
Abstract
In the present work we studied synaptic protein concentrations in relation to behavioral performance. Long-Evans rats, aged 22-23 months, were classified for individual expression of place memory in the Morris water maze, in reference to young adults. Two main subgroups of aged rats were established: the Aged cognitively Unimpaired (AU) had search accuracy within the range (percent of time in training sector within mean+2 SEM) of young rats and the Aged cognitively Impaired (AI) rats had search accuracy below this range. Samples from the hippocampus and frontal cortex of all the AI, AU and young rats were analyzed for the expression of postsynaptic protein PSD-95 by Image J analysis of immunohistochemical data and by Western blots. PSD-95 expression was unchanged in the hippocampus, but, together with synaptophysin, was significantly increased in the frontal cortex of the AI rats. A significant correlation between individual accuracy (time spent in the training zone) and PSD-95 expression was observed in the aged group. No significant effect of age or PSD-95 expression was observed in the learning of a new position. All together, these data suggest that increased expression of PSD-95 in the frontal cortex of aged rats co-occurs with cognitive impairment that might be linked to functional alterations extending over frontal networks.
Keywords
Introduction
Aging is associated with a cognitive decline in various species including rodents and humans. Several studies have shown that aging is accompanied by memory deficits in humans.1,2 Aged rats also show memory impairments, mainly assessed by spatial memory tests like the Morris water maze. These spatial memory deficits are associated with neurobiological changes such as decrease in cholinergic transmission, increasedoxidative stress, decreased functional synaptic plasticity and neurogenesesis.3–6
Interestingly, in both rats and humans, a high heterogeneity is commonly observed, with some individuals showing strong cognitive impairments while others present preserved memory performance. Understanding the brain modifications related to cognitive impairments or to preserved efficacy is a major challenge. In humans, ‘successful aging’ has been linked to compensatory activation in frontal lobes during cognitive testing2,7 and synaptic alterations rather than massive cell death have been observed with pathological aging. 8 Increase in postsynaptic proteins and compensatory dendritic branching have also been reported in Alzheimer disease, mainly at earlier stages. 9
In aged rats, the water maze is one of the tests most frequently used to assess spatial memory performance. A common experimental rationale is based on a cognitive classification of the performance of older subjects, as cognitively impaired or unimpaired.10–12 This allows relating the cognitive decline in some individuals with various neurobiological changes, like gene expression in the hippo-campus, 12 dysregulation of synaptic plasticity-associated proteins 13 or pre- and postsynaptic proteins, including PSD-95 (postsynaptic density protein-95) protein variations in different neocortical regions of aged rodents.11,14,15
Synaptic proteins are important molecular markers of brain aging. PSD-95 is a major scaffolding protein molecule at glutamatergic synapses, known for its role in regulation of synaptic plasticity and signalling pathways. 16 Excess or deficiency of this protein interferes with its physiological functions. 17 As enhanced PSD-95 expression may be a marker in Alzheimer disease,18,19 we were interested to study the link between PSD-95 expression and cognitive impairment in aged rats.
The aim of the present work was thus to analyze age-related changes of pre- and postsynaptic proteins – including PSD-95 protein – in the prefrontal cortex and the hippocampus of two-year-old rats and to correlate them to behavioral performance. We used the Morris place learning task with a fixed escape position design over five days, then with a new daily escape position for a working memory task design. Rats in the aged group were classified as Aged Impaired (AI) or Aged Unimpaired (AU) relative to young rats using spatial accuracy during a probe trial with no escape platform. Immunohistochemistry for overall PSD-95 expression was carried out in the hippocampus and the frontal cortex of all the rats and correlations between behavior and PSD-95 expression were analyzed. Differences in immunohisto-chemical PSD-95 expression were corroborated by Western blots in a representative subgroup of AI and AU cases. Neuritic and presynaptic changes (e.g. Tau and synapto-physin) were assessed at the same time to further characterize neuronal alterations in non-pathological brain aging.
Materials and methods
Animals
A total of 26 male Long Evans rats were used in this study. The age groups were as follows: 7 young adults, aged 5 months and 19 old rats, aged 22-23 months. Rats were born in our animal facilities from eight pairs of two-month-old rats purchased from Elevage R. Janvier (Le Genest, St Isles, France).
These rats were used in compliance with the Swiss Guidelines for the Care and Use of Laboratory animals. They were housed in groups of two to three in plastic Macrolon cages type IV (595 × 380 × 200 mm; Indulab AG, Gams, Switzerland) with a wooden nest box. They had free access to food and water and were maintained under a controlled light-dark regimen 12/12 h (lights on at 7:00 hours).
Water maze
Apparatus
The set-up was located in a room with numerous uncontrolled distant cues (e.g. door, diffuse lights). The large gray circular water tank (160 cm in diameter, wall height of 60 cm) was filled to a height of 40 cm with water (25°C) made opaque with 0.5 L of milk. Four orthogonal starting positions around the pool perimeter divided its surface into four quadrants. An invisible escape platform (diameter = 14 cm) could be placed in the center of each quadrant, submerged 2 cm below the water surface. A videocamera was placed directly above the center of the pool. The animal's activity was recorded for later analyses.
