
Abstract
Tumor stroma plays a prominent role in cancer progression. Fibroblasts constitute a majority of the stromal cells in tumor, and yet the functional contributions of these cells to tumor angiogenesis and invasion are poorly understood, especially the anticancer drug interference to these processes. To estimate the effects of vinorelbine (VNR) on fibroblast-associated tumor invasion and angiogenesis, we evaluated the response of 95D and human umbilical vein endothelial cell (HUVEC) migration, tube formation in vitro, as well as capillary formation of rat thoracic aorta rings to hypoxic MRC-5 conditioned medium (CM) by VNR pretreatment. Our results demonstrated that VNR significantly inhibited 95D and HUVEC migration and angiogenesis induced by hypoxic MRC-5 cells. We also showed that hypoxic MRC-5 CM (Hypo-CM) had a higher level of stromal cell-derived factor 1 (SDF-1) secretion, while Hypo-CM up-regulated the CXCR4 expression in HUVECs and 95Ds. This increased activity of SDF-1/CXCR4 paracrine was clearly attenuated by VNR pretreatment. It was further found that pretreating HUVECs and 95Ds with AMD3100, a CXCR4 antagonist, markedly reversed the Hypo-CM promoting cell migration and angiogenesis, while adding exogenous SDF-1 attenuated the inhibition effects of CM collected from VNR-pretreated hypoxic MRC-5 (Hypo-CMV). These data indicate that VNR indirectly decreased 95D migration and angiogenesis through its effect on hypoxic MRC-5, via impacting SDF-1/CXCR4 paracrine, suggesting that VNR could interrupt the influence of fibroblasts on HUVECs and 95Ds to exert an anticancer role. Therefore, fibroblasts should be taken into consideration when evaluating and developing anticancer drugs.
Introduction
Genetic and cell biology studies indicate that tumor progression is not only determined by malignant cancer cells themselves, but also by the tumor stroma. 1 In most human tumors, the stroma microenvironment is heavily altered compared with the stroma of normal tissue. 2 The tumor stroma is thought to be composed of endothelial cells, fibroblasts, perivascular cells and inflammatory cells, which regulate critical components of the tumorigenic process: angiogenesis, desmoplasia and inflammation. 3
Within the tumor stroma, fibroblasts are the most abundant and of outstanding importance. Activation of fibroblasts with various stimuli, such as transforming growth factor-β, thrombin, interleukin (IL)-4, IL-13 or hypoxia, induces the expression of α-smooth muscle actin (α-SMA), which is considered to be a marker of myofibroblasts. 4,5 So, activated fibroblasts (which are sometimes termed as myofibroblasts) that are found to be in association with cancer cells are known as carcinoma-associated fibroblasts (CAFs). 6 CAFs secrete a variety of growth factors and proteinases facilitating tumor angiogenesis and invasion. 6 Given the important functions of fibroblasts, more attention should be paid to the influence of anticancer drugs on them. While recent studies focusing on CAFs have begun to uncover their prominent roles in promoting tumor angiogenesis and invasion, the specific mechanisms of these fibroblasts in tumor–stroma interactions are still not clear. 7 However, accumulating evidence indicates that the stromal cell-derived factor-1 (SDF-1)/CXCR4 pathway is of particular importance in the participation of fibroblasts in angiogenesis and invasion. 8 The chemokine SDF-1 (also known as CXCL12) is constitutively expressed and produced by fibroblasts. 9 The cognate receptor for SDF-1 is CXCR4 (also known as CD184), which is widely expressed by numerous cells in tumors. 10 Recent studies have revealed that hypoxia, particularly hypoxia-inducible factor 1α (HIF-1α), regulates the SDF-1/CXCR4 signal which is widely confirmed to contribute to the metastatic potential of tumors. 11,12
Vinorelbine (5′nor-anhydro-vinblastine) (VNR) is a semi-synthetic vinca-alkaloid that is manufactured from alkaloids extracted from the rosy periwinkle, Catharanthus roseus. 13 VNR induces cytotoxicity by inhibiting the polymerization of tubulin dimers into microtubules, which in turn disrupts mitotic spindle formation and prevents cell division. 14 VNR is rarely used in the first-line setting, with the exception being its use in combination with cisplatin and cetuximab, the epidermal growth factor (EGF) receptor antagonist. 15,16 As the treatment of all cancer becomes increasingly targeted and personalized, the role of VNR is likely to change. Besides its well-known functions on cancer cells, VNR has been found to inhibit angiogenesis via its direct toxic effect on vascular endothelial cells. Clinical research has also indicated that VNR administered at a metronomic dose without overt toxicity has the potential to serve as a novel angiogenesis-targeted therapy, 17 which initiates the requirement for further investigation of the thorough actions of VNR on tumor stroma.
In the current study, in order to explore the effects of VNR on the tumor stroma, we investigated the effects of fibroblasts on the angiogenesis and migration of cancer cells, as well as the possible molecular mechanisms underlying these events. We accordingly demonstrated that VNR could affect the fibroblasts to exert a complementary anticancer activity. A better understanding of the changes that occur in fibroblasts in response to the presence of malignant cells, human umbilical vein endothelial cells (HUVECs) and anticancer drugs, may lead to the development of new strategies for cancer treatment.
