
Abstract
Targeting the endosomal Toll-like receptors (TLRs) by specific agonists seems to be a promising tool for stimulation of the immunogenicity of dendritic cells (DCs). Since the functional outcome upon the engagement of TLRs may be different, the aim of our study was to examine if and how different concentrations of 7-thia-8-oxo-guanosine (7-TOG), a selective TLR7 agonist, influence differentiation, maturation and functions of human monocyte-derived DCs (MoDCs) and if its effects on MoDCs could be modulated by co-ligation of TLR3. Immature MoDCs were treated with different concentrations of 7-TOG (25, 100 and 250 μmol/L) alone, or together with polyinosinic:polycytidylic acid, Poly (I:C) (10 ng/mL), a selective TLR3 agonist, for an additional 48 h. We showed that the highest concentration of 7-TOG stimulated the differentiation, maturation and allostimulatory capability of MoDCs. These changes were accompanied by an increased production of interleukin 12 (IL-12) and induction of T helper (Th)1 and Th17 immune responses. Both Th responses were significantly augmented by additional stimulation of MoDCs with Poly (I:C). The treatment of MoDCs with the intermediate concentration of 7-TOG resulted in the up-regulation of co-stimulatory molecule (CD86) and increased production of IL-1β and IL-6 by MoDCs, followed by the stimulation of the Th17 immune response. The lowest concentration of 7-TOG down-regulated the expression of CD40 on MoDCs and potentiated the Th2 immune response. The Th2 response was not significantly modulated by additional treatment of MoDCs with Poly (I:C), but this combination of TLR3/TLR7 agonists also stimulated both Th1 and Th17 responses. In conclusion, our results show that 7-TOG influences the phenotype and functions of MoDCs in a dose-dependent manner and suggests that fine-tuned signaling through TLR7 may be modified by the engagement of TLR3, resulting in a different outcome of immune response.
Keywords
Introduction
Toll-like receptors (TLRs) serve as sensors which recognize a number of pathogen-associated molecular patterns of invading pathogens and thus represent a significant component within the first line of defense against infections. 1 These receptors are predominantly expressed by the cells of innate immunity, especially dendritic cells (DCs). DCs are essential in initiation of the immune responses. 2 It has been shown that ligand recognition by TLRs promote DC maturation and induce DCs to secrete cytokines and chemokines and stimulate their capability to control T helper (Th)1, Th2 and Th17 cells and regulatory T (Treg) cell differentiation and function. 3,4 A number of studies indicated that specific TLR agonists stimulate the immunogenic properties of DCs, much better than a well-known cocktail of proinflammatory mediators. 5,6
TLR7, together with TLR3, TLR8 and TLR9, represents a group of antiviral TLRs which are localized within endosomal compartments. 1 TLR7 and TLR8 are structural homologs and natural receptors for single-stranded RNA (ssRNA). 7 Besides ssRNA, several chemical compounds have been described as TLR7/8 agonists, such as resiquimod, CL075. 8 Stimulation of monocyte-derived (Mo) DCs with these synthetic compounds resulted in their maturation and polarization of the immune response towards Th1. 9 It has been shown that strong stimulation of the Th1 immune response is a prerequisite for antitumor immunity. In this context, compounds inducing strong Th1 immunity, such as polyinosinic:polycytidylic acid (Poly (I:C)), a selective TLR3 agonist, have been extensively investigated as candidates for the preparation of DC tumor vaccines. 10 In the human system, TLR7 is strongly expressed on plasmacytoid DCs, whereas myeloid DCs express high levels of TLR8 and low levels of TLR7. 11 Therefore, to explain the mechanisms of MoDC activation based on the results using common TLR7/8 agonists, it was necessary to examine the effects of selective TLR7 and TLR8 ligands. Specific groups of imidazoquinoline (imiquimod and gardiquimod), as well as guanosine derivatives, including 7-allyl-7,8-dihydro-8-oxo-guanosine (loxoribine) and 7-thia-8-oxo-guanosine (7-TOG), are described as selective TLR7 agonists. 8,12 Their effects on human MoDCs are controversial. Most papers confirmed that selective TLR7 agonists stimulated MoDCs only if these cells were preactivated. 6,13 Zhu et al. 14 described that imiquimod inhibited the differentiation but enhanced the maturation of human MoDCs. In our previous paper, we showed that loxoribine was able to induce the maturation of MoDCs without additional stimuli and the effect was partly dependent on up-regulation of TLR7 expression. In addition, loxoribine-treated MoDCs were potent inducers of Th1 and Th17 immune responses. 15
7-TOG is a strong modulator of the immune response in rats. 16,17 Based on the difference in the expression of TLR7 in rodents and humans and the fact that the immunomodulatory effect of different TLR7 agonists may depend on the dose and its chemical structure, 8 the aim of this study was to investigate if and how different concentrations of 7-TOG influenced the differentiation and function of human MoDCs and if its effects on MoDCs could be modulated by co-ligation of TLR3.