For training trials, the tested animal was released in the water from one of the four starting positions, facing the pool wall. It was given 60 s to locate and climb onto the platform, where it stayed 30 s before the next trial. If a rat did not find the platform in 60 s, the experimenter guided the animal gently to it. The start sequence was pseudorandomly selected and was different for each block, and the escape latencies were measured in seconds with a stopwatch.
During the 60-s probe trial with no escape platform, the rat was released from a starting position located opposite the trained escape location. The time spent in four circular zones (diameter: 40 cm) each centred around possible platform locations was analyzed by a video tracking system (Ethovision Noldus IT, Wageningen, Netherlands). These zones were named from their position relative to the training position (i.e. t for training location, t − 1, t +1 and opp).
Procedure
During the acquisition phase (five daily sessions of four training trials), the submersed platform remained at a fixed position. To assess immediate retention, a probe trial was introduced 30 s after the last trial of session V.
One week after the acquisition phase, training was continued during two days (sessions VI and VII) while the platform position was changed daily (new position learning). Two different new positions were chosen (adjacent and opposite to the original escape position, with equal distance to the wall). Four different start positions were pseudoran-domly selected, as during the acquisition phase. Rats were allowed 30 s on the platform. A short delay (1 min) was introduced between trials 1 and 2 in a bucket containing absorbent paper, after which it was released in the water for the second trial from another start position. Two subsequent trials with the usual short intertrial interval (30 s in the bucket) and same platform position were conducted.
Cognitive assessment
During the probe trial, the total time spent by each subject in four circular symmetrical zones including the target area is taken as 100%. A chance distribution of the swim paths leads to around 25% of time spent in each zone. Individual scores of the percent time spent in the trained zone during the probe trial were used to propose a classification of cognitively impaired old rats based on individual spatial accuracy. Rats in the aged group were regarded as impaired if the time spent in the training zone was shorter than the mean value minus 2 standard error means observed in the young group. The remainder of the rats were considered as cognitively Unimpaired (AU), as their score fell within the range of those of the group of young rats providing a standard value. There is a high diversity in the behavioral indicators of cognitive impairment in the water maze. Accuracy during probe trials has been used by other studies to classify the performance of old rats.12,13,20 It estimates spatial accuracy and we think that this measure is more appropriate than latencies for categorization.11,14,15 The fact that our method reveals two of the seven tested young subjects not satisfying the criterion for accuracy is a problem common to this sort of classification and has already been observed. 21
Brains
All rats were euthanized three weeks after water maze training. Rats were first anesthetized with Nembutal (pentobar-bital 50 mg/mL) and then decapitated. One hemisphere of the removed brains was immediately frozen with dry ice and stored at -80°C for biochemical Western blot analyses. The second hemisphere was fixed in 4% formaldehyde for three days, immersed in phosphate-buffered saline (PBS) at 4°C, then dehydrated during two days in successive alcohol concentration baths (70%, 80%, 95% and 100%), incubated three days in Cedar essential oil and subsequently embedded in paraffin for immunohistochemical analysis and densitometry measurement. Serial frontal sections were cut at a thickness of 20 μm with a microtome (Reichert-Jung, Seefeld, Germany) and mounted on Super-frost Plus microscope slides. Room temperature dried slides were then put for one hour at 60°C.
Histology and immunohistochemistry
Immunohistochemical analyses were made following brain extraction in all rats, i.e. seven young and 19 old subjects. The whole hemisphere was cut in a coronal plane and immunohistochemical staining was performed on 20-μm thick serial sections after they were deparaffined and subsequently treated 10 min in methanol, rinsed five minutes in filtered H2O (Millipore, Zug, Switzerland), microwaved five minutes in citric acid and finally washed 10 min in PBS. After a 30-min incubation in normal serum (swine 1:10; Dako X0901, Baar, Switzerland), sections were incubated overnight at 4°C with the primary anti-PSD-95 antibody (Abcam 18258, Lucerna Chem AG, Lucerne, Switzerland; dilution 1:150). After 2 × rinsing in PBS for 10 min, sections were incubated for two hours at room temperature with the secondary antibody (Swine anti-rabbit; Dako E0353; dilution 1:300). Sections were then rinsed again 2 × for 10 min in PBS and incubated in Avidin Biotin Complex/horseradish peroxidase (HRP; Dako K0355) for one hour, following recommendations of the manufacturer. After rinsing 2 × in PBS for 10 min, the immunoreaction was revealed by a 10-min incubation in 0.03% 3,3’-diaminobenzidine (Sigma D5637, Sigma-Aldrich Chem GmbH, Buchs, Switzerland) and 0.015% H2O2/PBS. Absence of labelling resulted from immunostaining performed with omission of the primary or secondary antibody. All sections were mounted with pertex mounting medium (Histolab 00811, Gothenburg, Sweden). Please see Table 1 for details on antibodies.
