
Abstract
Weight regain after a long-term weight-loss program is a greater problem in obesity treatment than is weight reduction. Hence, the gut may elevate the absorption rate and nutrient transportation remarkably during chronic food restriction. The extension of gut absorption may be one possible reason for weight regain. But there is little information about the mechanisms that regulate intestinal absorption during food restriction. In this study, we show that the surface absorptive areas of gut villi may be enlarged in the jejunum of rats maintained on a food restriction regimen compared with animals submitted to swimming or sedentary behavior. Our findings show that simply reducing the amount of food intake results in an increased appetite accompanied with obvious weight regain, and suggest that the resulting enlargement of villi surface areas plays a key role in the regain of weight reduction. These results bolster accumulating evidence that gut absorption may be a substantial mechanism for resistance of weight loss and enhancing the weight regain process.
Introduction
Obesity is a worldwide health problem and is one of the highest risks for multiple diseases, such as malignant tumors, bone and joint injuries, insulin resistance and low reproductivity. 1,2 Therefore, weight loss is the concern of both patients and clinical providers. Based on the mechanisms of nutritional obesity development, exercise (increases of energy expenditure) and energy restriction (decreases of energy intake) are the major strategies to weight loss. 3 However, weight regain after a long-term weight-loss program is a greater problem confronted in the obesity treatment in clinical practice than is weight reduction. 4–7 It is estimated that more than 75% of obese individuals with weight loss would eventually confront weight regain, and typically return to their original weight in 3–5 years. 8,9 The exact mechanisms are yet to be identified.
The body is exquisitely designed to resist weight loss and is maintained in a normal range in a natural condition even in overweight or obese. Reducing body weight by decreasing energy intake has been shown to improve biomarkers for disease risks. 10 The propensity to gain and hold onto weight has been attributed to natural selection: powerful evolutionary forces, such as those that increase appetite and decrease the metabolic rate when fat stores are reduced. Many hormones (e.g. ghrelin, glucagon-like peptide 1, cholecystokinin, peptide YY3–36, leptin and insulin) are known to regulate appetite and metabolic responses to weight reduction. 11,12 However, the role of the gut absorption in weight regain has not been well established. Hence, the gut may elevate the absorption rate and nutrient transportation remarkably during chronic food restriction 13,14 and the elevation of gut absorption may be one possible reason for weight regain. But there is little information about the mechanisms that regulate intestinal absorption during food restriction.
In our recent study, we noticed that the gut villi may enlarge their surface absorptive areas by forming depressions during food-restriction weight-loss programs. This result provides morphological evidence for the gut mechanism of weight regain. This report is the result of our findings.
Methods and materials
Diet-induced obese rat model
Male Sprague-Dawley (SD) rats (Charles River, Beijing, China) were fed a high-fat diet (HFD), 5.58 kcal/g with 67% energy intake from fat (16% protein, 42% carbohydrate and 31% fat; KeAo Corp., Beijing, China) (HFD rats, n = 100) or a chow diet, 3.16 kcal/g with 11.4% energy intake from fat (20% protein, 50% carbohydrate and 4% fat, KeAo Corp.) (chow rats, n = 40). All animals were free to access food and drink ad libitum. Rats were individually housed in a specific pathogen-free facility with 12:12-hour dark–light cycles. The mean room temperature was 22 ± 2°C and the mean humidity level was 65 ± 5%. Animal protocols were approved by the Animal Welfare Committee of Peking University. After the 15-week HFD challenge, the top quartile of HFD rats were chosen as the obese and signed onto weight-loss interventions.