Materials and methods
Medium and reagents
Human MoDCs were cultured in RPMI 1640 medium (ICN, Costa Mesa, CA, USA) supplemented with 2 mmol/L
Cell preparation and MoDC culture
MoDCs were generated from peripheral blood mononuclear cells (PBMCs), obtained from buffy coats of healthy volunteers (Institute for Transfusiology, Military Medical Academy, Belgrade, Serbia) by density gradient centrifugation (Lymphoprep, PAA, Vienna, Austria). The use of human cells for in vitro research was approved by the Ethical Committee of Military Medical Academy, Belgrade, Serbia. After isolation, PBMCs were re-suspended in 5 mL of RPMI-1640 containing 10% fetal calf serum and 2-ME and allowed to adhere to the plastic surface of flasks. Upon incubation of 90 min at 37°C, non-adherent cells were removed. To obtain MoDCs, adherent cells were cultured in 2 mL of culture medium containing GM-CSF (100 ng/mL) and IL-4 (20 ng/mL). On the third day, half of the medium was replaced with a fresh one (the same cytokine concentrations). On day 5, non-adherent cells (immature DCs) were re-plated (5 × 105 cells/mL) and stimulated with three different concentrations of 7-TOG (25, 100 and 250 μmol/L), either alone or together with Poly (I:C) (10 ng/mL). The control wells contained non-stimulated DCs. After 48 h, the cells were collected, washed and used for phenotypic and functional assays. Cell-free supernatants were collected and stored at −20°C for the subsequent determination of cytokine levels.
Flow cytometry
Control and treated MoDCs (1 × 105 cells/tube) were collected from culture wells, counted and washed in phosphate-buffered saline supplemented with 0.1% sodium azide and 2% FCS and incubated for 30 min at 4°C with one of the following monoclonal antibodies (mAbs) conjugated to either phycoerythrin (PE) or fluorescein isothiocyanate (FITC): HLA-DR-PE, CD40-FITC, CD86-PE and CD83-FITC (Serotec, Oxford, UK). Isotype controls were samples with irrelevant mouse mAbs, conjugated to PE or FITC (Serotec). Cells were analyzed using an EPICS XL-MCL flow cytometer (Coulter, Krefeld, Germany).
Allogeneic mixed leukocyte reaction
The ability of T-cells to proliferate was tested in an allogeneic mixed leukocyte reaction (MLR). CD4+ T-cells, used as responders in MLR, were purified from PBMCs by a CD4+ T-cell isolation kit (MACS Technology; Miltenyi Biotec, Bergisch Gladbach, Germany) following the instructions of the manufacturer. The purity of CD4+ T-cells, recovered in the negative fraction, was higher than 95%, as verified by staining with anti-CD4-FITC mAb (Serotec) and flow cytometry analysis. Purified CD4+ T-cells (1 × 105 cells/well) were co-cultured with MoDCs as stimulators in ‘U’-bottom 96-well microtiter plates in different DC:CD4+ T-cell ratios. After five days of cultivation, cell proliferation was assessed by pulsing the cells with [3H]-thymidine (1 μCi/well; Amersham, Buckinghamshire, UK) for the last 18 h of culture. Labeled cells were harvested onto glass fiber filters and radioactivity was counted in a scintillation counter (LKB-1219 Rackbeta; Wallac, Turku, Finland). The results are presented as the proliferation index (PI). PI was calculated as follows: PI = counts per minute (c.p.m.) (DC/CD4+ T-cell co-culture)/(c.p.m. [CD4+ T-cell culture alone] + c.p.m. [DC culture alone]).
A parallel set of cultures was used for the determination of Th cytokine profiles. After five days of cell cultures as described above, phorbol myristate acetate 20 ng/mL (Sigma) and ionomycin 500 ng/mL (Merck, Vienna, Austria), were added to the wells for an additional eight hours, after which the supernatants were collected.
Autologous T-cell activation
Th polarization capability of MoDCs was also evaluated using autologous naïve (CD45 RA+) CD4+ T-cells. Autologous CD4+ T-cells were isolated from PBMCs using the negative immunomagnetic sorting as described for allogeneic CD4+ T-cells. The sorted CD4+ T-cells were further separated using a CD4+CD45RA kit (MACS Technology; Miltenyi Biotec) and CD4+CD45RA+ cells were recovered in the positive fraction. The purity was higher than 95% as checked by using specific fluorochrome-labeled mAbs and flow cytometry. MoDCs (1×104 cells/well) were co-cultivated with CD4+CD45RA T-cells (1 × 105 cells/well) with the addition of antihuman CD3 mAb (UHCT1 clone; Serotec) at a concentration of 100 ng/mL for five days. After that, the supernatants were collected and frozen at −70°C until determination of cytokine levels.
Cytokine assays
The levels of IL-10, IL-12p70 (further in the text, IL-12), IL-23 and transforming growth factor-β (TGF-β) were measured in MoDC culture supernatants using sandwich enzyme-linked immunosorbent assays (ELISAs) from R&D Systems (Minneapolis, MN, USA) following the manufacturer's instructions. Concentrations of tumor necrosis factor-α (TNF-α), IL-1β, IL-6 and IL-8 were determined in the supernatants using commercial ELISA kits (eBioscience, Vienna, Austria). Levels of interferon-γ (IFN-γ), IL-17, IL-4 and IL-10 were measured in autologous and allogeneic CD4+ T-cell/DC co-culture supernatants using sandwich ELISA kits. Kits for IL-17 and IL-10 were purchased from R&D Systems, the IFN-γ kit was from Bender MedSystems, Vienna, Austria and the IL-4 kit was from Invitrogen Biosource, Carlsbad, CA, USA.
Statistical analysis
The significance between the means of experimental data was evaluated using one-way analysis of variance (ANOVA). P values less than 0.05 were considered to indicate statistical significance.
Results
Phenotypic characteristics of MoDCs treated with different concentrations of 7-TOG
Effect of different concentrations of 7-TOG on phenotypic characteristics of human MoDCs
Immature monocyte-derived dendritic cells (MoDCs) were cultivated with different concentrations of 7-thia-8-oxo-guanosine (7-TOG; 25, 100 and 250 μmol/L) for two days, as described in the Materials and methods section. After that, phenotypic characteristics of MoDCs were determined by flow cytometry
Results are given as mean % ± SD or mean fluorescence intensity (fl.) ± SD (n = 3 different donors)
*P < 0.05, **P < 0.01, ***P < 0.005 compared with the control
The treatment of MoDCs with the intermediate concentration of 7-TOG (100 μM) resulted only in the up-regulation of CD86. In contrast, the lowest concentration of 7-TOG (25 μM) slightly down-regulated CD40, whereas the expression of other examined markers was not significantly changed.
