractionation was evaluated by assessing the activities of lactate dehydrogenase (GENMED, Arlington, MA, USA), which is known to be intracellularly localized. 27
Distribution analysis of ClpX by PCR and immunoblotting
To determine whether ClpX was conserved in different S. pneumoniae isolates, we tested the distribution of ClpX at different levels. S. pneumoniae standard strain D39 and clinical isolates CMCC 31109 (serotype 1), CMCC 31203 (serotype 3), CMCC 31446 (serotype 4), CMCC 31011 (serotype 5), CMCC 31207 (serotype 6B), CMCC 31507 (serotype 7F), CMCC 31614 (serotype 14) and CMCC 31693 (serotype 19F) were grown in C + Y medium and collected. PCR amplification was performed with specific primers to detect ClpX at gene levels, while Western blot analysis was used at protein levels.
Immunoreactivity of ClpX protein with human serum samples
Serum samples from 28 healthy adults were collected from the Blood Bank of Chongqing. Human immunodeficiency virus, hepatitis B virus or hepatitis C virus infections, acute diseases and severe chronic diseases were excluded from this study. Forty-nine serum samples from children aged from five days to eight years old (including 23 healthy children and 26 pediatric patients with acute pneumococcal pneumonia) were obtained from the Children's Hospital of Chongqing Medical University. This study was approved by the Ethical Committees of Chongqing Medical University, Children's Hospital of Chongqing Medical University and Blood Bank of Chongqing.
Sera anti-ClpX IgG levels were analyzed by enzyme-linked immunosorbent assay (ELISA). 10 μg/mL of purified rClpX was coated on 96-well plates and incubated at 4°C overnight. The plates were washed three times with PBS-0.1% Tween 20 (PBST) and then blocked with 2% bovine serum albumin-PBST for two hours at 37°C. After washing, serial dilutions of serum samples were added to the plates and incubated for one hour at 37°C. Bound immunoglobulin was detected by horseradish peroxidase-conjugated anti-human immunoglobulin G (IgG; Sangon Biotech, Shanghai, China), followed by the substrate 3,3′,5,5′-tetramethylbenzidine/H2O2. Absorbance was measured at 450 nm. The antibody titers were expressed as the reciprocal of the highest sera dilution, giving a 2.1-fold of the background absorbance.
Active immunization of mice
Six-to-eight-week-old female BALB/c mice were anesthetized with ethyl ether and then received into the nostrils 30 μL of PBS containing 20 μg of rClpX along with 1 μg of cholera toxin (CT; Sigma-Aldrich) once a week for three weeks. Mice receiving CT alone served as the negative control, while PPV23 (Chengdu Institute of Biological Products, Chengdu, China) served as the positive control.
Determination of humoral responses
One week after the last immunization, serum samples were collected from intranasally immunized mice. Blood was obtained by retro-orbital puncture. Serum samples were separated by centrifugation and stored at −80°C until analyzed. Sera IgG, IgG1, IgG2a, IgG2b, IgG3 and IgA concentrations were detected by ELISA as described previously with relevant secondary antibodies (Santa Cruz Biotechnology). Saliva was also collected from intranasally immunized mice. Briefly, mice were intraperitoneally injected with 0.25 mg carbamylcholine chloride (Sigma-Aldrich) per kg body weight. Approximately 50 μL saliva was obtained from each mouse with a sterile pipet tip, and then stored at −80°C until analyzed. ELISA was carried out for the measurement of saliva IgG and IgA levels as previously described.
Cytokine assays
Spleens collected from vaccinated or control mice were teased through stainless steel screens to obtain the separated cells. The splenocytes were then washed and resuspended in Dulbecco's modified Eagle's medium medium (Hyclone, Barrington, IL, USA) supplemented with 10% fetal bovine serum (5 × 106 cells/mL). Cells (1 mL) were cultured in 24-well plates and stimulated with rClpX at 37°C and 5% CO2. At the time of 12, 24, 48 and 72 h, the supernatants were collected and detected by ELISA kits (Biolegend, San Diego, CA, USA) for the secretion of interleukin (IL)-4, interferon-γ (IFN-γ) and IL-17A, following the manufacturer's instruction.
Challenge studies
For mouse nasopharyngeal colonization models, approximately 1.5 × 107 colony-forming unit (CFU) S. pneumoniae serotype 19F (CMCC 31693) cells were introduced intranasally to each immunized mouse (12 per group) 14 days after the last immunization. Five days postinfection, mice were sacrificed and nasal washes were harvested by flushing 300 μL of sterile PBS through the nasal cavities. Samples were serially diluted with sterile PBS and 100 μL was plated on Columbia sheep blood agar plates. The colonies were counted after incubation overnight at 37°C and 5% CO2.
For the mouse pneumococcal lethal intranasal challenge, all bacteria were passaged into mice intraperitoneally to enhance the virulence. BALB/c mice (12–15 per group) were anesthetized and then inoculated intranasally with 30 μL of bacterial suspension containing S. pneumoniae strains D39 (2 × 107 CFU) or CMCC 31614 (1.5 × 108 CFU), respectively. The survival of each mouse was monitored for 21 days.
Statistics
Antibody titers, cytokine levels of each time point and the numbers of pneumococci (CFU) were compared by the Mann-Whitney U test, and a statistical analysis of the survival rate was compared by log-rank test (Prism 5; Graphpad Software, La Jolla, CA, USA). P values of <0.05 were considered to be significantly different.
Results
Cloning, expression and purification of recombinant ClpX protein
The plasmid pET-28a(+) comprising the ClpX gene of S. pneumoniae D39 was constructed and then introduced into E. coli BL21(DE3). The recombinant ClpX protein was successfully expressed from the transformed cells, as the induced band corresponded well with its predicted size (46 kDa). The protein was predominantly expressed in soluble form. Approximately 18 mg of purified protein per liter of culture was harvested after purification. The purity of rClpX was >90%, as determined by SDS-PAGE with Coomassie blue staining (Figure 1).