Materials and methods
Cell line and cell culture
The human fetal lung fibroblast cell line MRC-5 was obtained from the Cell Bank of the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences (Shanghai, China). MRC-5 cells were cultured in minimum essential medium (MEM; Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS; Gibco), 100 U/mL penicillin, 100 μg/mL streptomycin and 1% non-essential amino acids. Human lung cancer cell line 95D (high metastasis potential) was obtained from the Cell Bank of the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, and cultured in RPMI-1640 medium (Sigma, St Louis, MO, USA) supplemented with 10% FBS. HUVECs were isolated from human umbilical cord veins by collagenase treatment as described previously. 18 The harvested cells were grown in medium 199 (M199; Gibco) containing 20% FBS, 30 μg/mL endothelial cell growth supplement (Sigma), 10 ng/mL EGF (Sigma), 100 U/mL penicillin and 100 μg/mL streptomycin. After three to five passages, HUVECs were collected for use in all experiments. The cells were incubated in a humidified atmosphere of 95% air plus 5% CO2 at 37°C.
Preparation of conditioned medium
The MRC-5 cells were cultured in MEM/10% FBS in normoxic conditions. When the cells grew to 70–80% confluence in 100 mm diameter culture dishes, the medium was changed to fresh serum-free MEM in the absence or presence of VNR (0.1, 1 and 10 μmol/L). The medium was replaced with fresh serum-free MEM after 12 h and the cells were incubated under hypoxia (1% O2, 5% CO2 and balanced N2) for 12 h, and the control cultures were incubated in 95% room air and 5% CO2. Then the conditioned medium (CM) from normoxic MRC-5 cells (Norm-CM), hypoxic MRC-5 cells (Hypo-CM) and hypoxic MRC-5 cells pretreated by 0.1, 1 and 10 μmol/L VNR (Hypo-CMV [0.1], Hypo-CMV [1] and Hypo-CMV [10]] was collected and stored at −70°C until further use.
Cell proliferation and viability assay
To confirm the effect of VNR on MRC-5 proliferation, we performed the proliferation assay using the Cell Counting Kit-8 (CCK-8) assay (Dojindo, Kumamoto, Japan). In brief, MRC-5 cells were seeded at 2000 cells per well in a 96-well plate (Corning Costar Corp., Corning, NY, USA) and allowed to adhere overnight. Subsequently, the cells were replaced with media containing different concentrations of VNR. The control cells were treated similarly except the media in the absence of VNR. After 12-, 24- or 48-h incubation, 10 μL/well of CCK-8 solution (Dojindo) was added to each well and incubated for 1.5 h at 37°C. Absorbance at 450 nm was measured by a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA). Cell viability was assessed using trypan blue staining as described previously. 19
To evaluate the influence of the MRC-5 cell CM on HUVEC and 95D proliferations, HUVECs or 95Ds were grown in 96-well plates for 24 h and then the medium was replaced with the CM from the MRC-5 cells. After 12, 24 or 48 h, the absorbance was measured. The cell proliferation under each condition was expressed as a percentage of the control, which was set as 100%.
Cell migration assay
The chemotactic motility of cells was performed by using a transwell chamber (24-well chamber) with polycarbonate membranes (8.0 μm pore size), as described previously. 20 Briefly, HUVECs (3 × 104 cells per well) or 95Ds (8 × 104 cells per well) were re-suspended in warmed fresh medium containing 1% FBS and seeded into the upper chamber. To block the function of SDF-1/CXCR4, the cells were incubated with the CXCR4 inhibitor AMD3100 (10 μmol/L, Sigma) for 30 min before being seeded into the upper chamber. 21 The transwell chambers were then placed into 24-well plates, into which we added 600 μL CM. For exogenously increasing the content of SDF-1 in Hypo-CMV, recombinant human SDF-1 (100 ng/mL; PeproTech Inc., Rocky Hill, NJ, USA) was included in the Hypo-CMV in the lower chamber. 21 After incubating HUVECs for 6 h or 95Ds for 8 h, the media from the wells were withdrawn. The non-migratory cells on the upper membrane were gently wiped with cotton swabs, and the invading cells were fixed with 4% paraformaldehyde and stained with gentian violet. The number of stained cells in five randomly selected fields was counted.
Tube formation assay
The tube formation assay was assessed as described previously. 22 For reconstitution of a basement membrane, a 96-well plate was coated with Matrigel (50 μ/well) at 4°C and allowed to solidify (37°C, 30 min), prior to adding HUVECs (1.5 × 104 cells/well). To block the function of SDF-1/CXCR4, the cells were pretreated with 10 μmol/L AMD3100 for 30 min before being seeded onto the basement membrane. Subsequently, for exogenously increasing the content of SDF-1 in Hypo-CMV, 100 ng/mL recombinant human SDF-1 was premixed into the Hypo-CMV before the CM was added. The cells were incubated for six hours to allow formation of capillary-like structures in M199 with 20% FBS plus 10% CM. The changes in cell morphology were then captured by an inverted phase-contrast microscope (IX 70; Olympus, Tokyo, Japan). The tube numbers were quantified from five random fields.