Effects of 7-TOG on the cytokine production by MoDCs
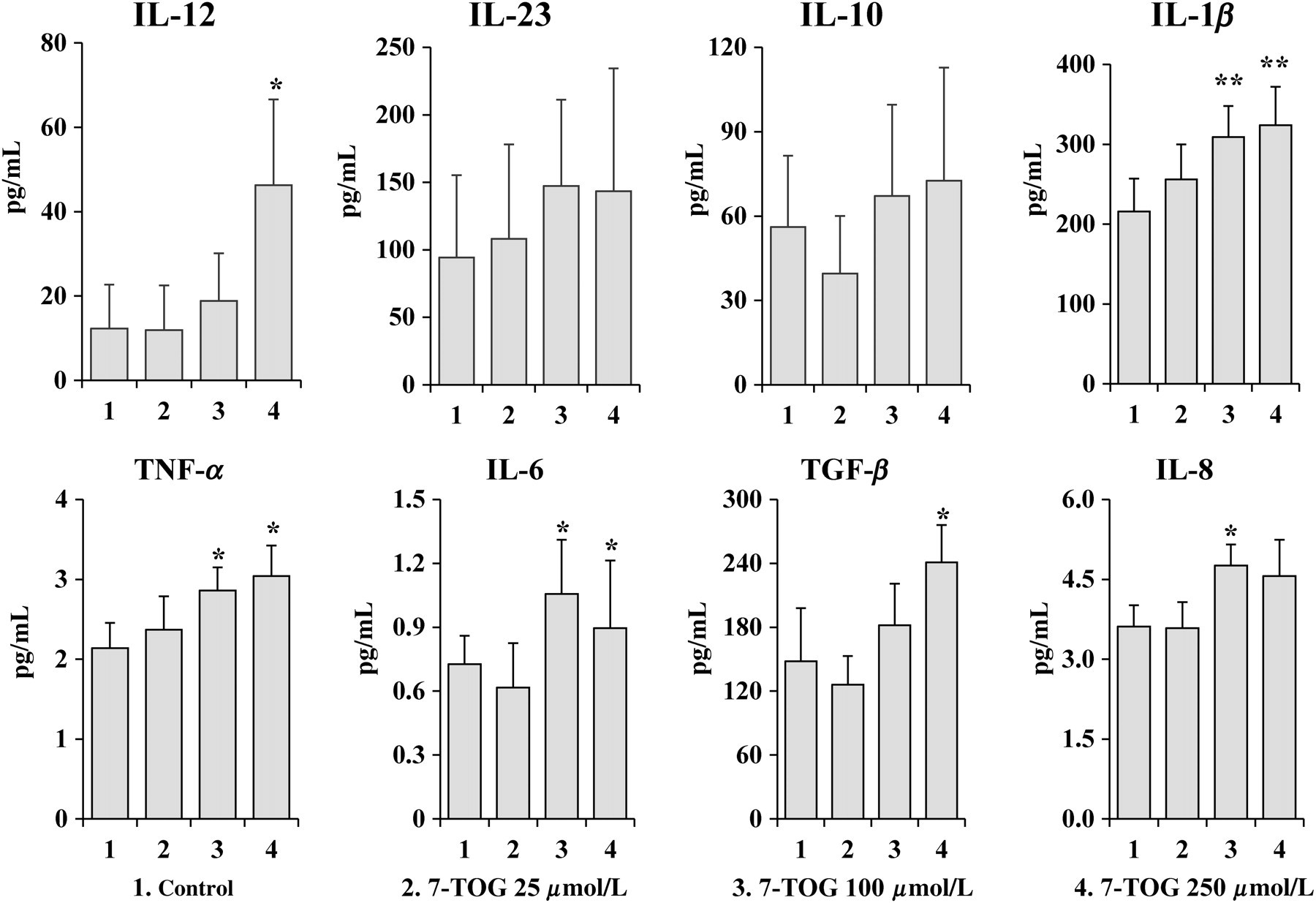
Cytokine levels were determined in culture supernatants of MoDCs. The highest concentration of 7-TOG up-regulated the production of IL-12, IL-1β, TNF-α, IL-6 and TGF-β by MoDCs compared with control, untreated MoDCs. The intermediate concentration of 7-TOG augmented the production of IL-1β, TNF-α, IL-6 and IL-8, whereas the lowest concentration of the agonist had no significant effect on the cytokine production by MoDCs (Figure 1).
Effects of different concentrations of 7-TOG on the cytokine production by MoDCs. Immature MoDCs were obtained by cultivating monocytes in the presence of GM-CSF (100 ng/mL) and IL-4 (20 ng/mL) for five days, when they were re-plated (5 × 105 cells/mL) and stimulated with three different concentrations of 7-TOG (25, 100 and 250 μmol/L). Levels of secreted cytokines were determined in cell-free supernatants by ELISA. Values are given as x ± standard deviation (SD) (n = 3 different donors). *P < 0.05, **P < 0.01 compared with the control. 7-TOG, 7-thia-8-oxo-guanosine; MoDC, monocyte-derived dendritic cell; GM-CSF, granulocyte/macrophage colony-stimulating factor; ELISA, enzyme-linked immunosorbent assay; IL, interleukin; TNF, tumor necrosis factor; TGF-β, transforming growth factor-β
Allostimulatory capacity of MoDCs cultivated with different concentrations of 7-TOG
The allostimlatory capacity of MoDCs was tested in a MLR, where purified CD4+ T-cells were used as responders. Figure 2 shows that only the highest concentration of 7-TOG enhanced the proliferative response of allogeneic CD4+ T-cells. The other two concentrations of 7-TOG did not have a significant influence on the allostimulatory capability of MoDCs, compared with control, untreated cells.