SDS-PAGE analysis (10% gel) of the expression and purification of rClpX protein. Lane 1, lysate of untransformed Escherichia coli BL21(DE3) cells; lane 2, lysate of uninduced transformed cells; lane 3, supernatant of transformed cells induced by isopropyl β-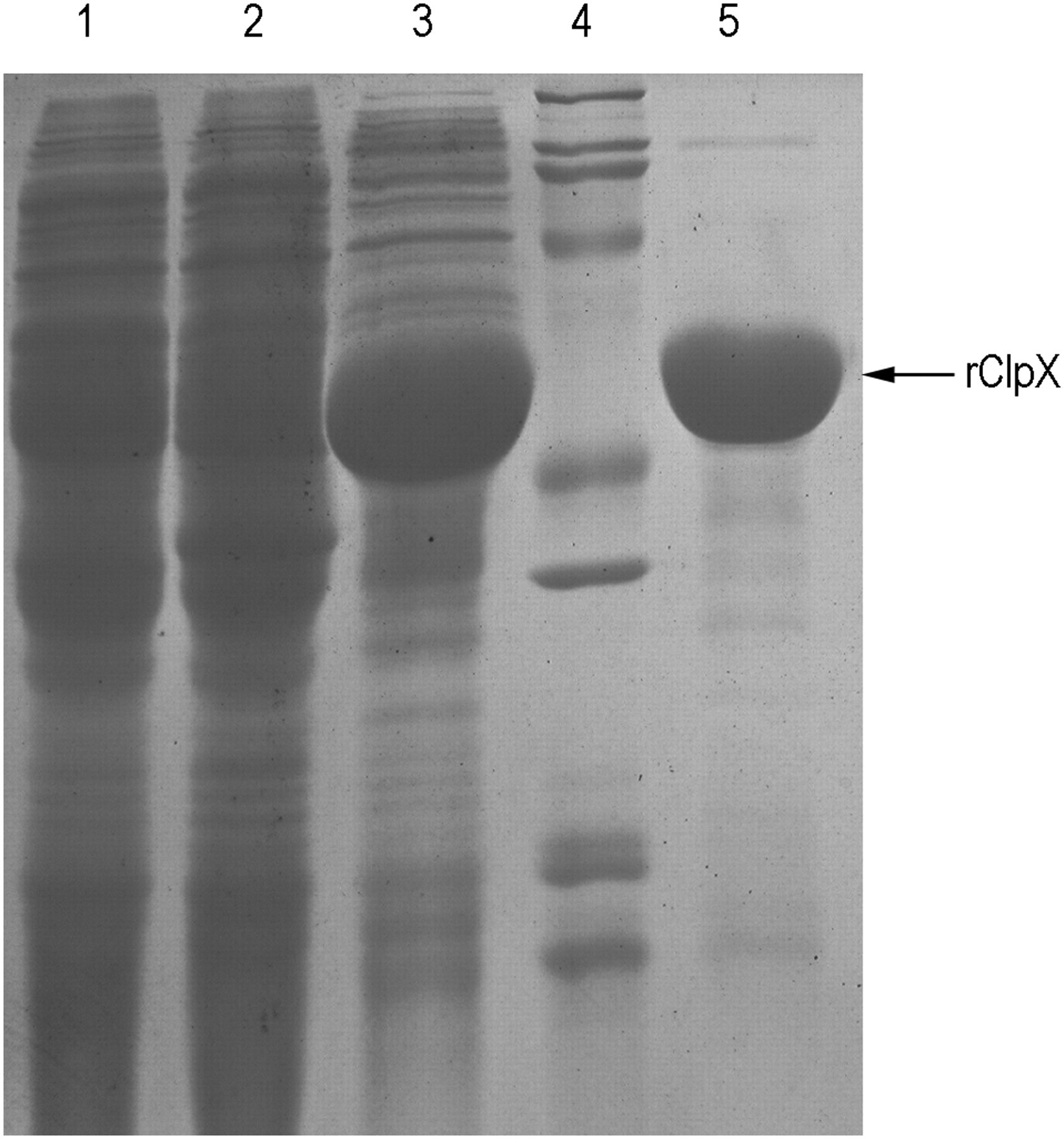
ClpX is a secretory protein localized in the cytoplasm of S. pneumoniae
It was predicted by TransMembrane prediction using Hidden Markov Models (TMHMM) that S. pneumoniae ClpX protein lacked transmembrane helices (Figure 2a), implying the possibility that ClpX is a cytoplasmic or secreted protein. To verify this prediction, surface staining with mice anti-ClpX polyclonal serum samples was detected by fluorescence-activated cell sorting analysis. No elevated level of fluorescence intensity was observed in the bacteria stained by anti-ClpX sera (Figure 2b), which suggested that ClpX would not be surface-exposed. To further explore the localization of ClpX protein in S. pneumoniae, cellular fractions were prepared by cell fractionation. Western blot analysis revealed that ClpX was present predominantly in the culture supernatant and cytoplasm of S. pneumoniae (Figure 2c), indicating the secretory expression of ClpX.
