
Abstract
SERPINB3 is a serine protease inhibitor with pleiotropic functions. It is involved in several physiological and pathological processes, where it appears to exert antiapoptotic effects. Little is known about its expression on immune system cells, the major players in mechanisms of viral defense and autoimmune disorders. The aim of this study was to characterize the expression of SERPINB3 on the surface of peripheral blood mononuclear cell subsets in both normal subjects and in patients with chronic viral infections and autoimmune diseases. Sixty-two patients were analyzed by flow cytometric analysis, including 45 with hepatitis C virus (HCV)-related chronic liver disease and 17 with systemic lupus erythematosus (SLE). SERPINB3 was expressed on B lymphocytes in 79% of the controls, in 32% of the HCV-infected patients and in none of the SLE patients. Surface localization of SERPINB3 was confirmed by confocal microscopy. SERPINB3 positivity was associated with CD27 reactivity (r = 0.98), but not to other activation molecules (CD69, CD71, CD86 and CXCR3). SERPINB3 is physiologically expressed on the surface of CD27+ B lymphocytes, but its expression is reduced in HCV viral infection and not detectable in SLE patients. These results may suggest a role for SERPINB3 in B-cell defects typically found in viral infections and autoimmune disorders.
Background
SERPINB3 (formerly known as squamous cell carcinoma antigen 1 or SCCA 1) is a member of the ovalbumin serine protease inhibitor family. 1 It is physiologically present in the spinous and granular layers of normal squamous epithelium and highly expressed in tumors of epithelial origin 2,3 and in primary liver cancer. 4,5 SERPINB3 has been identified as a cellular surface protein able to bind hepatitis B virus in the liver (HBV). 6,7 However, since peripheral blood mononuclear cells (PBMCs) can be infected by HBV, 8,9 it is possible that SERPINB3 can also behave like a virus receptor, allowing internalization of HBV. 10 Previous studies have described a cytosolic localization for this serpin, 11 but more recent data indicate an additional surface localization of the protein, expanding its role as a functional surface receptor or an active proteinase inhibitor. 12 Furthermore, serpins generally regulate different signaling pathways such as cytoskeletal rearrangement and apoptosis, 1 and experimental data indicate that SERPINB3 is involved in tumor growth by inhibition of apoptosis 13 and induction of cellular proliferation. In apoptosis, the involvement of SERPINB3 has been documented, 14 but the specific molecular target of this serpin still remains unknown. 15,16
Among chronic virus infections, hepatitis C virus (HCV) is one of the most successfully persistent hepatotropic viruses. HCV carriers may progress to a broad spectrum of liver diseases, as a consequence of complex interactions between viral and host factors. 17,18 Cellular immune response plays a crucial role in the immunopathogenesis of HCV-related chronic liver disease and in viral clearance impairment, 19,20 although the mechanisms are largely unknown. The functional specialization of B-cells is fundamental in antibody-mediated immune responses and it seems critical in some chronic inflammatory conditions, including persistent viral infections. 21 Chronic HCV infection is often associated with a large spectrum of B-cell abnormalities, such as aberrant proliferation and production of rheumatoid factor. 22 In addition, B-cells may provide a vehicle for HCV to evade humoral immune responses. 23
Large arrays of heterogeneous aberrations of the immune system have been described in autoimmune diseases. In systemic lupus erythematosus (SLE), in addition to other immunological disturbances, B-cell hyperactivity, characterized by the production of a broad variety of autoantibodies, represents a central feature. 24 Recently, a possible role of increased apoptosis and impaired clearance of dying cells has been suggested. 25 Interestingly, the loss of B-cell tolerance as well as B-cell activation and autoreactivity can be induced by viruses. 26,27
The aim of the this study was to explore SERPINB3 expression on different PBMC subsets both in normal subjects and in patients with impaired immune response, including chronic HCV infection and SLE, to assess the potential involvement of SERPINB3 in the deregulation of B-cell reactivity.
Methods
Patients
PBMCs were obtained from 45 patients with chronic HCV infection, 17 patients affected with SLE and 24 healthy subjects, enrolled as controls. All patients with chronic HCV infection had serological HCV-RNA positivity and liver disease defined as chronic active hepatitis, liver cirrhosis with or without hepatocarcinoma. Patients with SLE were classified according to the American College of Rheumatology (ACR) criteria. For term definitions of SLE-specific features, we used those included in the ACR criteria. 28 SLE disease activity was measured by European Consensus Lupus Activity Measurement (ECLAM) score 29 and ECLAM score ≥2 was considered indicative of active disease.
All patients and controls signed the informed consent for this study. The study was approved by the local ethics committee.
Cell lines and PEGylated interferon α treatment
The human hepatoblastoma HepG2 cell line transfected with SERPINB3 gene (HepG2SB3) is described elsewhere; 30 the human hepatoblastoma HA22T/VGH cell line was kindly provided by Professor G De Petro (University of Brescia, Italy); and the lymphoblast-like Raji cell line was provided by Dr S Indraccolo (University of Padua, Italy). The HepG2SB3 cell line was cultured in minimum essential medium (MEM; Sigma-Aldrich, St Louis, MO, USA); HA22T/VGH was maintained in Dulbecco's modified medium (Sigma-Aldrich); and the Raji cell line was cultured in RPMI medium (Euroclone, Pero, Italy). The media were supplemented with 10% fetal bovine serum, 100 U/mL of penicillin, 100 mg/mL of streptomycin, 200 mmol/L of L-glutamine and 1× of MEM non-essential amino acid solution (Sigma-Aldrich). The cultures were routinely maintained at 37°C in a 5% CO2/95% air atmosphere.