Weight-loss interventions
We designed three types of weight-loss paradigms for five weeks: 15 diet-induced weight loss (food restriction, FR), exercise-induced weight loss (swimming, SW) and diet plus exercise-induced weight loss (swimming plus food restriction, SF). 3 Obese rats were divided into four groups matched for weight. A sedentary, free-eating group was provided with food ad libitum (diet-induced obesity [DIO], n = 7). An exercising group, fed ad libitum, was subjected to a program of swimming (SW, n = 5). A sedentary, paired-weight group was food-restricted (FR, n = 7). An exercising group was subjected to a program of swimming and was diet-restricted (SF, n = 6). The intensity of each treatment was as follows: in the FR paradigm, the daily food intake was reduced to 45% of the usual intake; 16 in the SW paradigm, rats swam 40 min a day six days per week; 3 in the SF paradigm, the daily food intake was reduced one-third of the usual intake together with swimming 40 min a day six days per week. Due to the different mechanisms of lose weight in the obese, i.e. energy-intake restriction or energy-expenditure elevation, we took the visceral fat (epididymal and retroperitoneal fats) as the equivalent to evaluate whether or not they were iso-carloric. All rats were fed the same HFD during the treatment without meal replacement.
Dietary programs for weight loss
There were two calorie restriction (CR) strategies: the major form is to reduce energy composition of daily diet, for example, replacement of HFD for low-calorie diets (LCDs) or very low-calorie diets (VLCDs) daily or intermittently; 3,8,17,18 and the other is to cut the daily food intake without diet change. Considering that diet-switching may cast influences on villi morphology, 19,20 and food restriction had been almost routine among overweight human populations in developed countries, 14 we chose the latter strategy and added necessary vitamins and minerals to the water. 15 To avoid weight regain drastically, food was restricted progressively, i.e. reduced one-third of the usual food intake. When the body weight began to regain and return to their initial levels, another one-third was cut-off until the end of the experiment. 21
Physical activity
In the SW paradigm, animals were transported to a treatment room each day, where exercise animals were allocated to swim. Animals swam in a swimming pool with a diameter and height of 5 ft and 15 inches, respectively. Water was maintained at 30.0 ± 2.0°C. To minimize the influence on night-eating, swimming was performed at 8:00, as described previously. 22,23 At the end of the treatment period, rats were dried in a towel and left for about one hour in a heated room to minimize the effects of cold exposure. The SW rats were provided with food ad libitum.
In the SF paradigm, rats swam and were submitted to food restriction as for the SW and FR rats.
During weight-loss interventions, rats' body weight and appetite were monitored. Appetite was measured as the amount of food intake. In this experiment, we recorded the 24-h food-intake curve from 8:00 and took daytime eating as the stigmata of appetite increasing.
Histological assay
After a five-week weight-loss program, approximately 24 h after the end of the last exercise bout, rats were anesthetized with an intraperitoneal injection of pentobarbital sodium (5 mg per 100 g of body weight). To get rid of the possible influence of blood on the villi morphology, such as congestion or edema, animals were sacrificed by abdominal aorta blood draining with a 10 mL syringe. Then the jejunum was removed immediately for histological analysis. Tissue samples, 0.4 inches long, were excised four inches distant from the pylori ring. The tissue was split along the mesentery border, and was rinsed three times in 0.9% saline to clean food debris. Then the tissue was fixed in 10% buffered paraformaldehyde in phosphate buffer saline for 24 h and embedded in paraffin. Four micrometer thick sections were prepared (Leica RM2016; Leica Microsystems, Shanghai, China) for hematoxylin–eosin staining. Then the structures were observed using an Olympus BX-60 microscope (Olympus, Tokyo, Japan). For semi-quantitative analysis, the height and diameter of villi, thickness of epithelia layer, circumference and goblet cells per 100 cells were measured. The villi height was the distance from the apex of the villus to the base of the crypt in perpendicular oriented histological sections.
Intestinal tissues were analyzed using scanning electron microscopy (SEM) as previously described. 24,25 In brief, jejunal mucosa was fixed in 4% glutaraldehyde at 4°C for four hours and then postfixed in osmium tetroxide for one hour. Then the tissue was examined using a JSM-6700F SEM (JEOL, Tokyo, Japan) at 10 kV or embedded in Epon after graded ethanol (30%, 50%, 70%, 85% and 95%) dehydration. Ultrathin sections were stained with uranyl acetate and lead citrate. Ultrastructural aspects of villi were examined using a 80 kV accelerator voltage JEM-1400 transmission electron microscope (TEM) (JEOL) equipped with an Olympus digital camera.