Localization of ClpX in Streptococcus pneumoniae. (a) Transmembrane prediction of ClpX by TMHMM. (b) Surface staining of S. pneumoniae R6/D39 with anti-ClpX sera and FITC-conjugated secondary antibodies. Preimmune serum samples were used as negative control, while anti-DnaJ serum samples as positive control. (b) Western blot analysis of cell fractions from S. pneumoniae D39. Lane 1, whole cells; lane 2, cytoplasm; lane 3, membranes; lane 4, culture supernatant fluid; and lane 5, cell wall. The blot was reacted with Anti-ClpX polycolonal serum samples. ClpX, caseinolytic protease X; TMHMM, TransMembrane prediction using Hidden Markov Models; FITC, fluorescein isothiocyanate. (A color version of this figure is available in the online journal)
ClpX is present in different strains of S. pneumoniae
The presence of ClpX in nine different S. pneumoniae strains detected in this study was demonstrated by PCR and Western blot analysis. With ClpX primers, each of the nine strains yielded an approximately 1233-bp band by PCR amplification (Figure 3a). Moreover, for the detection of ClpX expression in these strains, Western blot was probed with anti-ClpX polyclonal serum samples. The lysates of nine different strains were loaded respectively, and a specific band that migrated at approximately 46 kDa was observed in each line, which corresponded well with the size of the native protein (Figure 3b). The results above suggest that ClpX were highly conserved at the gene level as well as the protein concentration among the nine different strains.
Determination of the presence of ClpX in different Streptococcus pneumoniae strains by polymerized chain reaction (a) and Western blot (b). Lane M, DNA marker; lane 1, S. pneumoniae standard strain D39 (serotype 2); lane 2, CMCC 31109 (serotype 1); lane 3, CMCC 31203 (serotype 3); lane 4, CMCC 31446 (serotype 4); lane 5, CMCC 31011 (serotype 5); lane 6, CMCC 31207 (serotype 6B); lane 7, CMCC 31507 (serotype 7F); lane 8, CMCC 31614 (serotype 14); and lane 9, CMCC 31693 (serotype 19F). For Western blot analysis, rClpX protein was used as a positive control, while untransformed E. coli BL21 (DE3) as a negative control. No band was observed when preimmune mice serum samples were used (data not shown). ClpX, caseinolytic protease X; rClpX, recombinant ClpX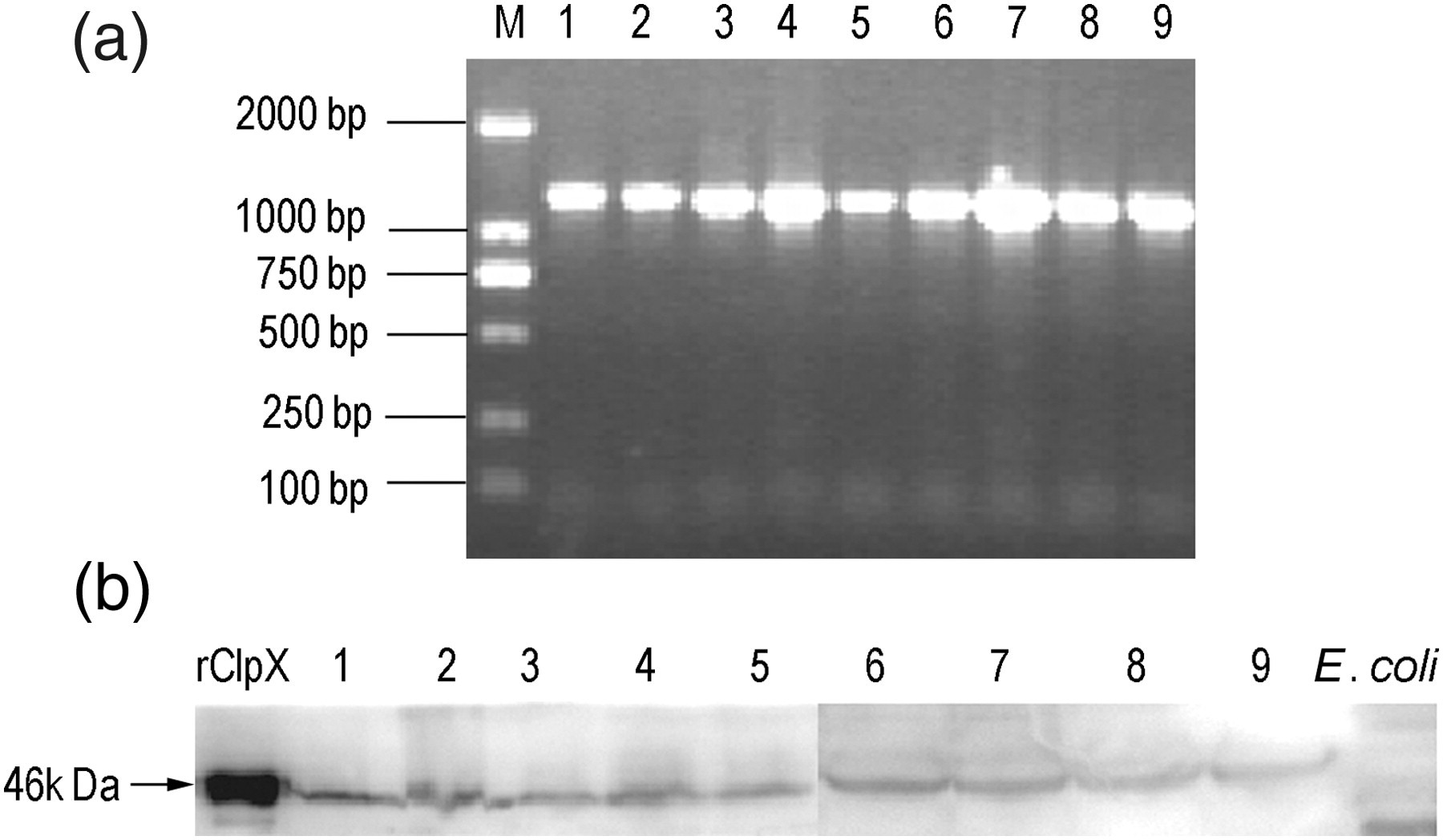
Levels of IgG antibodies specific for ClpX in pediatric patients with pneumococcal pneumonia, healthy children and naturally exposed adults
To study the human immune response against ClpX protein, human serum samples were collected and analyzed for the IgG titers specific for ClpX by ELISA. A broad range of IgG titers was observed in these groups (Figure 4). Although not statistically significant, the ClpX-specific antibody titers were lower in pediatric patients compared with the healthy children of the same age groups, especially in the 1–8-year-old groups (P = 0.31 for the <1-year-old groups and P = 0.08 for the 1–8-year-old groups). Furthermore, we observed a good correlation between the anti-ClpX antibody levels and the year of age, with significantly elevated antibody titers in adults (P = 0.007 and 0.27 for adults compared with healthy children <1-year-old and 1–8-year-old, respectively).