For PEGylated interferon α (IFN-α)-based experiments, the cells were seeded onto six-well plates (5 × 105 cells/well), and incubated with different concentrations of drug (PegIntron; Schering Plough, Kenilworth, NJ, USA) for 30 min, 6, 24 or 48 h. At the end of the treatment, the cells were harvested and pellets used for molecular biology studies. PEGylated IFN-α concentration of 1 μg/mL was used for subsequent studies with Raji and HepG2SB3 cell lines and an incubation time of 24 h for drug treatment was selected for SERPINB3 mRNA modulation studies.
Antibodies and reagents
For cell surface marker analysis, cells were stained with the following monoclonal antibodies: anti-human CD4 phycoerythrin cyanine 5 (PE Cy5.5), anti-human CD8 phycoerythrin (PE), anti-human CD16 PE, anti-human CD14 PE, anti-human CD19 PE Cy5.5, anti-human CD27 allophycocyanin (APC), anti-human CD69 fluorescein isothiocyanate (FITC), anti-human CD71 FITC, anti-human CD86 PE and anti-human CXCR3 FITC (BD Bioscience, San Josè, CA, USA). For SERPINB3 indirect immunostaining, the monoclonal anti-human SCCA antibody (Mab 6E12V; Xeptagen S.p.A., Marghera, Venice, Italy) was used at a concentration of 8 μg/mL and the polyclonal rabbit anti-mouse IgGs FITC (DakoCytomation, Glostrup, Denmark) was used at the manufacturer's recommended concentrations. Isotype-matched control antibodies were included in all the experiments.
For confocal microscopy analysis, mouse anti-human SCCA (Mab 6E12V; Xeptagen S.p.A.), mouse anti-human IgG (Caltag, Burlingame, CA, USA) and goat anti-mouse IgGs Alexa Fluor 488 (Molecular Probes, Eugene, OR, USA) were used.
Cell separation
PBMCs were isolated from freshly drawn heparinized blood by Ficoll Hypaque density gradient separation (Sigma-Aldrich). Cell viability was evaluated by trypan blue dye exclusion. Total B lymphocytes were positively selected from PBMCs with magnetic microbeads conjugated to anti-human CD19 and MACS columns (Miltenyi Biotec, Bergish Galdbach, Germany), according to the manufacturer's instructions. Fluorescence-activated cell sorting (FACS) staining for the B-cell marker CD19 and for the T-cell marker CD3 was used to control cell purity. This procedure usually yielded >95% CD19-positive B-cell preparations.
Flow cytometric analysis
Surface expression of SERPINB3 in PBMCs was assessed by FACS analysis. To characterize the cell subsets reactive for the serpin, the following panel of antibodies was simultaneously used for direct immunofluorescence staining: anti-CD4 PE Cy5.5, anti-CD8 PE, anti-CD16 PE, anti-CD14 PE and anti-CD19 PE Cy5.5. Appropriate isotypic controls were run for all the antibodies used. For SERPINB3 characterization, mononuclear cells (at 0.5 × 106 cells/mL) were incubated with mouse anti-SCCA antibody at room temperature for 15 min, followed by FITC-conjugated rabbit anti-mouse IgGs in a phosphate-buffered saline solution containing 0.1% NaN3 for 15 min. Phenotypes of PBMC were assigned to CD8, CD16 and CD14 using PE-conjugated antibodies and to CD4 and CD19 using PE Cy5.5-conjugated antibodies. Cells incubated with unconjugated anti-IgG, as isotypic controls, were included in each experiment. For each analysis, 30,000 events were acquired.
Immunofluorescence
Purified B lymphocytes were incubated with an anti-human CD19 FITC antibody, mounted on poly-
HepG2SB3 cells treated or not treated with PEGylated IFN-α for 48 h were seeded on slides (2 × 105/slide) and incubated with 8 μg/mL of rabbit anti-SERPINB3 primary antibody (Hepa antibody; Xeptagen), washed and incubated with Alexa Fluor 546-conjugated secondary antibody (Invitrogen, Carlsbad, CA, USA). Cellular nuclei were counter-stained with DAPI (4′,6-diamidine-2′-phenylindole dihydrochloride; Roche Applied Science, Mannheim, Germany). The slides were mounted with ELVANOL (Sigma-Aldrich) and observed under a fluorescence microscope (Axiovert 200M; Carl Zeiss MicroImaging GmbH, Gottingen, Germany).
SERPINB3 association with activation molecules
To define the profile of B lymphocytes expressing SERPINB3 on their surface, different activation molecules were explored in parallel using the following panel of monoclonal antibodies: anti-CD27 APC, anti-CD69 FITC, anti-CD71 FITC, anti-CD86 PE and anti-CXCR3 FITC. The labelling procedure was carried out as described before. The cells were scored using a FACS Calibur (BD Biosciences, Franklin Lakes, NJ, USA) and the total population of viable cells was gated according to their typical forward and right-angle light scatter. A total of 100,000 events were acquired for each analysis.