Description of antibodies used for Western blots
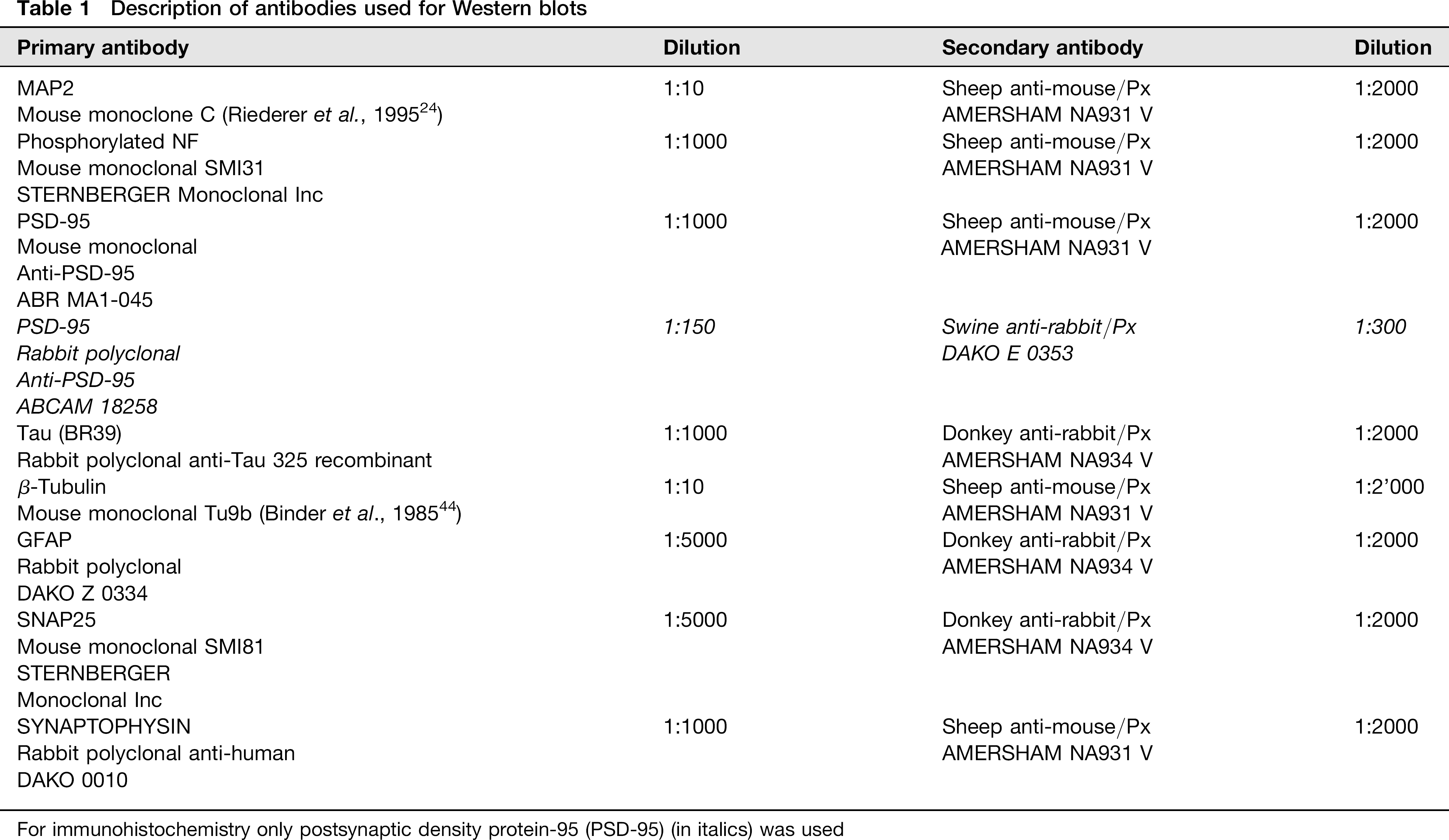
For immunohistochemistry only postsynaptic density protein-95 (PSD-95) (in italics) was used
Image J analysis of immunohistochemical data
Image J densitometry was carried out for the immunostaining of the PSD-95 marker, with a standardized method successfully used in human cortex. 19 We analyzed the intensity of optical density of PSD-95 both in the hippocampus and in the frontal cortex. The rodent frontal cortex can be divided into three parts, the medial, the lateral and the ventral parts. 22 According to the latter authors, we chose to analyze the medio-dorsal part of the prefrontal cortex, as it has been shown to be more implicated in memory for motor response than the ventral part. The medio-dorsal part includes the precentral cortex, the prelimbinc cortex, the anterior cingu-late and the medial orbital cortex, but we chose to analyze this region as a whole as the cytoarchitectonic limits between subregions were not clear on the PSD-95 staining. Coronal serial sections were chosen at about 2 mm of the rostral pole that correspond to the plate at Bregma 0.2 mm of the Paxinos atlas. 23 The hippocampus was analyzed first as a whole, taking together the dentate gyrus and the CA1-CA4 subregions, between the levels A 2.000 and A 2.500 of the same atlas. Then, samples of the dentate gyrus and of CA1 and CA3 were analyzed separately with the same method.