Aortic ring assay
The rat aortic ring assay was performed as described previously with some modifications. 23 The thoracic aorta was dissected from male Sprague–Dawley rats (six weeks old) and cut into 1 mm diameter rings. Clotting media contained M199+ (M199 with 100 U/mL penicillin and 100 μg/mL streptomycin) plus 0.3% fibrinogen and 0.5% amino-n-caproic acid (ACA; Sigma). The growth media consisted of M199+ with 20% FBS and 0.5% ACA. Then the growth media mixed with CM (at a rate of 3:1) was added to the wells. The thoracic aortas were incubated with 10 μmol/L AMD3100 during the assay to block the function of SDF-1/CXCR4. For exogenously increasing the content of SDF-1 in Hypo-CMV, 100 ng/mL recombinant human SDF-1 was premixed into the Hypo-CMV. The plates were then cultured in an incubator at 37°C and 5% CO2. After seven days, the sprouting microvessels in five randomly chosen fields were photographed and counted under a microscope for each group.
The use of animals was in accordance with the guidelines established by the National Science Council of Republic China, with adherence to the ethical guidelines for the care and use of animals.
Enzyme-linked immunosorbent assay
The CM was collected as above. The assays were measured using humane SDF-1 and vascular endothelial growth factor (VEGF) kits (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions. Absorbance was measured using a spectrophotometer (Molecular Device Co., Sunnyvale, CA, USA). The standard curves and levels were obtained using the program Softmax™ (Molecular Device Co.).
Western blot analysis
Cells lysates were prepared and Western blot analyses were performed as previously described. 24 Briefly, HUVECs or 95Ds were cultured in CM for six or eight hours, and then the cell lysates were collected. Total protein was quantified with the bicinchoninic acid protein assay reagent kit (Pierce, Rockford, IL, USA). Equal amounts of protein were separated on sodium dodecyl sulfate polyacrylamide gel electrophoresis gels and transferred onto nitrocellulose membranes, and incubated (overnight, 4°C) with primary antibodies followed by horseradish peroxidase-conjugated secondary antibodies. Detection was performed by using an ECL System (Amersham Pharmacia Biotech, Piscataway, NJ, USA). Primary antibodies for Akt (1:1000), p-Akt (1:500), ERK1/2 (1:1000) and p-ERK1/2 (1:1000) were obtained from Cell Signaling Technology (Beverly, MA, USA). Other primary antibodies included antibodies against HIF-1α (1:500; R&D Systems), α-SMA (1:800; Santa Cruz Biotechnology, Santa Cruz, CA, USA), CXCR4 (1:400; Santa Cruz Biotechnology) and β-actin (1:5000; Sigma). Horseradish peroxidase-linked anti-mouse IgG (Sigma) and anti-rabbit IgG (Cell Signaling Technology) were used as secondary antibodies. Immunoreactive proteins on the membrane were visualized by enhanced chemiluminescence Western blotting detection reagents (Amersham Pharmacia Biotech).
Statistical analysis
Quantitative data are expressed as mean ± SD. Comparisons were analyzed by the Student's t-test. All present data represent the results of at least three independent experiments.
Results
α-SMA and HIF-1α were up-regulated in MRC-5 under hypoxia
Activation of the α-SMA gene during the conversion of fibroblasts into myofibroblasts is an essential feature. Therefore, to characterize the shift of cultured fibroblasts, we investigated the level of α-SMA protein in hypoxic MRC-5 cells. As shown in Figure 1, the expression of α-SMA increased significantly under hypoxia for 12 h, suggesting that hypoxic MRC-5 are analogous to CAFs to some extent. Meanwhile, HIF-1α, confirmed to be the major transcriptional factor in response to hypoxia, was also up-regulated, which also demonstrated the response of MRC-5 to hypoxia (Figure 1).
Hypoxia up-regulated α-SMA and HIF-1α protein expression in MRC-5. MRC-5 cells were cultured under normoxia or hypoxia for 12 h, and the cells lysates were collected for Western blot assay. HIF-1α, hypoxia-inducible factor 1α; α-SMA, α-smooth muscle actin
Effect of VNR on MRC-5 proliferation
To evaluate the effect of VNR on the proliferation of MRC-5 cells, cell proliferation was assessed by CCK-8 assay with logarithmically growing cells. The MRC-5 cells were cultured in the absence or presence of various concentrations of VNR for 12, 24 or 48 h. As shown in Figure 2a, 0.1, 1 and 10 μmol/L VNR treatment, respectively, for 12 h had only a marginal inhibition to cell proliferation, but had no cytotoxicity to the cells as detected by trypan blue assay (Figure 2b). The time points which did not cause cytotoxicity were then employed in subsequent studies.