Allostimulatory capacity of MoDCs cultivated with different concentrations of 7-TOG. Double decreasing number of control MoDCs or MoDCs treated with different doses of 7-TOG were co-cultivated with allogeneic CD4+ T-cells (1 × 105 cells/well), as described in the Materials and methods section. Values are given as the proliferation index (PI) (mean ± standard deviation [SD]); (n = 3 different donors). The basal counts per minute (c.p.m.) in CD4+ T-cell cultures alone was 402 ± 53 c.p.m. The basal c.p.m. in DC cultures alone, independently of number and type of DCs, was between 62 and 102 c.p.m. (similar to the background radioactivity). *P < 0.05 compared with the control. 7-TOG, 7-thia-8-oxo-guanosine; DC, dendritic cell; MoDC, monocyte-derived DC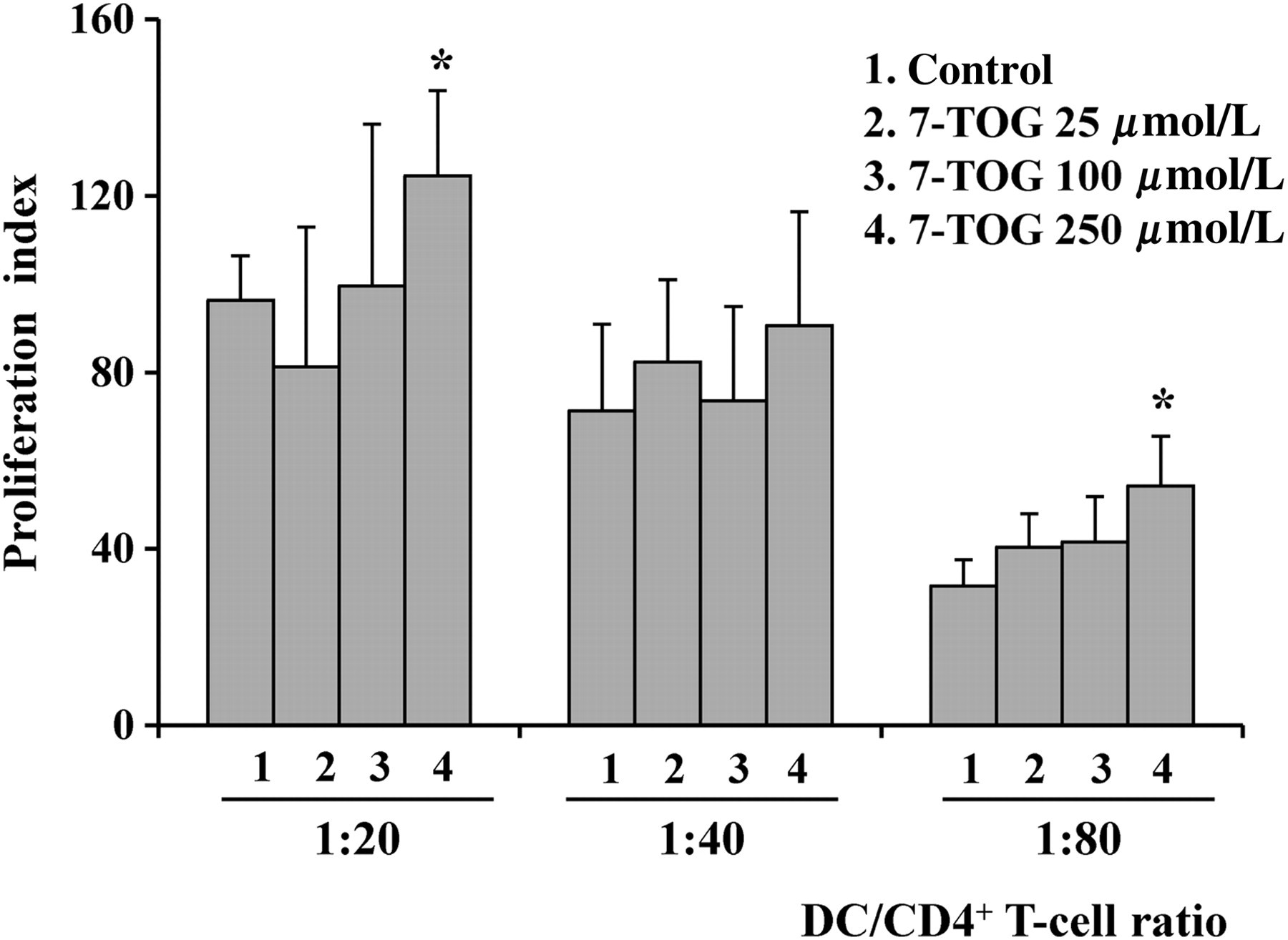
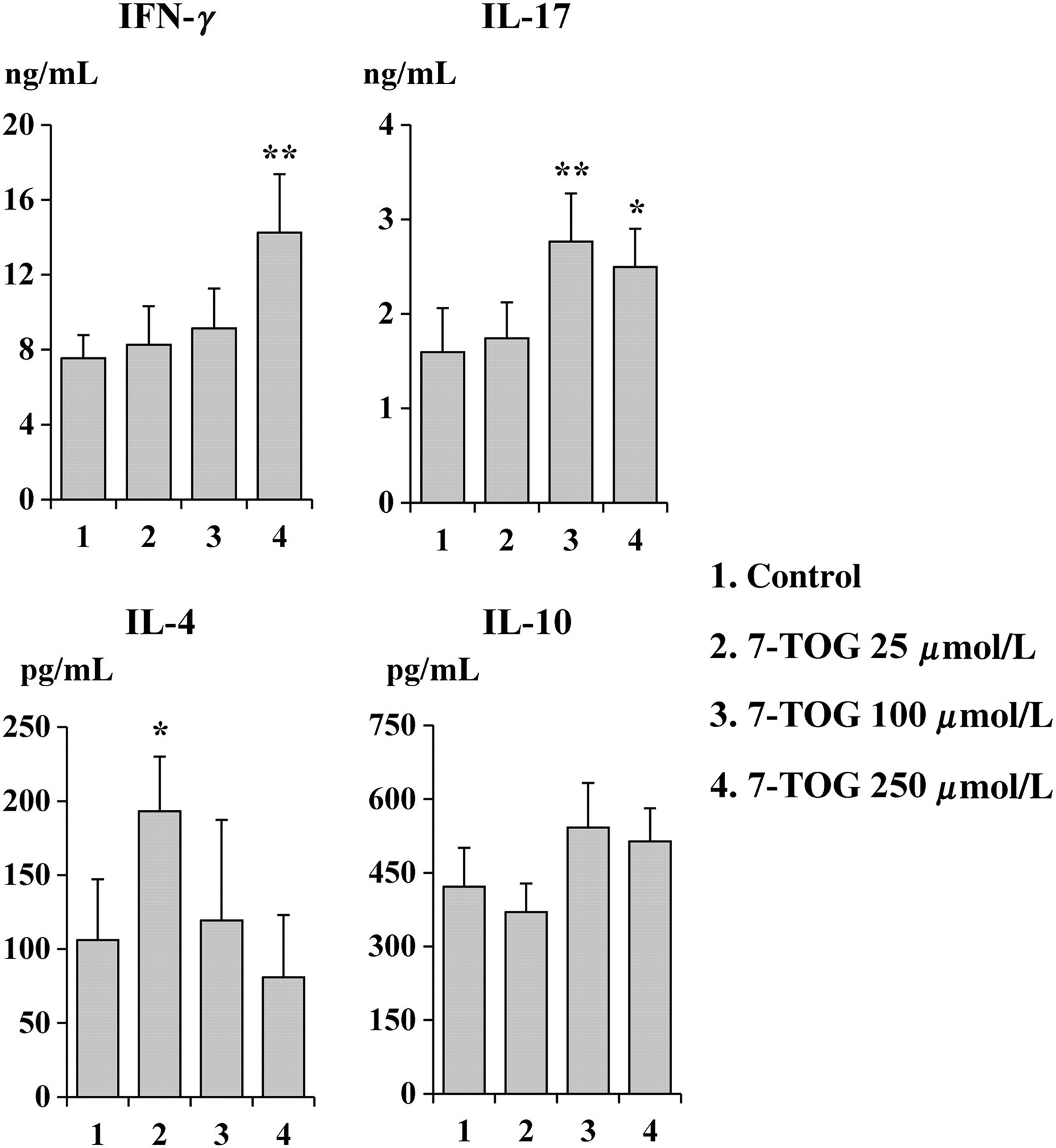
Effects of 7-TOG treatment on the Th polarization capability of MoDCs
The effect of 7-TOG on the Th polarization capability of MoDCs was firstly evaluated in the allogeneic DC/CD4+ T-cell co-culture, by measuring Th1, Th2, Th17 and Treg cytokine production. MoDCs cultivated with the highest concentration of 7-TOG increased the production of IFN-γ and IL-17 by allogeneic CD4+ T-cells, compared with the control MoDCs. A similar effect on IL-17 production was observed in the co-culture where MoDCs were treated with the intermediate concentration of 7-TOG. On the contrary, MoDCs treated with the lowest dose of 7-TOG, stimulated the Th2 immune response, as judged by increased production of IL-4.
The production of IL-10 in co-culture supernatants was not significantly modulated by MoDCs treated with any concentration of 7-TOG (Figure 3).