The Image J analysis system (NIH, Bethesda, MD, USA, public domain) on a Compacq computer was used for the densitometry. Color digital images were taken from the immunostained sections at a magnification allowing the visualization of the whole cortical plate or hippocampus, and at the same luminosity of the microscope for all captures, as a result of a controlled voltage (5.7 V). Images were then transformed into gray level values and densito-metric measurements were performed on each image. The intensity of optical density corresponding to the intensity of the transmitted light was measured on a scale going for each pixel from 0 (100% transmitted light) to 255 (0 transmitted light). Background was measured on individual photomicrographs outside of the tissue and automatically subtracted from the densitometric measurements. Two sections were analyzed blind per rat in the defined hippocam-pal and frontal regions and a mean densitometric value was calculated in each case.
Electrophoresis and Western blots
Electrophoresis and Western blots were performed as previously described. 24 Analyses were done in eight aged rat brains (four AU and four AI) from animals tested in the water maze, randomly selected among the ones with the best and worse performances, in order to confirm immuno-histochemical data obtained for PSD-95. In addition, other proteins were analyzed as well (Table 1). Frozen samples of frontal cortex and hippocampus were homogenized and prepared for sodium dodecyl sulfate polyacrylamide gel electrophoresis. Samples were separated on a 3.6–15% acrylamide gradient. Western blots were prepared and immunostained with antibodies described in Table 1. The immunoreaction was revealedby chemiluminescence according to the manufacturer's guidelines (Amersham Pharmacia Biotech, Dübendorf, Switzerland). Blots were wrapped in Saran Wrap and quickly exposed to an autoradiography film for several seconds or minutes (Kodak Biomax MR, Lausanne, Switzerland). Western blots with the same samples from the frontal cortex were repeated three times.
Quantitative differences in Western blots were analyzed by densitometry analysis with a computer software 1D Main (American Applied Biotechnology, Fullerton, CA, USA), expressed in relative density units, and corrected for loading differences by dividing with the tubulin values. Immunoreactivity quantified by densitometry was used for correlation with behavioral studies.
For the production of antibody Tau BR39, a plasmid pET-htau352wt was obtained from Prof Dr Roland Brandt (Abteilung Neurobiologie, University of Heidelberg, Germany) and used to produce recombinant tau according to a published protocol. 25 Antibodies against recombinant tau352 were produced in adult New Zealand White rabbits by injecting 30 μg of proteins and Specol as an adjuvant. After a month, fortnightly injections of antigen and Specol were given, until a satisfactory antibody titer was reached. The specificity of the serum samples of rabbit No. tau39 was tested on human brain homogenates and revealed a good reactivity between 1:1000 up to 1:10,000 dilution.
Statistical analysis
Statistical analyses were conducted using SPSS software (IBM SPSS Statistics, Hong Kong, China; version 18.0).
Results
Behavioral performance
Acquisition and retention
First, we compared the performances of young and old rats to assess age-related differences and then we compared subgroups of aged rats (AI and AU rats) with young rats. During acquisition, there was a general reduction over sessions in escape latencies. A two-way analysis of variance (ANOVA) (age and sessions II-V as repeated measure) revealed a non-significant decrease between sessions: (F(3,72) = 2.35; P = 0.079), a non-significant tendency of the old rats to have longer latencies (F(1,24) = 2.164; P = 0.154) and no sessions × age interaction (F(3,72) = 0.956; P = 0.418).
Immediate retention was tested in a probe trial conducted 30 s after the last trial of session V. Both groups of young and old rats spent more time in the training zone. This was confirmed by a two-way ANOVA (age and time in zone as repeated measure) showing a significant zone effect (F(3,72) = 17.23; P < 0.001) but no age × zones interaction (F(3,72) = 1.46; P = 0.23). Comparing the absolute time spent revealed no significant difference between old and young rats (unpaired Student's t-test t (24) = 1.524; P= 0.923).
Overall, there was no correlation between mean individual (old and young rats) latencies (sessions II-V) and percent time spent in the training zone (r = −0.26; P = 0.18, two-tailed), indicating that longer latencies were not correlated with less time spent in the training zone during the probe trial. Separate analyses confirmed that there was no correlation between mean individual latencies (sessions II-V) and percent time spent in the training zone neither in old rats (r = −0.285; P = 0.23, two-tailed) nor in young rats only (r = −0.393; P = 0.38).
As described in the Materials and method section, the performance during the probe trial (percent time in the target) was used to separate cognitively preserved old rats from impaired ones. Seven rats in the aged group were thus classified as Aged Impaired (AI, n = 7), i.e. rats with a score under 35.70% (mean — 2 SEM of the young rats’ performance, i.e. 48.10% - 12.40). The remainder of the rats were considered as unimpaired (AU, n = 12).