Effects of VNR and its CM on cell proliferation. (a) Effect of VNR on proliferation of MRC-5 cells. MRC-5 cells were incubated overnight and then exposed to different concentrations of VNR (0.1, 1.0 and 10 μmol/L) and cultured for 12, 24 and 48 h. Cell proliferation was determined by CCK-8 assay. (b) Effect of VNR on the viability of MRC-5 cells. Cells were treated with VNR (0.1, 1.0 and 10 μmol/L) and incubated for 24 h. Cell viability was measured by trypan blue assay. (c) Effect of CM from hypoxic MRC-5 cells pretreated by VNR on HUVEC proliferation. Hypoxic MRC-5 was pretreated with different concentrations of VNR (0.1, 1.0 and 10 μmol/L). The CM was then collected and added to HUVECs 12, 24 or 48 h later. HUVEC proliferation was determined by CCK-8 assay. (d) Effect of CM from hypoxic MRC-5 cells pretreated by VNR on 95D proliferation. Hypoxic MRC-5 was pretreated with different concentrations of VNR (0.1, 1.0 and 10 μmol/L). The CM was then collected and added to 95Ds. 12, 24 or 48 h later, cell proliferation was determined by CCK-8 assay. VNR, vinorelbine; CM, conditioned medium; HUVEC, human umbilical vein endothelial cell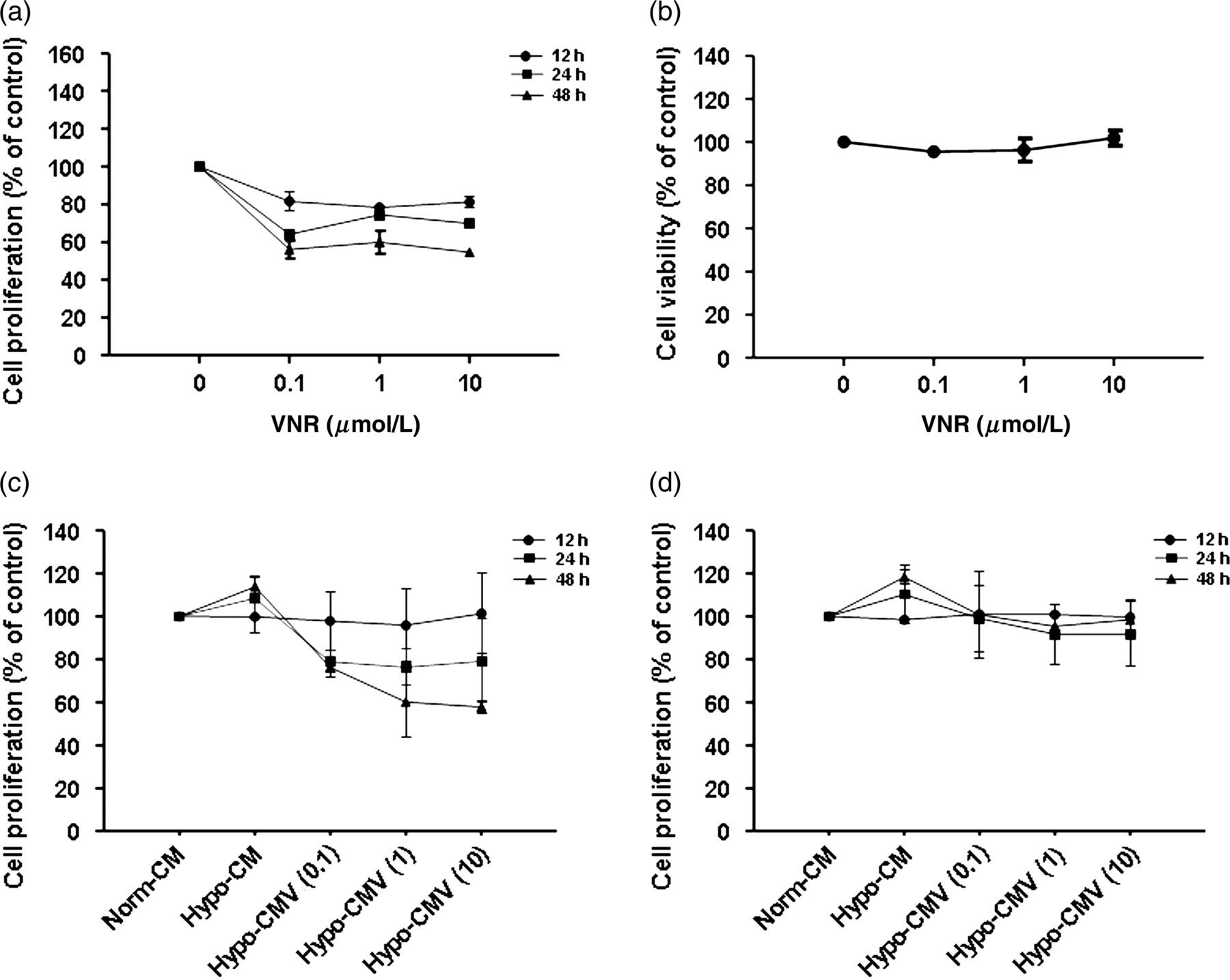
Effect of hypoxic MRC-5 CM triggered by VNR on HUVEC and 95D proliferation
Angiogenesis is a complex process that includes the proliferation, migration and tube formation of endothelial cells. To analyze the effect of hypoxic MRC-5 CM trigged by VNR (Hypo-CMV) on proliferation, HUVECs were plated at a certain density and volume, followed by treatment with the CM from hypoxic MRC-5 cells in the absence or presence of VNR within the CCK-8 assay. As shown in Figure 2c, the stimulation of the CM from hypoxic MRC-5 without VNR treatment had no significant effect on HUVEC proliferation compared with Norm-CM at 12 h. Likewise, the CM from hypoxic MRC-5 cells pretreated with VNR did not alter HUVEC proliferation for 12 h either, when we chose to detect the effects of the CM on the migration and angiogenesis assay later, to avoid possible effects of the CM on cell growth.