Effects of 7-TOG treatment on the Th polarization capability of MoDCs. MoDCs were generated as described in the Materials and methods section. Allogeneic CD4+ T-cells (1 × 105 cells/well) were co-cultured with MoDCs (1 × 104 cells/well) for five days, when PMA (20 ng/mL) and ionomycin (500 ng/mL) were added for an additional eight hours. After that, cell supernatants were collected and used for detection by ELISA. Cytokine levels are given as mean ± SD (n = 3 different donors). The levels of all cytokines in CD4+ T-cell (stimulated with PMA and ionomycin) cultures without MoDCs or MoDCs alone was less than 100 pg/mL (IFN-γ in CD4+ T-cell cultures) or undetectable. *P < 0.05, **P < 0.01 compared with the control. Th, T helper cell; 7-TOG, 7-thia-8-oxo-guanosine; MoDC, monocyte-derived dendritic cell; ELISA, enzyme-linked immunosorbent assay; PMA, phorbol myristate acetate; IL, interleukin; IFN-γ, interferon-γ
Th polarization of naïve autologous CD4+ T-cells by MoDCs treated with 7-TOG and Poly I:C
We further studied the Th polarization capability of MoDCs using naïve (CD45 RA+) autologous T-cells. Since MoDCs are not able to activate autologous T-cells without additional stimuli, we added an agonistic anti-CD3 mAb to the culture. Similarly, as in the allogeneic co-culture system, MoDCs treated with the lower concentration of 7-TOG (25 μmol/L) stimulated the production of IL-4, compared with control MoDCs. MoDCs pretreated with a higher concentration of 7-TOG (250 μmol/L) augmented the production of both, IFN-γ and IL-17 (Figure 4).