Enzyme-linked immunosorbent assay antibody titers against pneumococcal protein ClpX in human serum samples. Data are shown as median antibody titers, and each point denotes an individual's data. ClpX, caseinolytic protease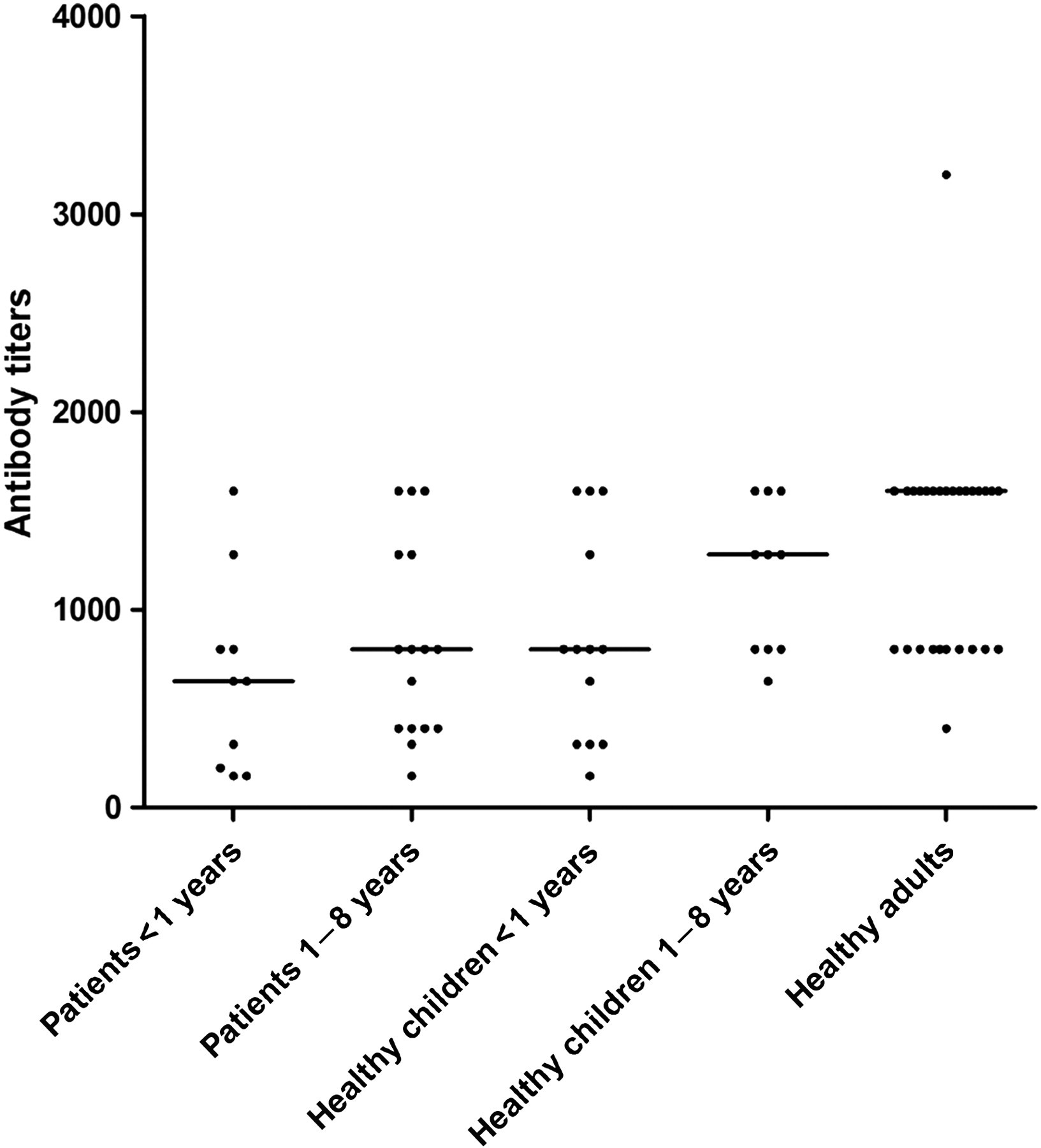
Antibody responses in ClpX-immunized mice
Intranasal immunization with rClpX in combination with CT adjuvant induced the production of IgG and IgA in both saliva and serum samples, while no detectable antibody response was observed in samples from mice immunized with adjuvant alone (Figures 5a and b). Moreover, serum IgG isotypes were analyzed to assess the type of immune response. The results revealed that both IgG1 and IgG2b responses were predominantly induced, indicating a Th2-biased immune response (Figure 5c).