Reverse transcription and quantitative realtime polymerase chain reaction
Total RNA, both from purified B lymphocytes (CD19+) of healthy controls, 10 HCV-infected patients and 10 patients with SLE and from in vitro cultured cell lines, was extracted using RNasy Trizol (Invitrogen) according to the manufacturer's instructions and quantified by NanoDrop (Thermo Fisher Scientific, Wilmington, DE, USA). Total RNA (up to 1 μg) was reverse-transcribed using Superscript II reverse transcriptase (Invitrogen) following the manufacturer's instructions. The levels of SERPINB3 mRNA were measured by realtime polymerase chain reaction (PCR), using the SYBR® green method (Roche Applied Science). The designed oligonucleotide sequences of the primers were as follows. SERPINB3: sense, 5′ GCA AAT GCT CCA GAA GAA AG 3′, reverse, 5′ CGA GGC AAA ATGAAAA AGA TG 3′; 2′–5′-OAS: sense, 5′ CAA GCT CAA GAG CCT CAT CC 3′, reverse, 5′ TGG GCT GTG TTG AAA TGT GT 3′. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the internal reference and the oligonucleotide sequences were as follows: sense, 5′ TGG TAT TCG GGA AGG ACT CAT GAC 3′, reverse, 5′ ATG CCA GTG AGC TTC CCG TTC AGC 3′. The single-tube realtime PCR assays were performed using the LightCycler instrument (Roche Diagnostics GmbH, Indianapolis, IN, USA). To assess SERPINB3 mRNA concentration in purified B lymphocytes, a standard curve was generated using serial six-fold dilutions of a plasmid containing the SERPINB3 insert and the results were expressed as copy number/100 ng of total RNA.
Data analysis
Data were processed using the Macintosh CellQuest software program (Becton Dickinson Biosciences, San Jose, CA, USA). The results were analyzed using Fisher's test, Mann-Whitney U test and Pearson's correlation, when appropriate. A normality test was performed by the Kolmogorov–Smirnov test. Two-tailed P values were calculated for all tests and P < 0.05 was considered significant. Statistical analyses were carried out using statistical packages GraphPad View version 4.0 (GraphPad Software, San Diego, CA, USA) and Statistical version 7.1 (StatSoft Italia, Padova, Italy).
Results
Clinical and demographic features of patients and controls
Chronic HCV infection
Forty-five patients with chronic HCV infection were enrolled: 14 with chronic active hepatitis (7 men, 7 women, mean age ± SD 50.4 ± 17.1 y), 17 with liver cirrhosis (8 men, 9 women, mean age ± SD 59.5 ± 13.7 y) and 14 cirrhotic patients with hepatocellular carcinoma (7 men, 7 women, mean age ± SD 72.5 ± 6 y). None of the patients received antiviral treatment in the last five years or had mixed cryoglobulinemia.
SLE