Statistic analysis
Statistic analysis was performed using SPSS software version 13.0 (SPSS Inc., Chicago, IL, USA). All variables were checked regarding their normal distribution using the Shapiro-Wilk test. All data are shown as means ± SD. A P value <0.05 was considered statistically significance. One-way analysis of variance (ANOVA) was used for single-point measures of terminal body, adipose depot weights and villi sizes at the end of each phase. ANOVA for repeated measures was used to analyze body weight, Lee Obese Index and appetite. When significant intergroup differences were found (P < 0.05), post hoc Scheffe's multiple comparison tests were carried out to identify the specific location of the significant effects. Weight change (gain, loss or regain) during intervention was assessed by independent sample t-test. The effect of intervention on the body weight (before and after treatment) of each subject was assessed using paired sample t-tests.
Results
Diet-induced obesity
Induction of dietary obesity
Values are expressed as means ± SD
*Obese was the upper quartile of high-fat diet (HFD) rats, selected at the end of the 15th week and submitted to further weight-loss program
‡Chow versus obese and HFD groups, P < 0.0001
Weight-loss program
During weight loss, there were no abnormal physiological impacts of gut activity observed, except for a significant reduction of feces in FR and SF rats (data not shown). After five weeks of the weight-loss program, FR and SF rats did not show any signs of malnutrition.
Weight-loss effect of different treatments
DIO, diet-induced obesity; FR, food restriction; SF, swimming plus food restriction; SW, swimming; BW, body weight
Values are expressed as means ± SD. Means sharing the same symbol denote significant differences between before and after treatment: *P < 0.05; **P < 0.0001 Values with dissimilar superscript letters differ from each other at P < 0.05 or less by post hoc test after analysis of variance showed there to be significant intergroup differences ‡Visceral fat: epididymal and retroperitoneal fats
Villi morphology
The morphology of gut villi dramatically changed during obesity or weight loss. The villi of obese rats distended with a broadened subepithelial space, and the uppermost tip of villi was opened (Figures 1a and b). This morphological change was a result of the extra accumulation of lipid droplets in cytoplasm. The villi of the jejunum in FR and SF rats appeared corrugate as evidenced by the histological appearance of a petal-shaped configuration with lots of depression formations, and by the SEM appearance of a rather rough surface of villi (Figures 1c–g). In addition, all depressions were coated with microvilli (Figures 1e [arrowhead] and g). This configuration was not a consequence of villi shrinkage, but epithelial deformation, as indicated by a lack of basement membrane distortion (Figure 1e, arrow). While the villi morphology of SW rats was similar with those of the chow rats, there were no corrugations in epithelial cells (Figures 1h–j). Also, the accumulation of lipid droplets in cytoplasm was subtle despite the same HFD fed as DIO rats.