Effect of mucosal immunization with rClpX on antibody responses in mice. (a) ClpX-specific IgG and IgA responses in saliva, with CT as adjuvant. (b) IgG and IgA responses in serum samples after intranasal immunization, with CT as adjuvant. (c) IgG isotypes in serum samples after ClpX immunization. The antibody titers were expressed as the reciprocal of the highest serum samples dilution, giving 2.1-fold of the background absorbance at 450 nm. **P < 0.01 compared with the adjuvant group. ClpX, caseinolytic protease X; rClpX, recombinant ClpX; CT, cholera toxin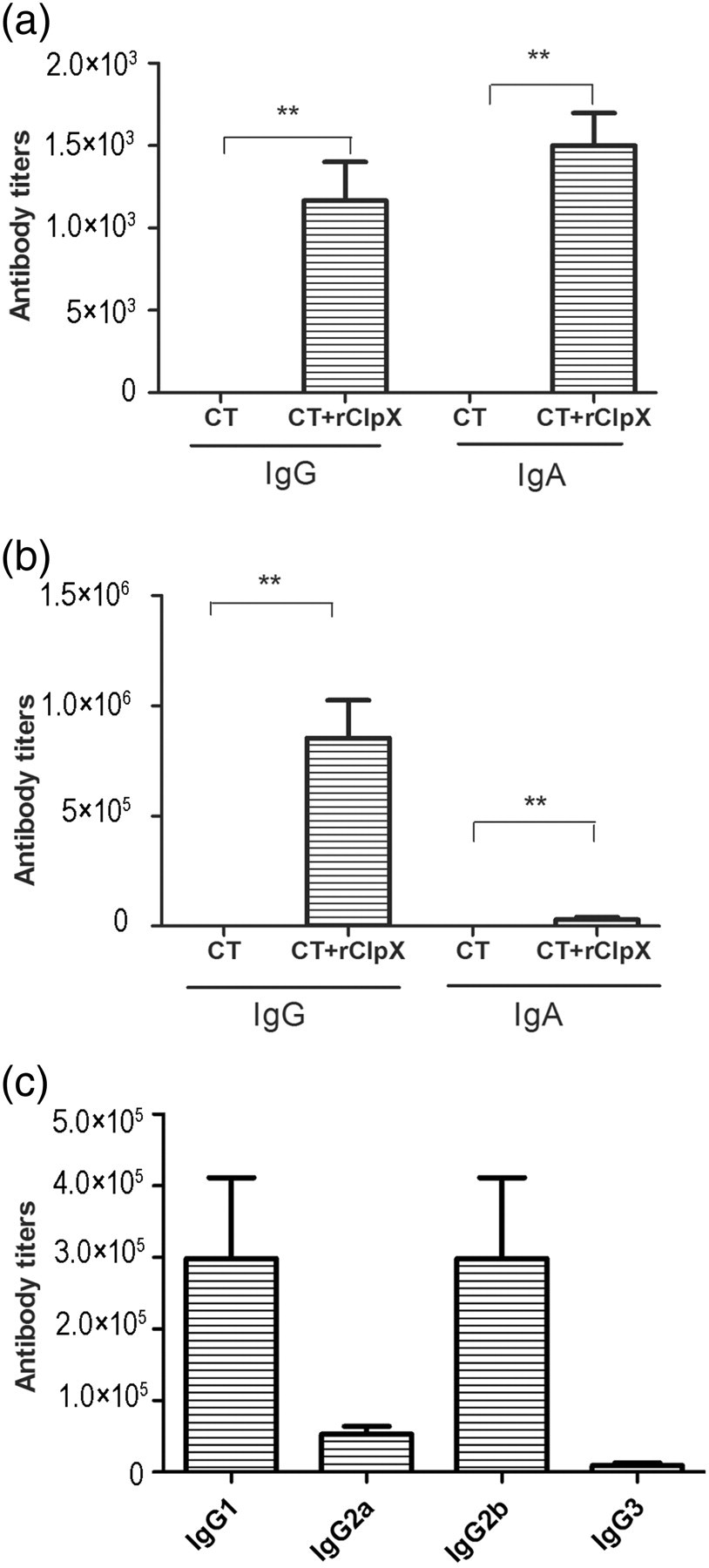
Cytokine responses in splenocytes of ClpX-immunized mice
We subsequently investigated the cytokine responses induced by intranasal immunization with rClpX. The splenocytes were harvested from control and immunized mice, followed by stimulation in vitro with rClpX (5 μg/mL). The levels of IL-17A, IL-4 and IFN-γ in the culture supernatant were analyzed. The results revealed that the levels of IL-17A and IL-4 secreted by splenocytes from the ClpX-immunized mice, which both peaked at the time of 72 h, were significantly higher than those from the control mice (Figures 6a and b). However, compared with the adjuvant control group, no significant upregulation of IFN-γ was triggered by ClpX immunization (Figure 6c).