Separate analyses of escape latencies in the AI (n = 7) or AU (n = 12) as in Figure 1a (left) revealed that AI rats had prolonged latencies in comparison with AU and young rats. This impairment was confirmed by the significant group effect (F(2,23) = 4.512; P = 0.022) provided by a two-way ANOVA (groups and sessions II-V as repeated measure). The session effect was significant (F(3, 69) = 3.216; P = 0.028) but there was no group × sessions interaction. Post hoc analyses (Bonferroni) confirmed that the AI group had significantly longer latencies than young (P ≤ 0.001) and nearly significant than AU (P = 0.057) groups, whereas AU rats’ latencies were not significantly different from those of young rats (P = 1).
(a) Left: escape latencies (mean ± SEM) during the five sessions of task acquisition. Right: escape latencies during new platform position learning trials combined over two sessions (VI–VII) in young rats (n = 7), Aged Impaired rats (AI; n = 7) and Aged Unimpaired rats (AU; n = 12). (b) Spatial discrimination during the probe trial occurring immediately following the last acquisition trial of session V (time spent in each zones, mean ± SEM, t = training zone) in young (n = 7) and old rats (AI (n = 7) and AU (n = 12). Significance of statistical comparisons between mean time in the training zone and other zones is indicated by **(P < 0.01). Representative paths are presented in the left of the figure illustrating swim styles in the two groups. The exact percent spent by each subject in the training sector is indicated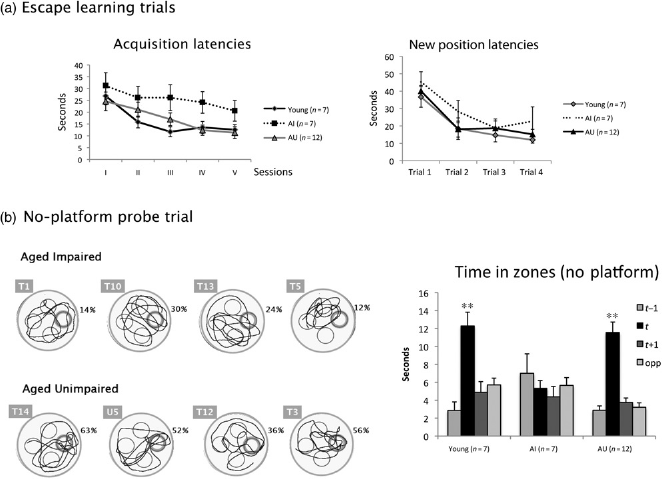
The poor performance of the AI during the probe trial (Figure 1b) is obviously an expected consequence of our classification. However, there was no difference in swim velocity neither between AI and AU (P = 0.54) nor between AI and young rats (P = 0.13), showing that the difference was not due to motor deficit and consisted in spatial accuracy and focusing on target impairments (see also representative swim path in Figure 1).
New position learning
A two-way ANOVA of the mean escape latency of trials 1-4 of the two new position learning sessions in the three groups (young, AI and AU) showed a significant trial effect (F(3,69) = 28.89; P < 0.001), no group effect (F(2,23) = 1.07; P = 0.357) and no group × trial interaction. A separate ANOVA (young versus AI, trials 1-4 as repeated measure) showed no significant group effect (F(1,12) = 1.635; P = 0.225) despite a tendency of the AI to have longer latencies (Figure 1a, right).
Image J analysis of PSD-95 expression in AI and AU groups
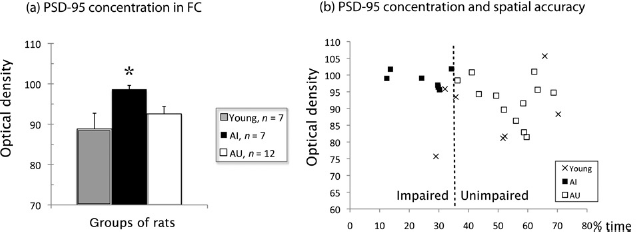
Image J densitometry of PSD-95 protein was performed in the hippocampus and frontal cortex of young, AI and AU rats. No disparity in the level of PSD-95 was observed across groups in the hippocampus of AI compared with AU rats (Figure 2b) and analysis by Image J showed no significant difference between groups (P > 0.05). In addition, there were no significant differences in hippocam-pal subregions such as the dentate gyrus or CA1 and CA3. In the frontal cortex, a high immunoreactivity for PSD-95 was observed at the tissue level, i.e. in neurons and neuro-pile. Qualitative observation revealed first, a diffuse signalling corresponding to postsynaptic expression as well as dendritic staining, visible throughout the whole cortex, (Figure 2a, left), and second, an increase of PSD-95 expression in AI compared with AU rats, particularly in the middle cortical layers, with neuronal bodies appearing increasingly well stained (Figure 2a, right). Image J analysis of all PSD-95 stained elements, including neuropil and neur-onal cell bodies throughout the whole cortex, confirmed these observations. Indeed, a one-way ANOVA comparing PSD-95 concentrations in the frontal cortex of the three groups showed a significant group effect (F (2,23) = 3.545; P = 0.046). Separate post hoc analyses (Bonferroni) showed (Figure 3a) that AI had a significantly higher level of PSD-95 than young rats (P = 0.045), whereas AU did not differ from the young rats (P = 0.823). The difference between AI and AU was not significant (P = 0.239), as expected from individual data distribution. Individual data emphasize the increased expression of PSD-95 in the AI rats, together with very small variability compared with other groups (Figure 3b).