Besides angiogenesis, tumor growth and metastasis also rely on the proliferation and migration of cancer cells themselves; therefore, we also studied the effect of fibroblasts on 95D cells. To address this issue, we observed the effect of Hypo-CM with or without pretreatment by VNR on 95D proliferation. As shown in Figure 2d, Hypo-CM had scarcely any influence on 95D proliferation. Hypo-CMV did not significantly impact 95D proliferation at each time point either.
Hypoxic MRC-5 CM triggered by VNR inhibited HUVEC and 95D migration
As cell migration is a prerequisite for tumor, we used the transwell chamber assay to examine if VNR could affect HUVEC and 95D migration induced by Hypo-CM. After stimulating HUVECs or 95Ds with Hypo-CM for six or eight hours, respectively, a greater number of cells moved to the lower side of the membrane in the transwell chamber compared with Norm-CM (Figures 3a and c), indicating that MRC-5 under hypoxia was able to promote HUVEC and 95D migration. However, pretreatment of the MRC-5 cells with 0.1, 1 and 10 μmol/L VNR, respectively, for 12 h dramatically reduced the HUVEC and 95D migration induced by the Hypo-CM. The inhibitory rates (Figure 3b) for HUVECs were 36.09%, 52.98% and 42.18%, respectively, and the inhibitory rates (Figure 3d) for 95Ds reached 39.84%, 74.81% and 64.24% respectively.
Effect of CM from hypoxic MRC-5 pretreated by VNR on HUVEC and 95D migration. (a) Photomicrographs of HUVEC migration to the lower side of chamber. Cells were pretreated with Norm-CM, Hypo-CM, Hypo-CMV (0.1), Hypo-CMV (1), Hypo-CMV (10), respectively, before being transferred into the up side of chamber, and migration for six hours. (b) Bar graph shows results of quantitative analysis of HUVEC migration. Values are reported as mean ± SD from three independent experiments. #
P < 0.05 compared with Norm-CM, *P < 0.05, **P < 0.01 compared with Hypo-CM. (c) Photomicrographs of 95D migration to the lower side of chamber. Cells were pretreated with Norm-CM, Hypo-CM, Hypo-CMV (0.1), Hypo-CMV (1), Hypo-CMV (10), respectively, before being transferred into the up side of chamber, and migration for eight hours. (d) Bar graph shows results of quantitative analysis of 95D migration. Values are reported as mean ± SD from three independent experiments. #
P < 0.05 compared with Norm-CM, **P < 0.01 compared with Hypo-CM. VNR, vinorelbine; CM, conditioned medium; HUVEC, human umbilical vein endothelial cell
Hypoxic MRC-5 CM trigged by VNR inhibited HUVEC tube formation
As organization of endothelial cells into a network of tubes is a late event during angiogenesis, we next introduced the Matrigel-induced tube formation assay to determine whether Hypo-CMV impacts HUVEC tube formation. The results showed that the Hypo-CM induced much more tube-like structure formation in HUVECs compared with the Norm-CM group (Figure 4a). In comparison to the Hypo-CM group, the tube structures were disrupted when HUVECs were exposed to the CM, which was collected from VNR-pretreated hypoxic MRC-5 cells. HUVECs treated with Hypo-CMV (0.1) or Hypo-CMV (1) differentiated into short tubes but were unable to form meshes, whereas those treated with Hypo-CMV (10) remained dotted on the matrigel. HUVECs incubated in Hypo-CMV (0.1, 1 and 10) formed much less tube-like structures. The number of tube-like structures versus Norm-CM were 126 ± 3% (Hypo-CM), 83 ± 5% (Hypo-CMV [0.1]), 3 ± 0.4% (MV [1]) and 2 ± 0.1% (Hypo-CMV [10]) (Figure 4b). These observations revealed that Hypoxic MRC-5 CM trigged by VNR suppressed HUVEC tube formation.