Combined effect of 7-TOG and Poly (I:C) on the capability of MoDCs to influence cytokine production of naïve autologous CD4+ T-cells in co-cultures. MoDCs were obtained by the cultivation of human monocytes for five days with GM-CSF (100 ng/mL) and IL-4 (20 ng/mL) and then stimulated with 7-TOG (25 and 250 μmol/L), Poly (I:C) (10 ng/mL) or their combination for an additional 48 h. Purified autologous CD4+CD45RA+ T-cells (1 × 105 cells/well) were co-cultivated with MoDCs (1 × 104 cells/well) with the addition of antihuman CD3 mAb (100 ng/mL) for five days. The levels of IFN-γ, IL-17, IL-4 and IL-10 were determined in cell-free supernatants of co-cultures using sandwich ELISA kits. Cytokine levels are presented as mean ± SD (n = 3 different donors). *P < 0.05, **P < 0.01, ***P < 0.005 compared with the control, untreated MoDCs. Δ
P < 0.05, ΔΔΔ
P < 0.005 compared with corresponding controls as indicated. 7-TOG, 7-thia-8-oxo-guanosine; MoDC, monocyte-derived dendritic cell; GM-CSF, granulocyte/macrophage colony-stimulating factor; ELISA, enzyme-linked immunosorbent assay; mAb, monoclonal antibody; IL, interleukin; IFN-γ, interferon-γ
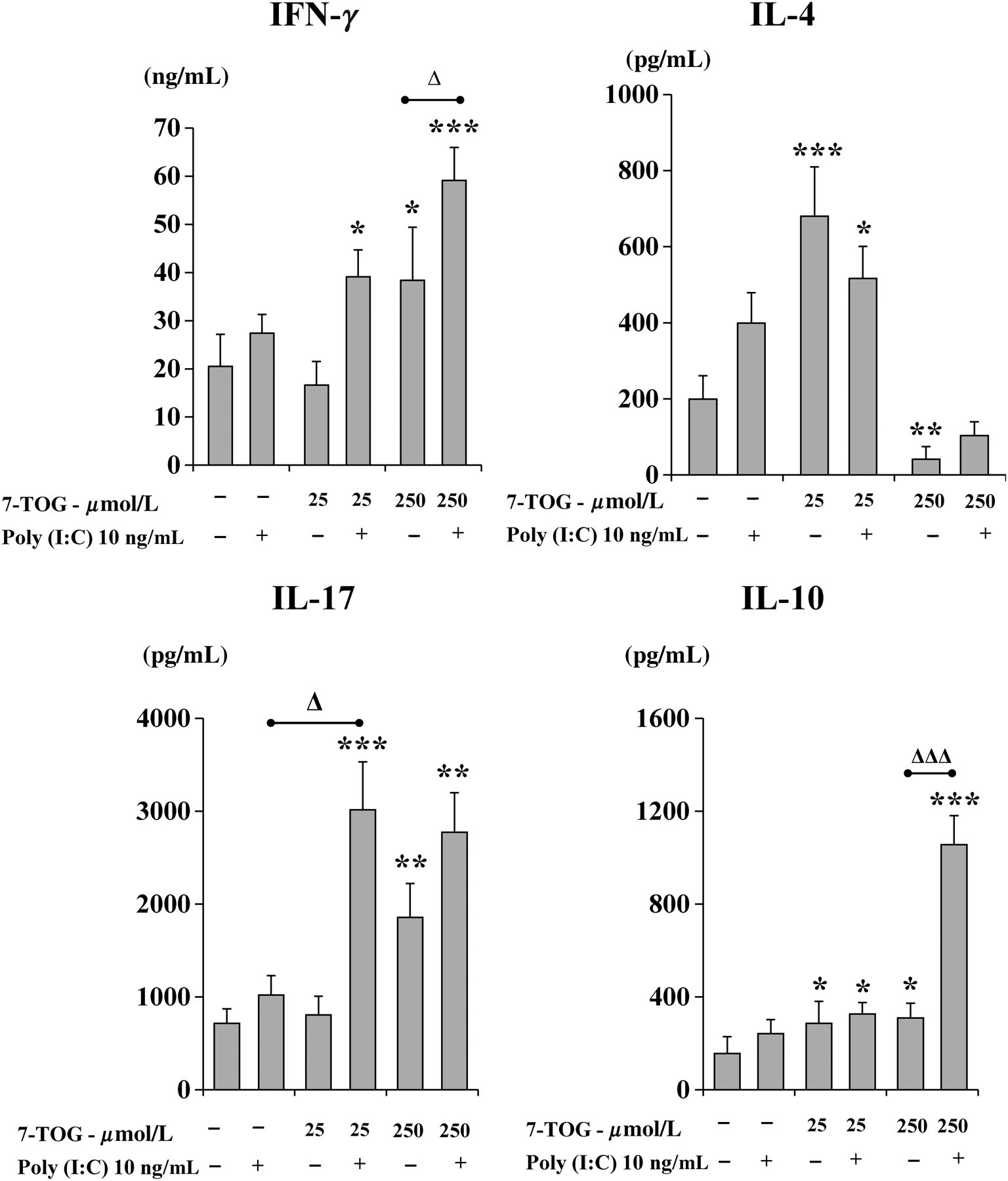
To assess how co-ligation of two investigated endosomal TLRs modify the function of MoDCs, we treated the cells with 7-TOG and Poly (I:C), a selective TLR3 agonist. As shown in Figure 4, a single concentration of Poly I:C (10 ng/mL) directed MoDCs to polarize the Th immune response predominantly toward Th1, as judged by the high level of IFN-γ in the cell co-culture. Co-stimulation of MoDCs with Poly (I:C) and a lower concentration of 7-TOG stimulated the production of IFN-γ and IL-17, compared with the effect of single agonists, whereas the production of IL-4 was not significantly modulated.