Photomicrographs of 20-μm paraffin sections immunostained with for PSD-95. For localization of sections, see Materials and methods. (a) From left to right: low magnification (scale bar: 100 μm) of the frontal cortex in Aged Unimpaired rat (AU), Aged Impaired rat (AI), and two higher magnifications (scale bar: 10 μm/top, 5 μm/bottom) in layer 5 of the frontal cortex in one AI rat. Note that staining is increased in the AI rat, particularly in the middle layers, and higher magnification confirms that both neuronal bodies (arrows), dendritic segments (arrow heads) and puncta corresponding to synapses are well marked in AI rat. Image J analysis throughout the whole cortex indicates a significant increase in PSD-95 staining in AI rats. (b) Hippocampus in AU rat (left) and AI rat (right). Scale bar: 200μm. Image J analysis of subregions reveals no significant difference (see data). PSD-95, postsynaptic density protein-95. (A color version of this figure is available in the online journal) (a) PSD-95 (mean ± SEM) expression level measured by Image J intensity of optical density in the frontal cortex (FC) of young (n = 7), Aged Impaired (AI, n = 7) and Aged Unimpaired (AU, n = 12) rats. The value in AI rats is significantly different (*) from that in young, but not in AU rats. (b) Correlation between PSD-95 optical density in frontal cortex and percent of time spent in the training zone during the probe trial in young (n = 7) and aged rats (12 AU and 7 AI). The threshold (35.70% time in the training zone) for classification as AI or AU is indicated by the dashed line crossing the x-axis, as explained in the Materials and methods. PSD-95, postsynaptic density protein-95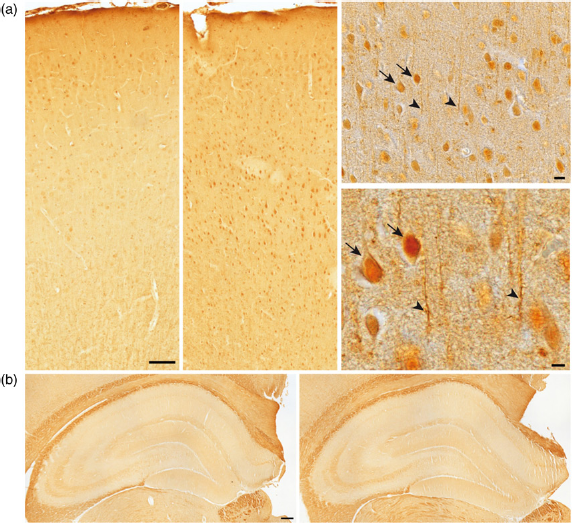
Western blot expression of PSD-95 and other proteins in AI and AU groups
Immunohistochemical data for PSD-95 were corroborated by Western blot analyses in the frontal cortex of representative samples of rats, as shown in Figure 4a. Similarly to immunohistochemical data, there was no significant difference for PSD-95 expression across groups in the hippocampus (data not shown).
(a) Fresh tissue of the frontal cortex from four Aged Unimpaired (AU; 1-4) rats and four Aged Impaired (AI; 5-8) rats was used for immunoblot analysis. The Coomassie blue stained gel on the right, with molecular weights indicated in kDa, shows equal protein loading. Western blots on the left show membranes stained for MAP2 (clone C), phosphorylated neurofilament proteins (SMI31), PSD-95, Tau (BR39), Tubulin (tu9b), GFAP, synaptophysin and SNAP25 (SMI81). The increase in PSD-95 and synaptophysin in the AI rats is clearly visible. (b) Results of the Western blot analysis for PSD-95, synaptophysin and tubulin (mean ± SEM), in the frontal cortex of eight aged rats (4 AI and 4 AU). y-axis: integration units (IU) of optical densities of gray values. Tubulin was used as reference and to correct values for loading differences. Significance of statistical comparisons is indicated by **(P <0.01) or *(P<0.05). (c) Correlation between PSD-95 level (units as in b) in frontal cortex and percent of time spent in the training zone during the probe trial of eight aged rats (4 AI and 4 AU). Illustrations of the swim paths of these rats are presented in Figure 1b. PSD-95, postsynaptic density protein-95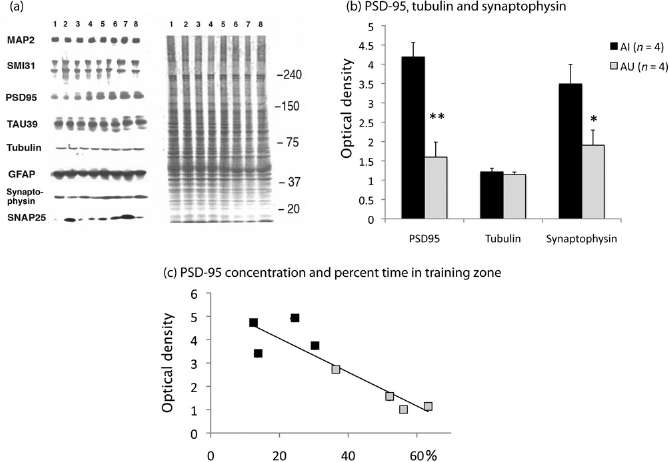
Western blot analyses (Figures 4a and b) in the frontal cortex of eight aged rats’ brains (among the four worst AI and four best AU) revealed a higher level of PSD-95 in the AI frontal cortex (unpaired Student's t-test, (t(6) = 4.81; P = 0.003). Synaptophysin concentrations were also significantly higher (t(6) = 2.47; P = 0.048) in AI (mean 3.49 ± SEM 0.51) compared with AU rats (mean 1.9 ± SEM 0.39). In contrast, there was no difference between AI and AU rats for tubulin, Tau, MAP2, SNAP25, NF-M, NF-H and GFAP (P > 0.05; Figure 4a).