Effect of mucosal immunization with rClpX on cytokine production in mice splenocytes. Splenocytes were harvested from mice immunized with CT + rClpX (solid line) or CT adjuvant alone (dashed line) for the detection of IL-17A (a), IL-4 (b) and IFN-γ (c) levels by ELISA. Data are shown as the mean value. *P < 0.05 for the ClpX-immunized group compared with the control. ClpX, caseinolytic protease X; rClpX, recombinant ClpX; CT, cholera toxin; IL, interleukin; IFN-γ, interferon-γ; ELISA, enzyme-linked immunosorbent assay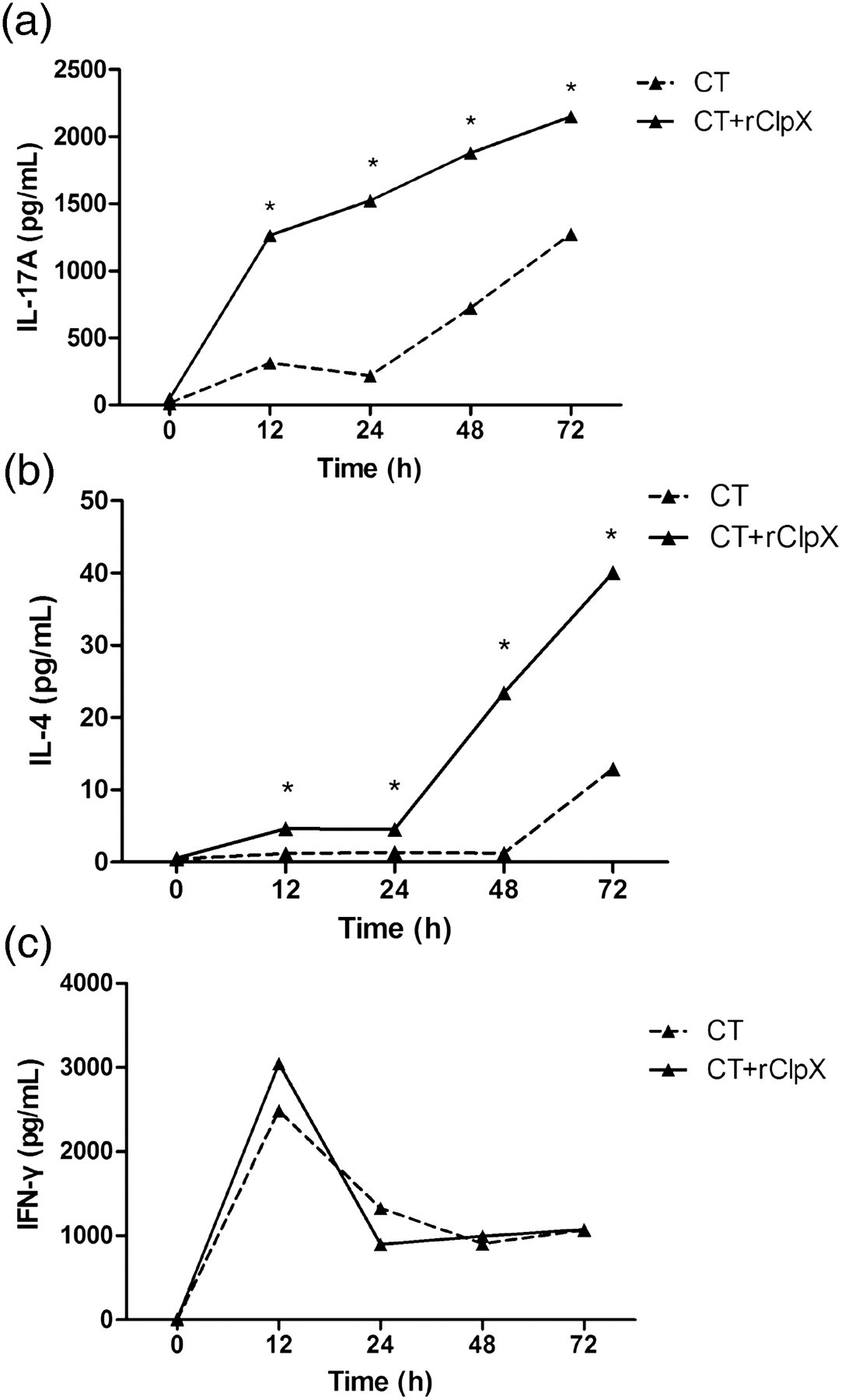
Protection against pneumococcal nasopharyngeal colonization by ClpX immunization
In order to investigate the protective effect of ClpX immunization against nasopharyngeal carriage, BALB/c mice were inoculated intranasally with S. pneumoniae serotype 19F (CMCC 31693) strain. We found that the inoculation dose of 1.5 × 107 CFU could trigger nasopharyngeal colonization without bacteremia five days later (data not shown), and approximately 10-fold reduction of bacterial loads was observed in nasal washes of mice immunized with rClpX compared with the control group (Figure 7).
Effect of mucosal immunization with rClpX in a pneumococcal nasopharyngeal colonization model. BALB/C mice immunized intranasally with rClpX mixed with CT adjuvant or CT alone (control) were challenged intranasally with pneumococcal serotype 19F strain (CMCC 31693, 1.5 × 107 CFU). Bacterial loads in nasal washes were determined five days postinfection. Each triangle represents one mouse. Horizontal lines denote the median CFU. ***P < 0.001 compared with the control group. rClpX, recombinant caseinolytic protease X; CT, cholera toxin; CFU, colony-forming unit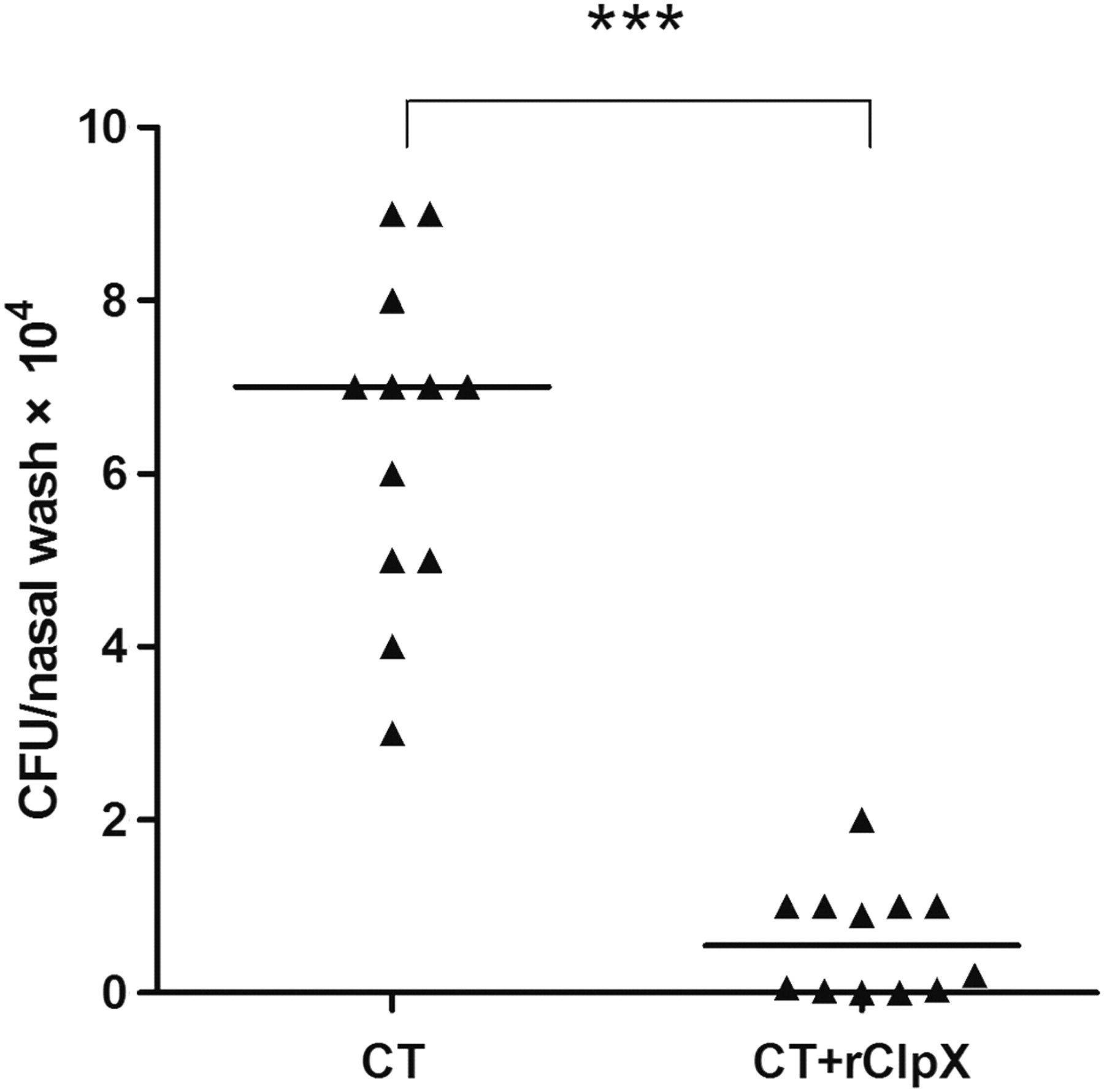
Protection against pneumococcal lethal challenge by ClpX immunization
Seven days after the last immunization, mice were challenged intranasally with pneumococcal serotype 2 (D39) and serotype 14 (CMCC 31614) strains, respectively, and mice survival was monitored for 21 days (Figure 8). For both pneumococcal strains, the survival rates of the ClpX-immunized mice (46.7% for D39 and 93.3% for CMCC 31614) were significantly higher than that of the control group (13.3% for D39 and 40% for CMCC 31614). Nevertheless, the protection efficacy of the ClpX immunization was not comparable with that of the PPV23 vaccination, which conferred 80% and 100% of mice survival against the D39 and CMCC 31614 strains, respectively.