Jejunum villi dynamically changed in morphology in obese and weight-loss rats. Villi of the obese rats distended with the subepithelial broadened space (a) and the uppermost tips opened as seen by the scanning electron microscope (SEM) (b). Food restriction caused the villi to shrink with the formation of many depressions in a petal-shaped configuration ([c] food restriction; [d] food restriction plus swimming), which showed a rough surface by SEM observation (f). The depression surface was coated with a brush-border (arrowhead, [e]) which was confirmed by transmission electron microscopy (g), and the basement membrane was not distorted (arrow, [e]). Panel e shows the fivefold magnifications of the corresponding boxed area in (c). SW rats' villi form and surface resembled that of the chow rats' smooth surface by light microscopy (h, i) and SEM (j). HE staining, magnification was ×400 (a, c, d, h and i). (A color version of this figure is available in the online journal)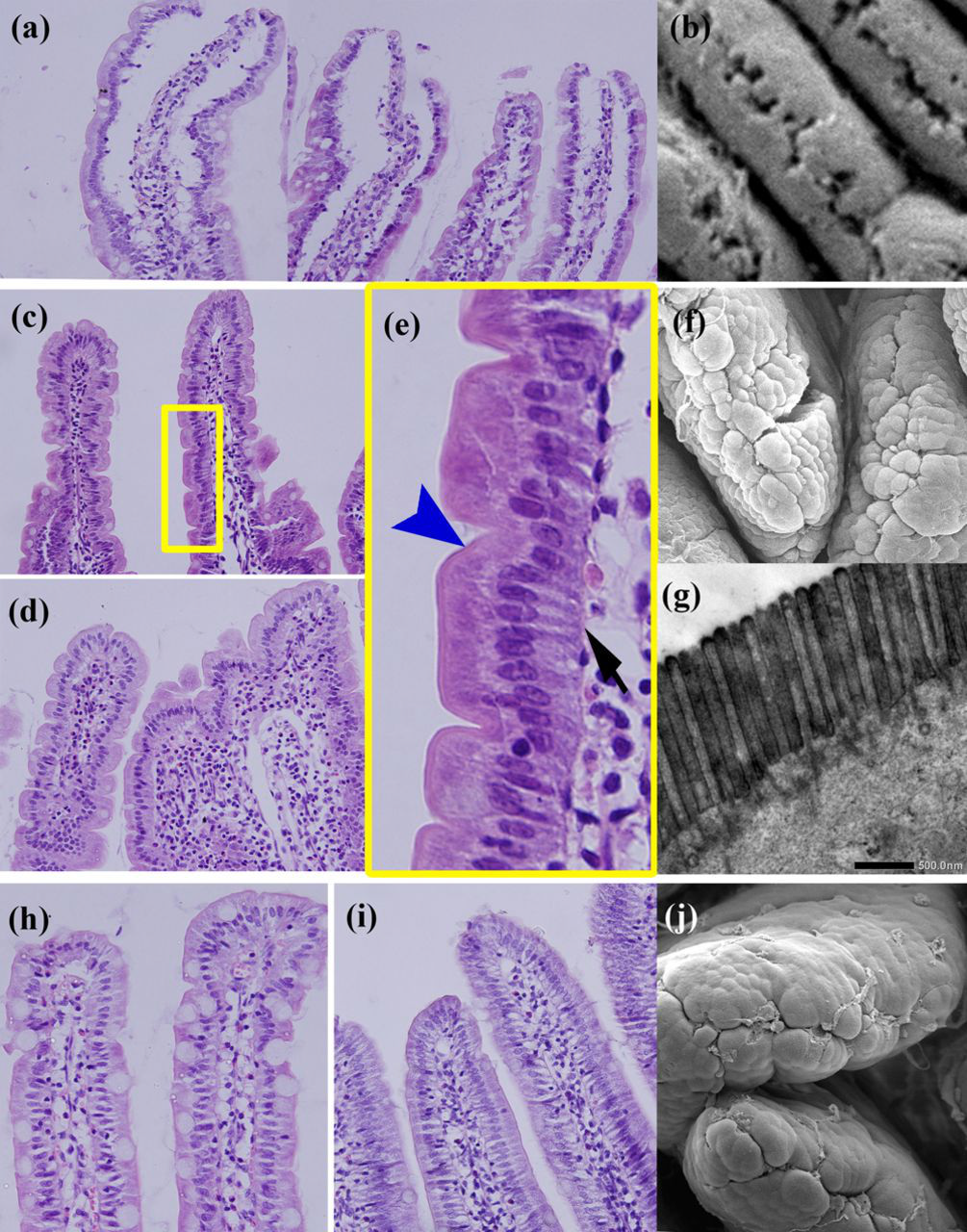
Villi measurement
Measurement of jejunal villi in obese and weight-loss rats
DIO, diet-induced obesity; FR, food restriction; SF, swimming plus food restriction; SW, swimming
Values are expressed as means ± SD. Values with dissimilar superscript letters differ from each other at P < 0.05 or less by post hoc test after analysis of variance showed there to be significant intergroup differences
*Magnification was ×200
‡Magnification was ×400
Appetite compensation and weight regain
Rats in the FR group ate nearly 80% of their designed daily diet within one hour during lights-on time (10:00–11:00), and rats in the SF group spent seven hours (10:00–17:00) of the lights-on time to eat 70% of their designed diet. Appetite in SW, chow and DIO rats was comparable. Most of them ate 80% of their daily food in 15 h during lights-off time (17:00–8:00), P > 0.05 (Figure 2).