Effects of CM from hypoxic MRC-5 pretreated by VNR on HUVEC tube formation and rat aortic ring microvessel sprouting. (a) Photomicrographs of HUVEC tube formation. Cells were pretreated with Norm-CM, Hypo-CM, Hypo-CMV (0.1), Hypo-CMV (1), Hypo-CMV (10), respectively, before being used to perform tube formation experiments. (b) Bar graph shows results of quantitative analysis of HUVEC tube formation. Values are reported as the mean ± SD from three independent experiments. #
P < 0.05 compared with Norm-CM, *P < 0.05, ***P < 0.001 compared with Hypo-CM. (c) Photomicrographs of rat aortic ring microvessel sprouting. Cells were pretreated with Norm-CM, Hypo-CM, Hypo-CMV (0.1), Hypo-CMV (1), Hypo-CMV (10), respectively, before being used to determinate aortic ring microvessel sprouting. (d) Bar graph shows results of quantitative analysis of aortic ring microvessel sprouting. Values are reported as the mean ± SD from three independent experiments. ##
P < 0.01 compared with Norm-CM, **P < 0.01 compared with Hypo-CM. VNR, vinorelbine; CM, conditioned medium; HUVEC, human umbilical vein endothelial cell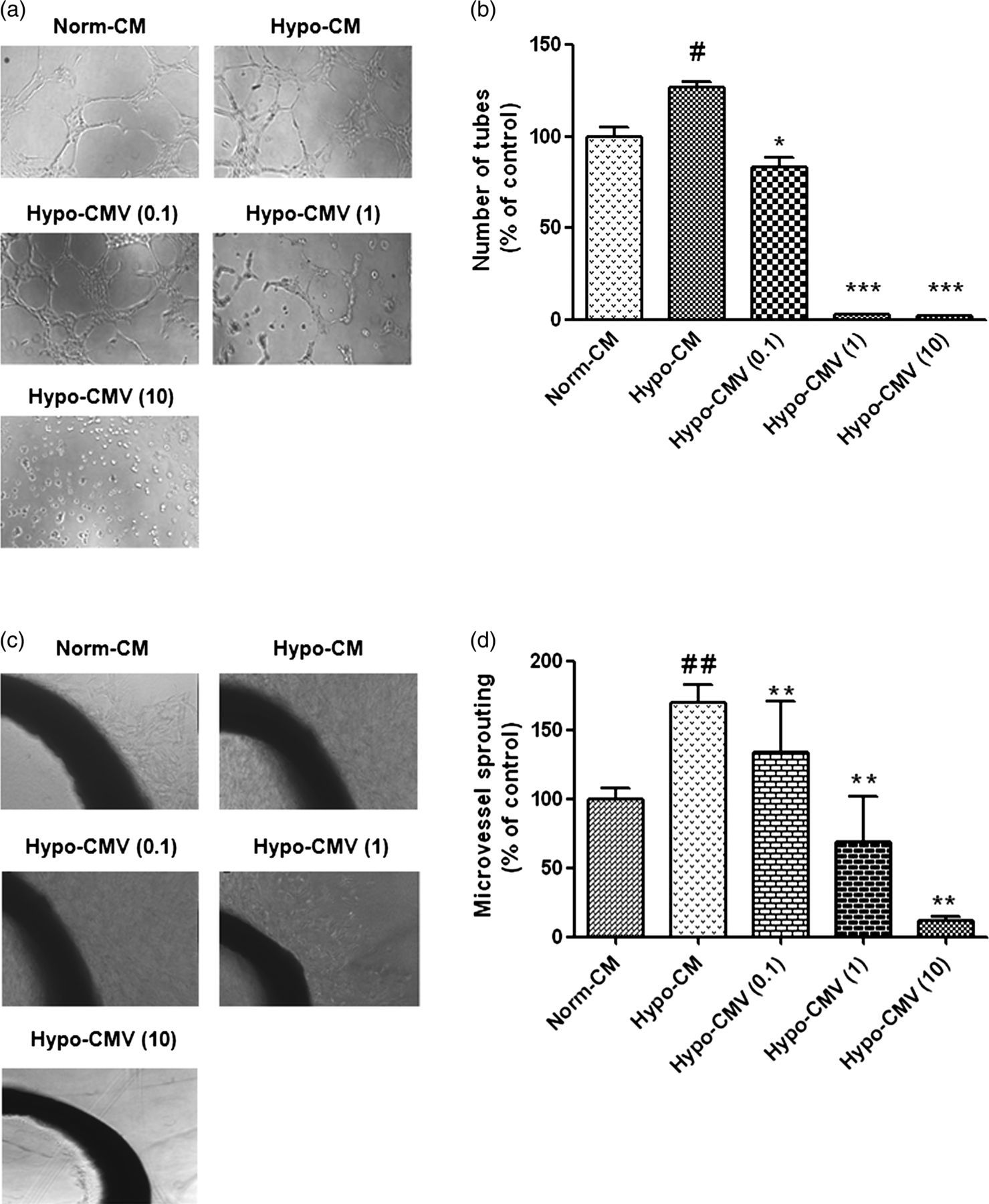
Hypoxic MRC-5 CM trigged by VNR inhibited angiogenesis ex vivo
Rat aortic ring assay includes several necessary steps involved in neovascularization, such as endothelial cell proliferation, migration and tube formation; therefore, it is considered to mimic in vivo angiogenesis closely. We used this model to further explore the anti-angiogenic effects of Hypo-CMV. The results (Figure 4c) showed that the Hypo-CMV facilitated the new microvessel growth compared with the Norm-CM group, with the growing sprouts being longer and more. When treated with Hypo-CMV (10), the microvessel was almost completely inhibited. These results correspond to those found in the tube formation assay, indicating that VNR obtained anti-angiogenic effects indirectly through its action on fibroblasts.
VNR impacted hypoxia-induced α-SMA protein expression in MRC-5
In the above study, we found that hypoxic MRC-5 converted to an activation phenotype, and promoted angiogenesis and 95D migration, which were all reversed by VNR. To reveal the probability mechanism of VNR action, we decided to investigate the effect of VNR on α-SMA protein expression in hypoxic MRC-5 cells firstly. The level of α-SMA protein was evaluated by Western blot analysis in MRC-5 cells, which were pretreated by VNR for 12 h and then exposed to hypoxia for 12 h. As shown in Figure 5a, pretreatment with VNR obviously attenuated hypoxic up-regulation of α-SMA.