The production of IFN-γ and IL-17 was additionally augmented when MoDCs were treated with Poly I:C and a higher concentration of 7-TOG, compared with when MoDCs were stimulated with corresponding concentrations of single agonists, but this combination also significantly increased the production of IL-10 in the co-culture. In contrast, the production of IL-4 was down-regulated, similarly as in the co-culture with naïve CD4+ T-cells and MoDCs treated with a higher concentration of 7-TOG alone (Figure 4).
In order to analyze the relationship between Th polarization and production of cytokines by MoDCs, we measured the levels of IL-6, IL-10, IL-12, IL-23 and TGF-β in culture supernatants of MoDCs treated with the combination of 7-TOG and Poly (I:C), including the corresponding controls.
Poly (I:C) alone was a strong inducer of IL-12 production by MoDCs, whereas the production of other cytokines was not significantly modulated. The production of IL-12 and IL-6 by MoDCs was additionally augmented using Poly (I:C) and higher concentration of 7-TOG. Neither combination of these two TLR agonists significantly modified the production of IL-23, IL-10 and TGF-β (Figure 5).
Cytokine production by MoDCs simultaneously treated with 7-TOG and Poly (I:C). MoDCs were generated as described in the Materials and methods section and then stimulated with 7-TOG (25 and 250 μmol/L), Poly (I:C) (10 ng/mL) or their combination for an additional 48 h. Levels of secreted IL-12, IL-23, IL-10, IL-6 and TGF-β were determined in cell-free supernatants by ELISA. Values are given as mean ± SD (n = 3 different donors). *P < 0.05, ***P < 0.005 compared with the control, untreated MoDCs. Δ
P < 0.05 compared with corresponding controls as indicated. 7-TOG, 7-thia-8-oxo-guanosine; MoDC, monocyte-derived dendritic cell; ELISA, enzyme-linked immunosorbent assay; IL, interleukin; TGF-β, transforming growth factor-β

Discussion
The activation of DCs by targeting particular TLRs seems to be advantageous in stimulation of DC immunogenicity, compared with the use of cytokine cocktails. 5,6 In this context, we studied the influence of different concentrations of 7-TOG, a selective TLR7 agonist, on maturation and functions of human MoDCs, with the aim of improving the protocols for preparation of immunogenic DC-based cancer vaccines. TLR7 and TLR8 are members of the TLR subfamily that mediates the recognition of ssRNA in an endosomal compartment. 19 The signaling generated by the engagement of these TLRs are similar, but its functional outcome might be different, mostly due to their different expression in a particular DC type. 20,21 In our previous paper, 15 we showed that loxoribine (a guanosine analogue, acting as a selective TLR7 agonist 8 ), at relatively high concentrations (250 μmol/L), was able to induce a strong activation of MoDCs, and subsequent Th1 and Th17 immune responses, in spite of a low expression of TLR7 by MoDCs. 22 Using a structurally similar guanosine analogue, we showed that 7-TOG was also able to activate immature MoDCs, but its effect was strongly dose-dependent. The lowest concentration of 7-TOG (25 μmol/L) did not trigger maturation of MoDCs, but primed these cells to polarize the immune response toward Th2. The highest concentration of 7-TOG (250 μmol/L) was a potent inducer of MoDC maturation, directing the immune response toward Th1 and Th17. The intermediate concentration of 7-TOG (100 μmol/L) was a weak stimulator of MoDC maturation, but stimulated the Th17 response. Such a dose-dependent effect of TLR agonists on the Th polarization has not been published so far.
The induction of MoDC maturation by using the highest concentration of 7-TOG was documented on the basis of up-regulation of CD86, CD40 and CD83 on MoDCs. CD83 is a key DC maturation marker. 23 CD86 is a main co-stimulatory ligand for T-cells, providing the second signal for proliferation of antigen-specific T-cells. 24 On the other hand, increased expression of CD40 is significant for interaction with its ligand (CD40L) on T-cells, necessary for additional up-regulation of co-stimulatory molecules on DCs and their inforced capacity to stimulate proliferative responses. 25 Thus, the enhanced proliferative response of allogeneic CD4+ T-cells triggered by MoDCs, which were pretreated with the highest concentration of 7-TOG, may be explained by the influence of this TLR ligand on MoDC co-stimulatory functions. MoDCs treated with the highest concentration of 7-TOG stimulated the secretion of IFN-γ by allogeneic CD4+ T-cells. We obtained similar results using autologous naïve (CD4+CD45+RA) T-cells, a T-cell-subset which is recommended for Th polarization studies. 6,26 The production of IL-4, a Th2 cytokine, was not significantly decreased, but the ratio between IFN-γ and IL-4 was significantly higher in these cultures. The higher production of IFN-γ correlated with the levels of IL-12, TNF-α, IL-1β and IL-8. IL-12 is a key Th1-inducing cytokine. 27 The level of IL-12 in MoDC cultures treated with the highest concentration of 7-TOG was not as high as that trigerred by Poly(I:C), a TLR3 agonist, as shown in Figure 5 and previous studies, 28 but it was comparable with the concentrations of IL-12 following lipopolysaccharide stimulation. 29 One explanation of this finding could be that signaling through CD40–CD40L interactions during the contact of MoDCs with CD4+ T-cells additionally enhanced the Th1 immune response through induction of IL-12. 30 This mechanism may also be responsible for a stronger Th1 immune response using additional stimulation of MoDCs with Poly (I:C), as judged by the higher production of both IL-12 and IFN-γ. However, this combination stimulated the production of IL-10 by CD4+CD45+RA T-cells in co-culture with MoDCs. It is known that IL-10 is a potent anti-inflammatory and immunosuppressive cytokine. 31 Therefore, its up-regulation could be explained as a self-limitation mechanism of the Th1-mediated immunopathology in conditions of strong immune stimuli. 32
Another important finding of this study was the ability of MoDCs, pretreated with the highest concentration of 7-TOG, to stimulate IL-17 production by allogeneic CD4+ T-cells. This finding correlated with the increased production of IL-1β, IL-6 and TGFβ, but not IL-23, by MoDCs. Cytokine requirements for human Th17 differentiation are still emerging. Recent reports have shown an involvement of all these cytokines, but the precise role have not been elucidated yet. 33–35 Namely, it is known that the final differentiation of effector Th17 cells depends on IL-23, 36,37 but TGF-β and IL-6 are important for their induction. 38 Benwell and Lee 39 showed quite recently that in the presence of TGF-β, a high level of IL-β, independent of IL-6, is sufficient for optimal Th17 differentiation. Therefore, some of these cytokines and their combination might be responsible for the induction of the Th17 response. The production of IL-17 was additionally augmented when MoDCs were treated with both TLR agonists compared with the effect of the highest dose of 7-TOG alone, and this finding correlated with the increased production of IL-6.