Appetite changes in obese rats during a five-week weight-loss intervention. Values represent means ± SD for five (SW), six (SF) and seven (chow, DIO, FR) subjects. Repeated measures analysis of variance showed a significant increase of appetite for FR (P < 0.0001) and SF (P < 0.0001), but no increase for SW (P > 0.05). SW, swimming; SF, swimming plus food restriction; DIO, diet-induced obesity; FR, food restriction. (A color version of this figure is available in the online journal)
FR rats lost 4.48 ± 5.67 g of body weight per week in the first two weeks of food restriction, but returned to their initial body weight levels by 10.57 ± 6.68 g body weight regain in the third week. When another one-third of the food was restricted, their body weight reduced significantly followed by weight regain in the following week (Figure 3b). SF rats lost 16.83 ± 6.66 g of body weight per week during the first two weeks, but only lost 3.86 ± 3.17 g per week in the following weeks (Figure 3c). Swimming alone only lost 2.66 ± 5.71 g body weight of rats per week, and regained 13.05 ± 9.30 g during the fourth week (Figure 3d).
Body weight maintenance or loss resistance during a five-week weight-loss intervention. Values represent means ± SD for seven (chow and DIO [a], FR [b]), six (SF [c]) and five (SW [d]) subjects. Independent sample t-test showed a significant weight regain of FR rats (P < 0.0001) in the third week and weight maintenance of SW rats (P > 0.05). The occurrence of weight loss of the FR rats during the fourth week was the result of the additional one-third food restriction. SW, swimming; SF, swimming plus food restriction; DIO, diet-induced obesity; FR, food restriction
Discussion
It is estimated in research and clinical practice that more than 75% of weight loss in obese individuals will eventually be regained. 8 However, the underlying mechanisms are still unclear; the change of the gut might be one target for understanding the retroversion weight regain in weight-loss obese patients. To date, there is no evidence showing that villi changes are involved in the pathophysiology of this disease.
In this study, we showed that villi corrugated with microvilli-coated depressions formed in the intestines of dietary weight-loss rats. These changes significantly increased the size of villous surface areas and are associated with the increase in appetite and weight regain. To the best of our knowledge, this is the first villi morphology observation in obese and weight-loss rats. The major significance of the morphological change is that the microvilli-coated depressions may increase the size of the villous surface areas significantly. This may be the structural foundation of the adaption enhancement of intestinal nutrient transport rates in chronic caloric restriction 13,28 and provides morphological evidence for the gut mechanism in weight regain.