VNR impacted hypoxia-induced α-SMA protein and HIF-1α expression and growth factors secreted by MRC-5 cells. (a) VNR inhibited α-SMA protein and HIF-1α expression in hypoxic MRC-5 cells. The MRC-5 cells were pretreated with indicated concentrations of VNR for 12 h, followed by culturing in hypoxia for 12 h, and then lysed for detecting α-SMA and HIF-1α expression by Western blot. (b) SDF-1 amounts in the media were measured by ELISA. The MRC-5 cells were pretreated with indicated concentrations of VNR for 12 h, followed by culturing in hypoxia for 12 h, and the cell supernatant was obtained for detecting SDF-1 secretion. Data are representative for the mean ± SD obtained from three independent experiments. #
P < 0.05 compared with Norm-CM, *P < 0.05 compared with Hypo-CM. (c) VEGF amounts in the media were measured by ELISA. The MRC-5 cells were pretreated with indicated concentrations of VNR for 12 h, and then cultured in hypoxia for 12 h. The cell supernatant was obtained for detecting VEGF secretion. Data are representative of the mean ± SD obtained from three independent experiments. ###
P < 0.001 compared with Norm-CM. VNR, vinorelbine; CM, conditioned medium; HUVEC, human umbilical vein endothelial cell; α-SMA, α-smooth muscle actin; HIF-1α, hypoxia-inducible factor 1α; SDF-1, stromal cell-derived factor 1; ELISA, enzyme-linked immunosorbent assay; VEGF, vascular endothelial growth factor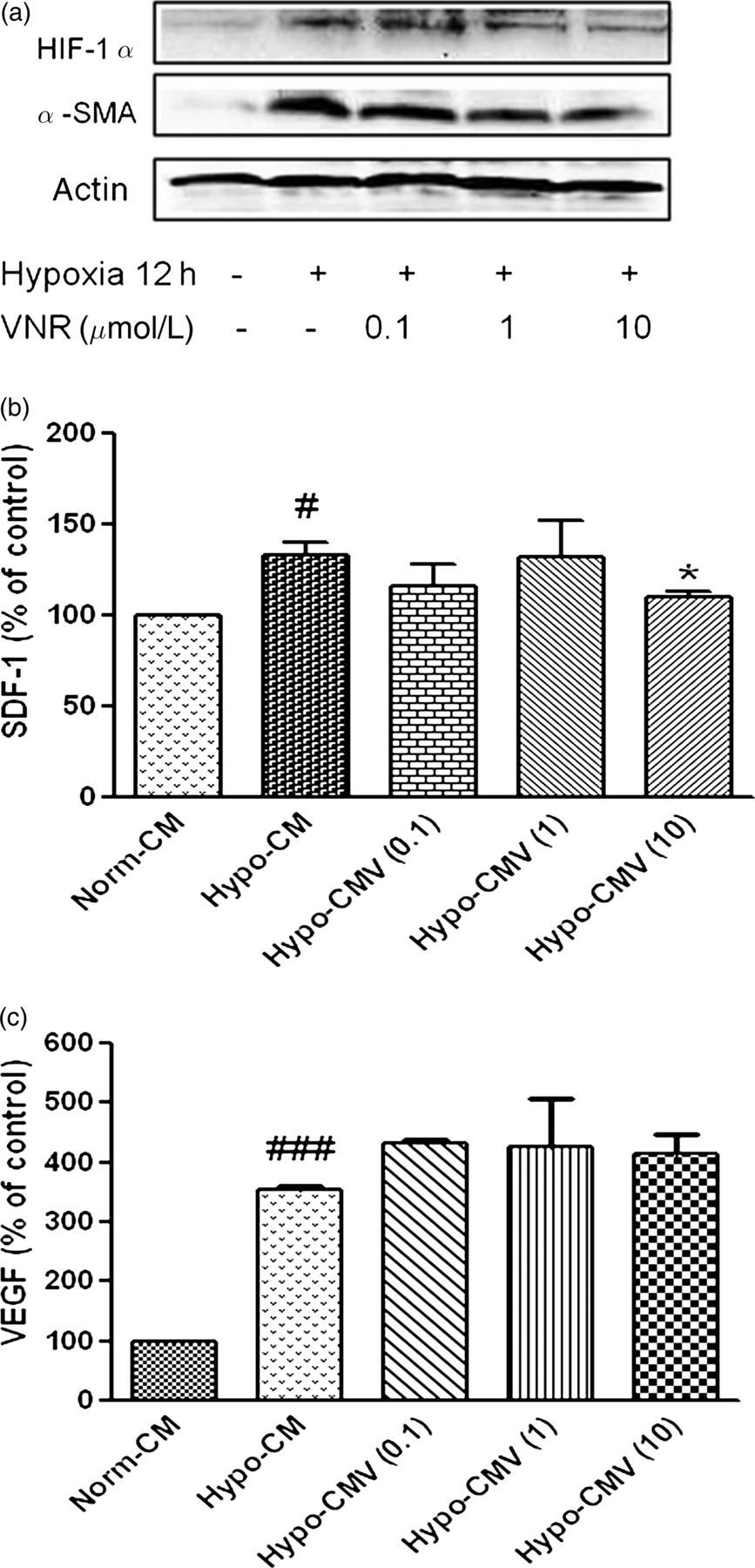
VNR impacted hypoxia-induced HIF-1α expression and growth factors secreted by MRC-5 cells
SDF-1 and VEGF, which could be regulated by HIF-1α under the hypoxic condition, play a central role in the promotion of tumor invasion and angiogenesis. To study the potential mechanisms of CM leading to mediation of the inhibition effects of VNR, we first detected if VNR was able to influence the expression of HIF-1α in hypoxic MRC-5. The results in Figure 5a showed that VNR down-regulated HIF-1α expression under hypoxia. We then used enzyme-linked immunosorbent assay to explore if the inhibition of VNR on HIF-1α further affected the release of SDF-1 and VEGF by hypoxic MRC-5 cells. As shown in Figures 5b and c, SDF-1 and VEGF production by hypoxic MRC-5 cells increased significantly at 12 h. Pretreatment with 10 μmol/L VNR for 12 h inhibited hypoxic induction of SDF-1 protein. However, VEGF secretion had no alteration in hypoxic MRC-5 CM pretreated by VNR.