Seventeen SLE patients were enrolled (5 men, 12 women, mean age ± SD 40 ± 5.1 y). The overall prevalence of the prominent SLE clinical and laboratory features was: arthritis 94%, skin rash 71%, serositis 41%, glomerulonephritis 71%, neuropsychiatric manifestations 35%, hematological abnormalities 94%, antinuclear antibodies 100%, antidouble-stranded (ds)DNA 82%, anti-Sm 29%, anti-U1RNP 29%, anti-SSA 23%, anticardiolipin 35% and lupus anticoagulant 18%.
The mean ECLAM score ± SD of SLE patients was 3.29 ± 1.58, and 13 patients (76.5%) had ECLAM score ≥2. As far as the treatment at the time of blood sample was concerned, twelve patients (71%) were taking prednisone: four patients at a dosage >5 mg/day and eight patients ≤5 mg/day (five patients were prednisone-free), six patients were taking immunosuppressants (35%) and six patients took hydroxychloroquine (35%).
Demographic and clinical characteristics of the study population
HCV, hepatitis C virus; CH, chronic hepatitis; HCC, hepatocellular carcinoma; SLE, systemic lupus erithematosus; AFP, alphafetoprotein; ALT, alanine aminotransferase; gamma-GT, gamma-glutamyltranspeptidase; n.a., not available; PLTs, platelets; SD, standard deviation
SERPINB3 surface expression in different cell subsets
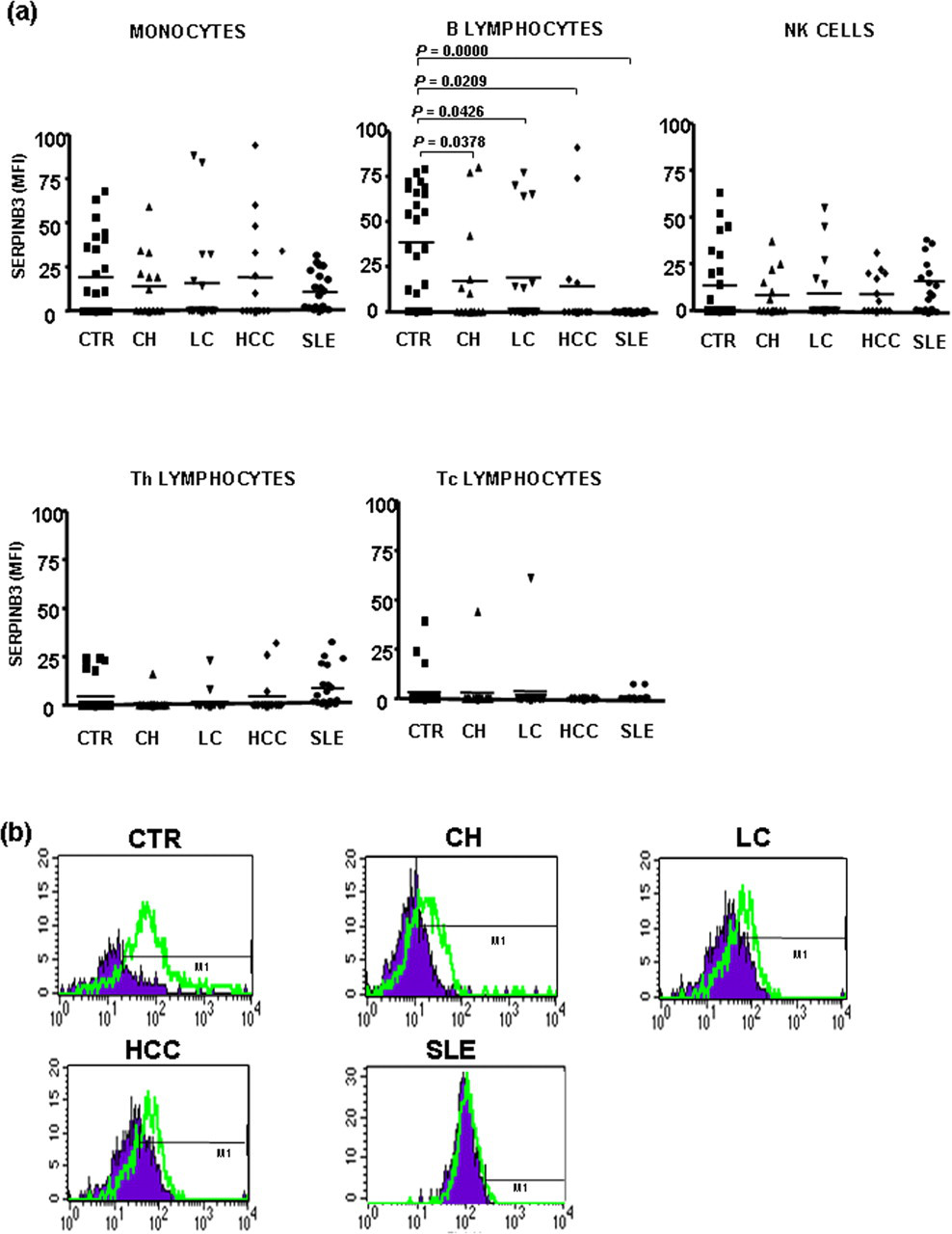
SERPINB3 was detected on the surface of B lymphocytes in 79% of the healthy subjects, while the percentage of surface positivity in the same cell subset was significantly decreased (32%) in patients with chronic HCV infection, independently of the disease stage, and was absent in SLE patients. No significant differences were observed between HCV and SLE patients versus controls in the other cell subsets (Figure 1). Beside the highest frequency of SERPINB3 on the surface of B lymphocytes, healthy subjects also showed the highest levels of serpin expression in this cell subset, compared with HCV and SLE patients (Figure 2).
SERPINB3 expression and distribution in different mononuclear cell subsets. Flow cytometric analysis of surface distribution of SERPINB3 positivity in different mononuclear cell subsets of the study population, including healthy controls, different groups of HCV-infected patients and SLE patients. To characterize the cellular subset, a panel of different antibodies was simultaneously used (anti-CD4, anti-CD8, anti-CD14, anti-CD16 and anti-CD19). Values of P were calculated by the Fisher's exact test. Only statistically significant differences were reported in the figure. HCV, hepatitis C virus; SLE, systemic lupus erythematosus SERPINB3 expression and distribution on mononuclear cell subsets. (a) SERPINB3 distribution on mononuclear cell subsets in different groups of patients and in controls. Surface SERPINB3 reactivity was expressed as mean fluorescence intensity (MFI). Lines across each group represent the median value. Values of P were calculated by the Mann-Whitney U test. Only statistically significant differences were reported in the figure. (b) Representative FACS analysis of the expression of SERPINB3 on the surface of B lymphocytes in different groups. CTR, controls; CH, chronic hepatitis; LC, liver cirrhosis; HCC, hepatocellular carcinoma; SLE, systemic lupus erythematosus; FACS, fluorescence-activated cell sorting. (A color version of this figure is available in the online journal)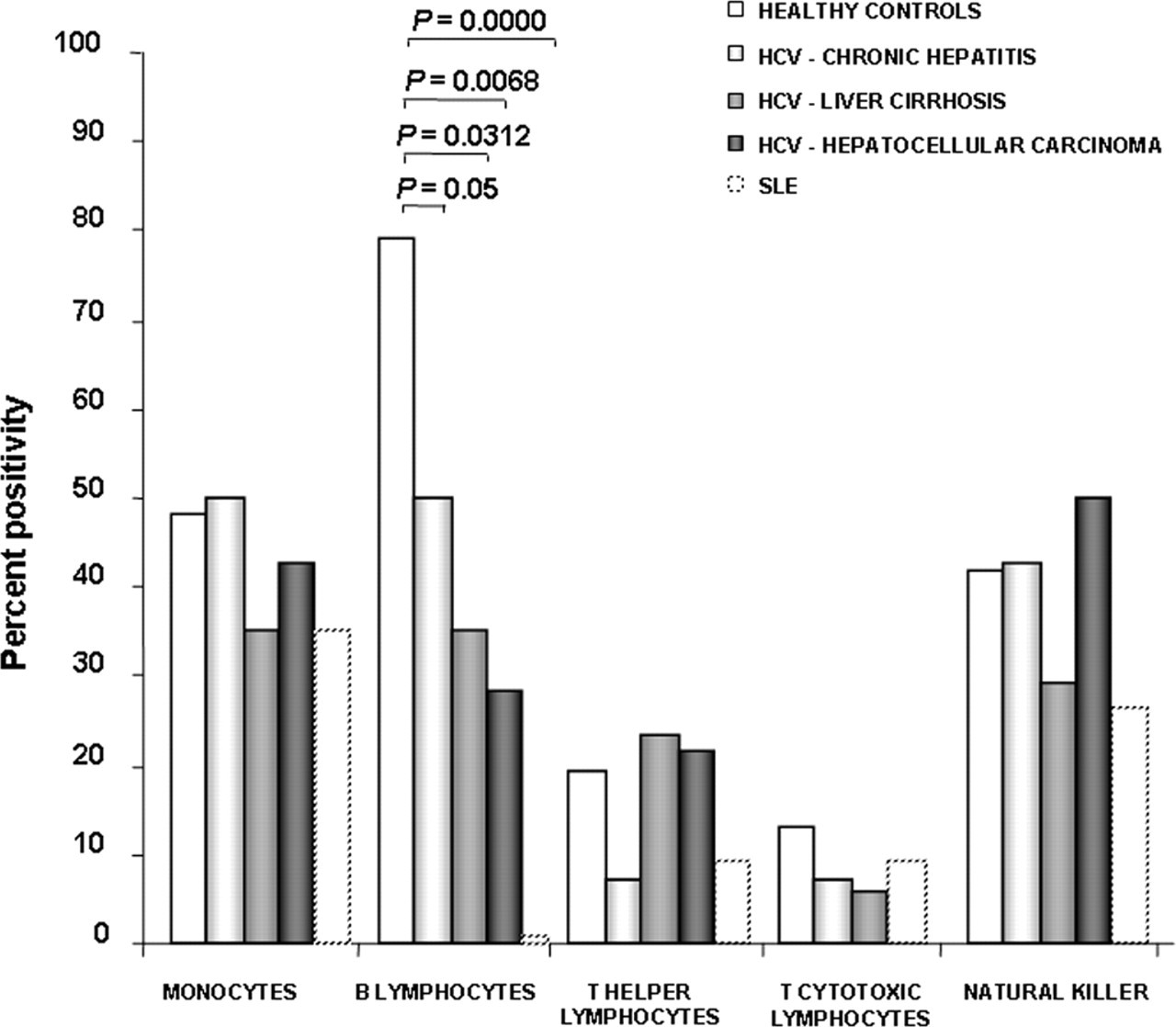