Previous studies have demonstrated that wild animals and starved laboratory animals 29,30 had atrophied villi and shortened and sparse microvilli with progressive decreases in the sizes of villus, crypt and mitotic pool according to their energy supply. Examples include the python and broad-nosed caiman 31 kept on irregular meal times, the green-striped burrowing frogs and 13-lined ground squirrels in seasonal dormant, and the red knot and sandpiper having varied kinds of food supply from shrimp to mussels; the gut villi of all of these animals atrophied at lean time and thrived quickly during eating time. 32 These morphological changes show the ‘functional adaptation’ of the gut to the food supply. In this morphological observation, it could be predicted that the gut can ‘functionally compensate’ for energy supply deficiency. We noticed that the morphological changes in FR rats were in line with their appetite increase. During the treatment, SW rats finished 80% of their daily food amount in 15 h mainly during the night (17:00–8:00), which was comparable to those of their chow and DIO partners. Meanwhile, food restriction resulted in significant daytime eating which implied an increase in their appetite. FR rats finished nearly 80% of their daily diet in one hour during the daytime (10:00–11:00), and SF rats finished 70% of their daily diet in seven hours during the daytime (10:00–17:00) (Figure 2). Since the appetite regulatory hormones, such as leptin and ghrelin, quickly defend impositions that promote a negative energy balance, and increase the appetite in weight loss resulting in metabolic and behavioral compensatory responses, 11,12,31–33 it can be speculated that the villi change in weight loss is another counter-regulatory way to a weight-reduced state, and responsible for weight regain.
The gut is a dynamic organ, subject to changes in functional molecules and morphology in response to many variables, including diet, the environment, drugs and health status. 31–37 It can only be deduced that the gut mechanism may be associated and contributes to the dietary weight regain. But how these morphological changes occur is still under speculation. Is it a reaction from weight loss, fasting or starvation? Since the processes of digestion, secretion of digestive enzymes and then absorption are widely regulated by the neuroendocrine system, 38 the gut–neural interaction may be one possible explanation. In fact, villi are innervated directly by many kinds of neurons derived from the sympathetic, parasympathetic (vagal nerves) and intrinsic system of the gut. Dietary weight loss is directly correlated to elevated levels of appetite – causing the activation of the autonomic nervous system. Therefore, the activation of the nervous system in dietary weight loss is likely accompanied by the change in villi morphology. 39 Further prospective studies are also needed to evaluate the underlying mechanisms in villi morphological changes during dietary weight loss.
Fasting and exercise are the most popular strategies for weight loss. However, the readouts are always distinct. This finding provides new considerations for weight maintenance program designs during or after weight loss. Dietary-restriction induced weight loss is more likely to have weight regain, partly due to hormone-regulated energy expenditure reduction and appetite increase. For instance, the plasma ghrelin concentration is strongly correlated with body weight, 40 and negatively correlated with body mass index in humans. The plasma ghrelin concentration returns to normal after diet-induced weight loss. 15,41,42 However, increase in appetite alone is not enough to explain weight regain especially under energy-restricted conditions. The gut mechanism may be one explanation. During treatment, FR rats lost weight in the first two weeks, but regained it in the third week and returned to their initial weight level. Considering the elevated villous surface areas, the third-week regain may have been the result of, at least in part, the gut absorption enhancement. Swimming is another popular obesity therapy strategy. In this study, it was observed that swimming also resulted in weight loss and regain. But the gut villi seemed to contribute less to the regain. Because lipid droplet accumulation was not seen even exposed to the same HFD as non-treated obese rats, their villi showed elevated energy metabolism rather than elevation of absorption. As the visceral fat was the same as FR and SF rats, this regain may be the result of the elevation of muscle hypertrophy. SF rats showed potential hopes for conquering the weight regain from villi absorption. Although they had the same villi absorption area enlargement as FR rats, and also experienced weight regain, their body weight never returned to the initial levels. It can be assumed that the compensated elevation of gut energy absorption was consumed by exercise. So, moderate exercise is a valuable strategy for food restriction to lessen the increase of appetite and absorption. This is consistent with previous reports that increased physical activity may help weight maintenance after dietary weight reduction. 3,43–45
In summary, this article provides morphological evidence for the enhanced absorption of the gut during fasting. By food restriction, we found that the gut may possess a compensatory absorption mechanism for the body rather than a passive adaptation which may contribute to the weight regain in obesity therapy. Further investigation is needed to explore the functional and molecular mechanisms that are connected with the morphological changes.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (30270636 and 30671015) and the Fundamental Research Funds for the Central Universities (lzujbky – 2010-125). We thank YL Yang and L Chen for their technical assistance, and XN Wang and SS Tsien for help in the modeling process.