Correlation between histological and Western blot changes and spatial accuracy
We found a negative correlation between PSD-95 expression and the percent time spent in the training zone by all the aged rats (n = 19; Figure 3b). This was confirmed by a Spearman test indicating that among these rats, PSD-95 expression level in the frontal cortex was inversely correlated to the percent time spent in the training zone (r = –0.549; two-tailed, P = 0.015). In the young rats, this correlation was not significant (r = 0.238; P = 0.57). A similar analysis between PSD-95 expression and latencies during the training trials at a new position (mean of trials 2–4 of escape latencies on the new position over the 2 sessions) showed no significant correlation, either in young (r = -0.357; two-tailed, P = 0.432) or in old rats (r = 0.047; two-tailed, P = 0.847).
Individual PSD-95 concentrations assessed by Western blot analysis of eight aged rats’ brains (among the four worst AI and four best AU) were negatively correlated with spatial accuracy (percent of time spent in the training zone; Figure 4c). This was confirmed by a Spearman test showing a negative correlation between PSD-95 concentrations and the time spent in the training zone (r = -0.857; two-tailed, P = 0.007). No significant correlation was found between PSD-95 and escape latencies (acquisition or new position learning latencies). Spearman tests indicated that synaptophysin concentration was correlated neither to the time spent in the training zone nor to latencies (acquisition or new position).
Discussion
Our approach relies on a selection of cognitively preserved aged subjects, clearly focusing on the training location during the probe trial whereas the remaining old subjects swam in an apparently disorganized manner. Our method discriminates aged subjects with preserved spatial accuracy – swimming in the exact area of the training position – from those with a lack of focusing during the probe trial and insists on selecting aged rats with accurate spatial focusing. Our selection allowed a differential analysis of functional aging with three main results.
First, we found that functional aging assessed by differential spatial accuracy was related to different levels of protein PSD-95. Second, this difference was only evident in the frontal cortex and was correlated to the time spent in the training zone, not to the escape latencies. Third, related impairments were mainly evident in reduced accuracy in the probe trial at the end of the acquisition phase. If AI rats had prolonged latencies during acquisition, it was only transient, as there was no difference between AI and AU rats during new position learning, often considered as an indication of working memory efficacy (see for example ref. 26 ).
PSD-95 expression in aged brains
PSD-95 concentrations assessed by immunohistochemistry with Image J analysis and Western blots were significantly elevated in the frontal cortex – but not in the hippocampus – of the aged rats expressing a weak or mere absence of spatial bias and thus classified as cognitively impaired (AI). There was no systematic increase in the brain of old rats defined as cognitively unimpaired (AU). Moreover, PSD-95 in the frontal cortex was correlated – in both immu-nohistochemical and Western blots data – to the time spent in the training zone, but not to the prolonged escape latencies in individual rats, whether during the reference acquisition or new position learning. This absence of difference in latencies suggests that the deficit in accuracy during the probe trial in AI rats cannot be attributed to vision or motor deficits. The fact that correlation between PSD-95 and accuracy was very significant in the group of aged rats suggests that a common factor might be involved in the maintenance of a high accuracy and lower PSD-95. Presynaptic markers such as synaptophysin, but not SNAP25, were also increased in rats with the highest cognitive impairments. Therefore, both the increase in synaptophysin and PSD-95 expressions indicate a potential reorganization of the presynaptic and postsynaptic compartments. These changes might indicate either compensatory attempts or local dysfunctions related to aging.