Effect of mucosal immunization with rClpX on mouse survival against lethal intranasal challenge. BALB/c mice (n = 15) were immunized with rClpX mixed with CT adjuvant or CT alone (control), and then intranasally challenged with pneumococcal serotype 2 (D39, 2 × 107 CFU) (a) or serotype 14 (CMCC 31614, 1.5 × 108 CFU) (b), respectively. Mice vaccinated intraperitoneally with PPV23 were conducted as positive control. *,** indicate P values of <0.05 and <0.01, respectively, compared with the control group. rClpX, recombinant caseinolytic protease X; CT, cholera vaccin; CFU, colony-forming unit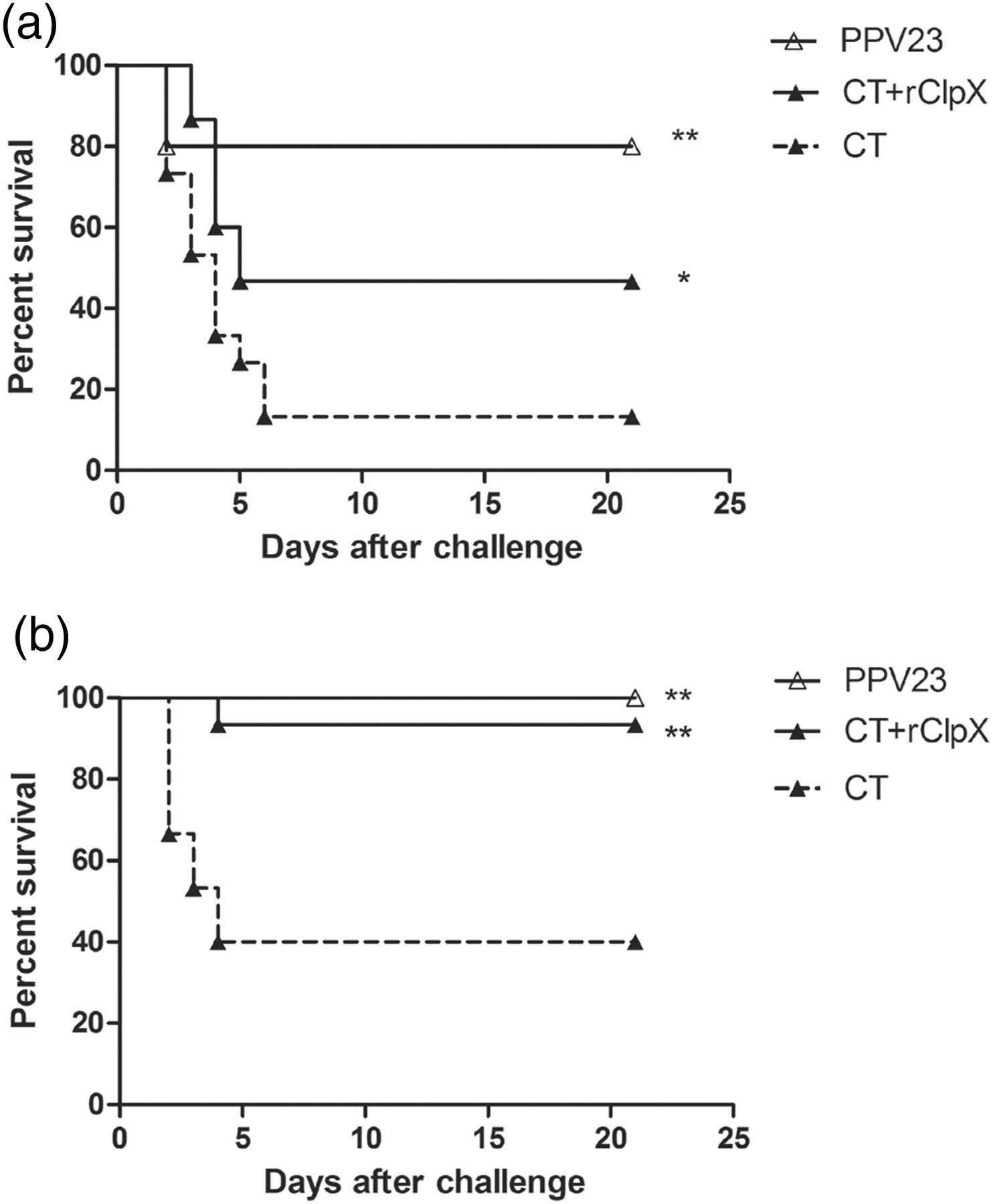
Discussion
Vaccination is an efficient and cost-effective strategy to decrease the morbidity and mortality of infectious diseases. 28 Numerous studies have been carried out in the development of pneumococcal vaccines, and research results suggest that the inoculation route and candidate antigen are important factors which should be taken into consideration. 1,8,29
Asymptomatic upper respiratory tract colonization is not only a prerequisite of S. pneumoniae to extend to relative sterile places and cause invasive infections, 30–32 but also an essential step for horizontal transmission within the community. 33,34 Therefore, it is assumed that vaccines that can eliminate the nasopharyngeal colonization could contribute to the protection against invasive infection and trigger herd immunity (indirect protection of non-vaccinated individuals). Two elements have been reported to be closely related to the S. pneumoniae nasopharyngeal colonization: Th17-mediated immunity was considered to have the potential to remove the pathogens from the mucosal surface, 35,36 while a study conducted with IgA−/− mice suggested that specific secretory-IgA (S-IgA) antibodies were the crucial factors for the regulation of S. pneumoniae nasopharyngeal colonization. 37 It was evident that mucosal administration was able to elicit both Th17-mediated immunity and S-IgA, and mucosal immunization was considered as an alternative strategy. 38,39 In this study, the mucosal route was adopted for the immunization of ClpX. The results showed that both anti-ClpX S-IgA on the mucosal surface and IL-17A produced by splenocytes were significantly elevated, and the significant decreases in bacterial loads from nasal washes and the subsequent mortality may be due to the synergic effect of these two factors.
Pneumococcal protein antigens are regarded as the most encouraging approach for vaccine development to date. 1 Secretory proteins are promising protein vaccine candidates. They are readily available for processing and antigen presentation and therefore represent the primary antigen targets of the host immune response. 