Surface localization of SERPINB3 on B lymphocytes was also confirmed by confocal microscopy, as shown in Figure 3a.
Surface localization of SERPINB3 and its mRNA quantification on purified B lymphocytes. (a) Surface localization of SERPINB3 on purified B lymphocytes by confocal microscopy. Cells were stained with an anti-SCCA antibody (a3) and specificity of the staining was assessed by the use of an isotypic anti-Ig antibody as control (a1). Corresponding bright fields are shown on the right (a4, a2); original magnification ×600. (b) SERPINB3 RNA concentration extracted from purified B lymphocytes, measured by realtime polymerase chain reaction in healthy donors (controls), in HCV+ and in SLE patients. SERPINB3 RNA concentrations were assessed on the basis of a standard curve using a plasmid containing the SERPINB3 insert and normalized to total RNA concentration (copy number/100 ng RNA). Columns represent mean values ± SD. HCV, hepatitis C virus; SLE, systemic lupus erythematosus; anti-SCCA, squamous cell carcinoma antigen
SERPINB3 mRNA quantification in B lymphocytes
The extent of SERPINB3 transcription in B lymphocytes, expressed as SERPINB3 plasmid copy number, was significantly higher in healthy subjects than in HCV-infected patients and in SLE patients, where trivial levels were detected (Figure 3b).
SERPINB3 expression is correlated with B lymphocyte activation molecules
No significant differences were found in the expression of CD69, CD71, CD86 and CXCR3 molecules, both in CD19+/SERPINB3+ cells (Figure 4a) and in CD19+/SERPINB3− cells (Figure 4b). However, the percentage of CD27 positivity was significantly higher in the SERPINB3+ B-cell population of healthy subjects than in those of HCV-infected patients (P < 0.05). No differences were observed between the two groups in SERPINB3− B-cells. The higher frequency of CD27 positivity on the surface of SERPINB3+ B lymphocytes was associated with higher levels of CD27 expression in healthy subjects, compared with HCV-infected patients (P = 0.019), while no differences were found in SERPINB3− B lymphocytes. In addition, a direct correlation (r = 0.98) between the extent of SERPINB3 and CD27+ reactivity was documented (Figure 4c). In keeping with the above results, the analysis of the expression of CD27 on B lymphocytes revealed that the frequency of CD27+ B-cells in healthy subjects was significantly higher than that in HCV-infected patients, and was negligible in SLE patients (Figure 4d).