Some authors, however,14,15 showed a significantly decreased PSD-95 expression in the prefrontal cortex of AI Fisher 344 rats, but only at the synaptic level in lamina 3 and by measuring postsynaptic densities. Their data take into account fluorescence measurements of synaptic puncta at a high magnification, while ours consider the overall PSD-95 immunohistochemical staining, including immunoreactive puncta, dendritic segments and neuronal somata. Moreover, Majdi et al.14,15 considered latencies, not spatial biases for dividing AI and AU rats. Van Guilder et al. 13 also found a decreased amount of PSD-95, however, in hippocampal synaptosomes of old impaired rats, based on another classification of cognitive impairment. In other studies, PSD-95 was shown to be upregulated in the hippocampus of trained mice but not in naïve caged controls 27 and PSD-95 knockout mice revealed impairment in spatial learning with enhanced long-term potentiation and absence of long-term depression, 17 suggesting that this postsynaptic protein might be implicated in information storage. In addition, the observation that the thickness of the postsynaptic density as well as PSD-95 expression are increased in rats assigned to enriched environment housing suggests also a role of PSD-95 in compensatory mechanisms.28–30 Thus beside age-related loss of synapses, dendritic spines and branches occurring in vulnerable cells, an increase of PSD-95 expression may happen in reactive neurons incurring neuritic outgrowth, as suggested by our previous data in human Alzheimer cases.18,19 However, synaptophysin and PSD-95 enhanced expression might represent an attempt rather than a successful compensation following synaptic loss and reorganization and PSD-95 might instead accumulate in a non-functional manner.
Hippocampus versus frontal involvement in spatial cognitive deficits
The fact that the spatial deficit was related to synaptic protein changes in the frontal cortex and not in the hippocampus does not appear in accord with the commonly admitted role of the hippocampus in spatial memory. Moreover, the observed reduction in spatial accuracy during the probe trial is not related to changes in escape latency, either during acquisition or in the expression of short-term memory, such as in the second trial to a new position which might be expected from hippocampal impairment. Although the integrity of the rat medial pre-frontal cortex seems unnecessary for active storage of spatial information, it is necessary for retrieving and using stored information.22,31 Frontal modifications in synaptic proteins might be involved in the focused searching assessed in the probe trial as in our data. Moreover, other papers discuss the potential role of the frontal cortex in spatial behavior. For instance, Terry et al. 32 have shown that age-dependent alterations in nerve growth factor-related proteins in the prefrontal cortex and in the hippocampus were implicated in spatial deficits in the Morris task, suggesting that a variety of neurochemical changes accompany brain aging in rats and that no simple direct links can be established with specific behavioral dimensions.
On the whole, our behavioral results emphasize the need for a more comprehensive and analytical view of the spatial inaccuracy assessed in the probe trial. In other words, the spatial deficits (or conversely the preservation of accurate spatial capacity) associated with aging must be qualified and quantified in order to approach the age-related rearrangement of the cognitive architecture in humans7,33 or in rats. 34 In the later work, we showed that in a radial maze, aged rats tend to prefer olfactory marked arms to the spatially correct ones, thus relying more systematically on local cues than on the arms’ distribution in space. This contrasts with the systematic spatial preference expressed by young adults in similar situations. 35 A subset of 10 aged rats used in the present experiment was tested in a radial maze, where they expressed a preference for olfactory marked arms over spatially correct ones (unpublished data). Interestingly, the preference was most clearly marked in the rats with a high PSD-95 concentration (i.e. a significant correlation between PSD-95 concentration and olfactory bias strength, r = 0.594, P = 0.035) and in the rats with the lowest spatial accuracy in the water maze (r = 20.756, P = 0.011). These preliminary data suggest that like brain connectivity, the behavioral performance undergoes some reorganization providing possible compensatory adaptation during aging.
Functional brain networks in aging
A unified frontal aging hypothesis suggests that the human frontal cortex – together with the fronto-striatal system – are the earliest aging targets, resulting in decreased executive functions affecting the whole process of cognition. 1 Accordingly, several studies have also pointed to the frontal importance for cognitive function in rodents. 22 Dendritic plasticity can decrease in the frontal cortex of aging rodents, particularly under stressful conditions,36,37 which could lead to some postsynaptic changes. Furthermore, PSD-95 has been identified as a regulator of dopamine-mediated synaptic and behavioral plasticity. 38 Aging deficits in the water maze have been related to several brain changes, including deterioration of dopa-minergic innervation in the neocortex, reduction in nerve growth factor-related proteins, L-arginine decrease in the frontoparietal cortex and synaptic alteration in the hippo-campus.32,39–41 Moreover, in humans, age-related deficits in spatial memory observed in a virtual Morris water maze have also been related to neurobiological changes in the prefrontal cortex. 42 In this context, our data bring new information related to synaptic proteins in the frontal cortex of aging rodents, as already shown in Alzheimer patients.18,43 A compensatory mechanism is possible, involving among others, an increase in presynaptic synaptophysin and postsynaptic PSD-95. However, more work is needed to define whether an increase in PSD-95 might also be followed by post-translational modifications that render the protein less functional or results in a lack of transport and accumulation in the soma.
Conclusion
Our results show that PSD-95 expression is increased in the frontal cortex of impaired aged rats. PSD-95 was inversely correlated in all aged rats with spatial accuracy while it was not the case in the group of young adults. This suggests some degree of cortical postsynaptic reorganization with aging and points to the involvement of aging-related alterations in brain networks including the frontal cortex. Further analysis of how these changes are related will allow a better understanding of the critical processes underlying pathological brain aging.
Footnotes
Acknowledgements
We thank Prof Pierre Magistretti for support. Financial and material support was institutional. Part of it was from a grant from the Swiss National Foundation for the Scientific Research 3100A0-105765 (Prof F Schenk).