2,40 Various novel secretory proteins such as those produced by Mycobacterium tuberculosis, 41 Chlamydia muridarum 42 and Streptococcus suis 43 have been identified to be protective against bacterial infection, and Th1- and/or Th2-mediated immune responses are thought to be important arms of protection. However, the molecular mechanism of the protection induced by secretory proteins still needs further exploration. In this study, S. pneumoniae ClpX was identified as a secretory protein. Apart from the Th17-mediated immune response we mentioned before, intranasal rClpX immunization also effectively elicited an Th2-mediated immune response, as anti-ClpX IgG1 and IgG2b levels in the serum and the IL-4 level produced by splenocytes were specifically increased. Furthermore, anti-ClpX antibody was observed in human serum samples, and it was implied that the antibody titers related to the infection status and natural exposure of S. pneumoniae. In a word, mucosal immunization with secretory protein ClpX elicited robust Th17- and Th2-mediated immune responses, which form part of the repertoire of protection against S. pneumoniae.
To further assess the protective efficacy afforded by intranasal immunization with rClpX, we challenged the immunized mice through the intranasal route, which mimics the natural infection through aspiration in humans. 44 Besides the homotypic strain D39, two other S. pneumoniae serotypes isolates (serotype 19F and 14) that are prevalent in China were employed. 45 We noticed that intranasal immunization with rClpX could elicit cross-protection against nasopharyngeal colonization and lethal challenge effectively, which confirmed the potential of ClpX to be a pneumococcal vaccine candidate.
To date, no single protein antigen can elicit protection comparable to that obtained by CPS-based vaccines. As a result, combinations of virulence proteins have recently been explored. Nevertheless, not all combinations of protein antigens have afforded higher degrees of protection than single ones. Some formulations, especially the triple combination, have generated inferior protection. It was noted that all the employed subunit antigens were surface-expressed, and the antibody accessibility to S. pneumoniae surface may be impaired by steric hindrance or some unknown mechanisms. 46–49 A formulation comprising both surface proteins and secretory antigens may probably avoid this interference. In this study, S. pneumoniae ClpX was identified as a secretory protein. Mucosal immunization with recombinant ClpX was shown to prevent nasopharyngeal colonization and subsequent death caused by different S. pneumoniae serotypes, which suggests that pneumococcal secretory protein ClpX is a promising antigen to be comprised in the multicomponent pneumococcal vaccines.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by the National Natural Science Foundation grants of China (No. 30800016, No. 81000711 and No.81000766) and Foundation of National Key Discipline in Laboratory Medicine (No. 2010302).