SERPINB3 expression is correlated with B lymphocyte activation molecules. (a, b) Mean distribution of different activation markers on B lymphocytes in cases positive for SERPINB3 on B lymphocytes. The columns represent the percentage of each activation molecule in CD19+/SERPINB3+ (a) and in CD19+/ SERPINB3− (b) gates. Values of P were calculated by the Mann-Whitney U test. *P < 0.05. Only statistically significant differences were reported in the figure. (c) Relationship between the expression of CD27 and SERPINB3 on B lymphocytes. The correlation between the percentage of CD19+/SERPINB3+ cells and of CD19+/CD27+ cells in individual samples was carried out considering the overall study population. (d) Distribution of the frequency of CD27 reactivity on B-cells by FACS analysis in healthy controls and in patients with HCV infection and SLE. Horizontal bars represent the median value for each group. Values of r and of P were calculated by the Pearson correlation and by the Mann-Whitney U test, respectively. **P < 0.01; ***P < 0.0001. HCV, hepatitis C virus; SLE, systemic lupus erythematosus; CTR, controls; FACS, fluorescence-activated cell sorting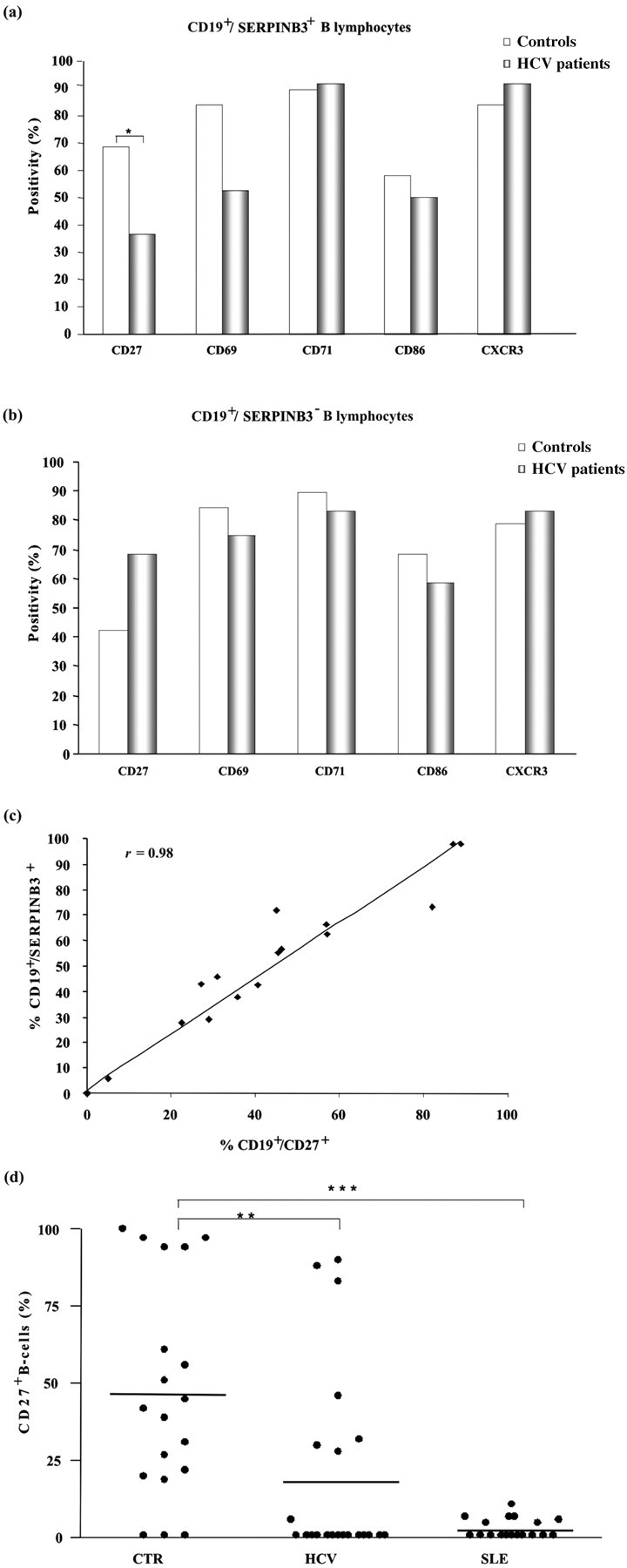
SERPINB3 mRNA early modulation following PEGylated IFN-α treatment
To assess the possibility that differential surface expression levels of SERPINB3 were correlated to an altered microenvironment able to interfere with SERPINB3 active transcription, we conducted a series of in vitro experiments using three different cell lines treated or not with PEGylated IFN-α. HA22T/VGH, Raji and HepG2SB3 cell lines were treated for 24 h with 1 μg/mL of the drug and SERPINB3 mRNA modulation was analyzed with realtime PCR. A sensor for IFN activity was the upregulation of 2′–5′-OAS mRNA. At early treatment time points, we observed a strong down-modulation (expressed in %) of SERPINB3 mRNA levels in all three cell lines studied, compared with controls, as reported in Figure 5a. At 48 h of treatment, a down-regulation of protein expression was also appreciable, as shown in Figure 5b.