A dominant finding of this study was that an intermediate concentration of 7-TOG (100 μmol/L) polarized MoDC functions toward the induction of the Th17 response, whereas Th1 and Th2 responses were not significantly modulated. This result was accompanied by up-regulation of CD86 and increased production of IL-1β and IL-6, but not TGF-β and IL-23 by MoDCs. The reason why the Th17 response is favored over Th1 and Th2 under such culture conditions, especially at the signaling level, is not clear. It is possible that stimulation of IL-1β and IL-6 was sufficient for differentiation of Th17 cells, as documented by a recent study. 40 The absence of significant Th1 induction could also favor the Th17 response, bearing in mind mutual antagonism between Th1 and Th17 immune responses. 41
So far, there is a lack of knowledge about the receptors on DCs which confer Th2-inductive capacity 42 and how pathogen-derived molecules bind to them. In this context, there are no data about the involvement of TLR7-induced signaling on the Th2 immune response. However, the role of TLR2 and TLR4 agonists were described in polarizing CD4+ T-cells toward the Th2 subset. 43–45 It has been implicated that ligation of TLR2 and MyD88 activation has some share in Th2 immune response initiation, 46,47 but Jankovic et al. 48 proved that DC-driven Th2 induction can also occur in an MyD88-independent fashion as well. In terms of cytokines, IL-4 and IL-10 are both interesting in driving the Th2 signal from DCs, but IL-10-deficient DCs can still initiate the Th2 response. 49 Moreover, the lowest concentration of 7-TOG slightly down-regulated the expression of CD40 on MoDCs by unknown mechanisms. This finding has some similarities with the results published by Karimi et al., 50 who showed that CD40 blockade in mice spleen DCs were associated with the increase in IL-4 secretion, shifting the DCs to stimulate Th2 cytokine production by allogeneic CD4+ T-cells, while the secretion of IL-12 by DCs was decreased. In addition, recent findings have shown that IL-4, through its activation of STAT6, upregulates GATA-binding protein 3 expression and also suppresses Th1 and Th17 cell responses, partly through the upregulation of growth factor-independent 1, a transcriptional repressor of IFN-γ and IL-17 production, 51 suggesting another mode of Th2 response prevalence over Th1 and Th17. The Th2 immune response was not significantly down-regulated when MoDCs were additionally treated with Poly (I:C), despite this combination being able to stimulate Th1 and Th17 responses. A number of studies showed that a single CD4+ T-cell may produce both Th1 and Th2 cytokines or Th1 and Th17 cytokines; 52,53 therefore, it would be interesting to study in future experiments if cytokine-producing cells in our experimental models were single- or double-positive.
An important goal in cancer immunotherapy is to identify an optimal protocol for preparation of immunogenic DC-based cancer vaccines. So far, the best results are achieved using potent activators of IL-12, such as Poly (I:C), a TLR3 agonist. 54,55 Higher concentrations of Poly (I:C) are toxic for the cells. 56,57 Therefore, the combination of Poly (I:C) with some other inducers of the Th1 response are promising. In light of recent findings showing that the Th17 immune response also participates in antitumor immunity, 58,59 our results suggest that 7-TOG, which is non-toxic even at very high concentrations 16 either alone, or in combination with lower concentrations of Poly (I:C), can be an alternative in the stimulation of MoDCs as cancer vaccines. However, this hypothesis deserves to be tested in future experiments using additional models such as the development of specific antitumor cytotoxic T-cells.
In conclusion, our results show that 7-TOG influences MoDCs in a dose-dependent manner and suggests that fine-tuned signaling through TLR7 may be modified by the engagement of TLR3, resulting in a different outcome of immune response.
Footnotes
ACKNOWLEDGEMENTS
The authors thank Sergej Tomic, Ivana Majstorovic, Sasa Vasilijic and Dragana Vucevic for helpful assistance during the experimental work. This study was supported by the project of Ministry of Science and Technological Development of the Republic of Serbia (project number 175102) and by a local grant of Military Medical Academy, Belgrade (VMA/06-10/A.3).