SERPINB3 mRNA early modulation following PEGylated interferon-α treatment. (a) Left: down-regulation of SERPINB3 mRNA in HA22T/VGH, Raji and HepG2SB3 cell lines treated for 24 h with 1 μg/mL of PEGylated interferon α. Realtime polymerase chain reaction results are expressed as % of fold decrease compared with controls (untreated cells). Right: parallel analysis of up-regulation of 2′–5′ OAS mRNA expressed as % of fold increase in the same treated cell lines (compared with untreated controls). (b) SERPINB3 down-modulation (red) on HepG2SB3 cells after 48 h of PEGylated interferon α treatment. Nuclei were stained with DAPI (blue). Representative immunofluorescence images are shown. Corresponding bright fields are shown on the right; original magnification ×600. DAPI, 4′,6-diamidine-2′-phenylindole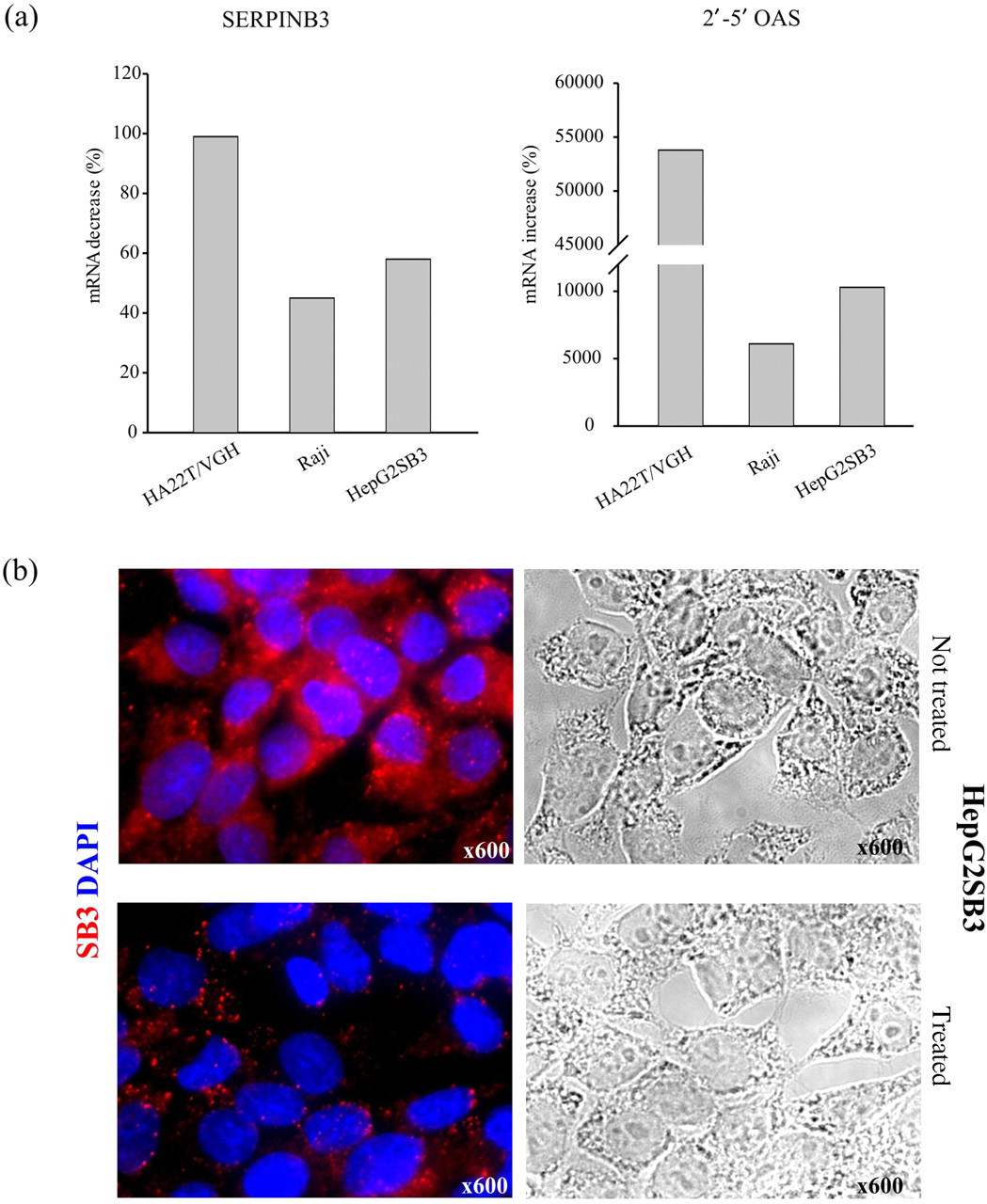
Discussion
Immune response plays a pivotal role in the control of autoimmune disorders and viral diseases. The present study was carried out to determine whether SERPINB3 was physiologically present on PBMC subsets and whether an abnormal expression of this serpin could be documented in viral and autoimmune diseases.
HCV persists in up to 70% of acutely infected individuals and it may develop several strategies to avoid immune clearance and establish persistent infection. The high rate of virus production during early infection may result in immune exhaustion and failure of the innate immune response to control virus replication. 31,32 Moreover, viral infections are accompanied by the establishment of an inflammatory milieu. 33 Proinflammatory signaling cascades are activated in HCV-induced hepatitis, for example. 34 Nevertheless, an endogenous activation of IFN-induced pathways in patients with chronic hepatitis C is reported. 35,36 IFN responses seem to be crucial to host antiviral defense but, at the same time, they represent targets for viral interference. 37 Moreover, such a milieu is linked to the set-up of an antiviral state, characterized by the secretion of cytokines and accruement of immune cells in the microenvironment. Several studies have shown the importance of T-cell-mediated immune response in controlling HCV infection, 38 while little is known about the role of the B-cell response in this setting, although chronic HCV infection is often associated with a large spectrum of B-cell abnormalities, 22 such as unusual activation and clonal expansion. 39 However, the mechanisms of HCV interaction with B lymphocytes remain largely undefined.
SLE is a chronic autoimmune disease in which defects of apoptosis and impaired clearance are contributing factors for susceptibility, onset and severity of the disease. 40 Deficiency in the clearance of apoptotic or necrotic cells, shown in patients with SLE, leads to accumulation of non-ingested nuclear material which may provide survival signals for autoreactive B-cells with the consequent production of antibodies against nuclear structures. 41 Apoptosis is important for the maintenance of both central and peripheral tolerance, typical features of a healthy immune system. 42 Interestingly, IFN signature seems to be one of the first steps in the pathogenesis of SLE and other autoimmune diseases. 43 IFN-α can contribute to the pathogenesis of SLE through various mechanisms, including direct and indirect effects on antigen-presenting cells, T-cells and B-cells. IFN-α is largely produced by immature dendritic cells (DCs), now known as plasmocytoid DCs (pDCs). These cells are stimulated by viruses to produce IFN-α and this production upregulates expression of Toll-like receptor 7 by B-cells, promotes cell death by increasing the release of certain RNA and DNA autoantigens and primes pDCs to respond more effectively to immunocomplexes. 44
In this study, SERPINB3 was described for the first time on the surface of B lymphocytes in the majority of healthy subjects, while HCV-infected patients revealed a marked reduction in its expression, independently of disease stage, and SLE patients were totally negative for protein surface expression. Realtime PCR results confirmed the actual transcription of the molecule in positive cells, excluding the possibility of its passive binding to the outside of the cell membrane. SERPINB3 detection on the surface of B lymphocytes corroborates the possibility of cellular localization of this molecule not only at the cytoplasmic 11 or nuclear level, 12 but also at the cell surface, and this localization supports its putative role as a cellular receptor for HBV. 6,10 The lack of SERPINB3 on B lymphocytes of patients with SLE could explain, at least in part, the lower frequency of HBV infection detected in this category of patients. 45 To further define the profile of B-cells bearing SERPINB3 on their surface, activation markers were analysed in SERPINB3-positive and SERPINB3-negative B-cell subsets. Flow cytometric analysis revealed that SERPINB3 expression was directly correlated with levels of CD27, a molecule of the tumor necrosis factor receptor family, typically associated with B-cell maturation and memory cell evolution. 46
In HCV-infected patients, the observed reduction in SERPINB3 expression with a parallel decrease in levels of CD27 at the cell surface of B lymphocytes might suggest a possible involvement of SERPINB3 in the deregulation of B-cell maturation. Alternatively, it may point to a dual effect of HCV infection, consisting of impairment in SERPINB3 surface translocation and B-cell maturation. HCV is indeed a lymphotropic virus, capable of replication in PBMCs of patients with chronic hepatitis C. 47
Conflicting results have been reported in the literature on CD27+ B lymphocyte frequencies in HCV-infected patients. Our results showed a significantly lower frequency of CD27+ B-cells in HCV patients than in healthy subjects. When expressed as a percentage of circulating total B-cells, an increase was described in HCV-infected patients, while lower frequencies, compared with controls, were reported by others, 48,49 suggesting lack of antigen-driven activation or proliferation of an expanded B-cell population. Our results are in keeping with the latter findings, since fewer CD27+ memory B-cells were found in HCV-infected patients compared with healthy controls. The role of B-cells in HCV infection has been mainly neglected over the years, but in a recent review, the multifaceted functions of B-cells in chronic HCV infection were considered, pointing out the importance of this ‘non-classical’ immune response. 22
The present study has also documented an impaired SERPINB3 expression in SLE patients, resulting in its absence on the surface of B lymphocytes. Previous data showed altered signaling mechanisms in B lymphocytes in SLE patients. 50 Defects in protein expression and function in specific pathways could support the complexity of SLE disease and its heterogeneity in different groups of patients. In our analysis, SLE patients showed a remarkable decrease in CD27+ B-cell frequency, compared with healthy subjects. These results might be interpreted in light of the substantial alterations in the distribution of different B-cell subsets commonly observed in SLE. Indeed, the lack of CD27+ B-cells seen in SLE patients may be due to lower numbers of CD27+IgM+ IgD+ unswitched memory cells in these patients, especially in those with active disease. 51
Moreover, the in vitro results, which showed an early down-modulation of SERPINB3 gene transcription and protein expression after an IFN-based treatment, could, in part, be translated into the in vivo context we can find in cases of autoimmune diseases. In such a microenvironment, characterized by the switch-on of IFN-driven pathways, the deregulation of the SERPINB3 transcriptional program, confirmed by an altered protein expression on the B-cell surface of both HCV and SLE patients, could be responsible for the impaired maturation of B-cells.
Notably, alfa1-antitrypsin, which is, like SERPINB3, a member of the ovalbumin serin protease inhibitor family, administered to MRL/lpr lupus-prone mice induced a decrease in proteinuria and hematuria as well as serum levels of interleukin (IL)-17, IL-12 and anti-dsDNA antibodies. 52 Thus, serpin family members seem to exert immunomodulatory effects and their administration might be helpful in conditions of immune system dysregulation, such as SLE.
Conclusions
In summary, considering the reported findings, SERPINB3 is physiologically present on the surface of CD27+ B lymphocytes and its expression is impaired in chronic HCV infection and in SLE, with a parallel effect observed in CD27+ reactivity. Further investigations are required to better define a potential involvement of SERPINB3 in B-cell defects, often present in viral and autoimmune diseases.
Footnotes
ACKNOWLEDGEMENTS
The authors are deeply grateful to Professor JM Dayer (Clinical Immunology Unit, Geneva, Switzerland) for his helpful discussion and critical reading of the manuscript and to Dr G Fassina (Xeptagen, S.p.A.) for anti-SCCA antibody supply. This study was supported, in part, by grants from the National Ministry of Education, University and Research (FIRB Project Prot. RBLA03S4SP_005) and from the Associazione Italiana per la Ricerca sul Cancro AIRC (Project No. 10235).