SDF-1/CXCR4 axes participated in the effect of hypo-CMV on angiogenesis and cell migration
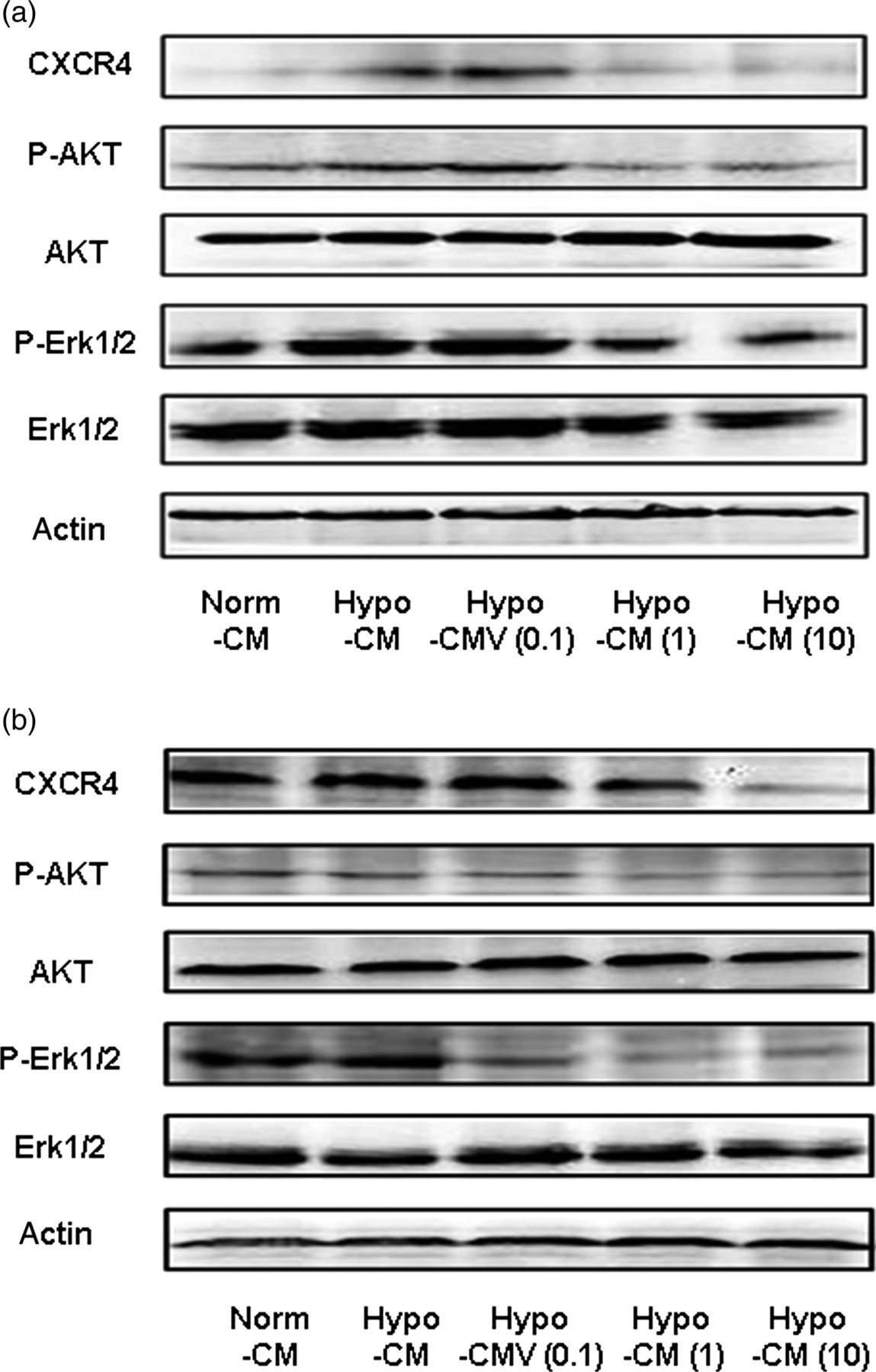
As we have shown that VNR inhibited SDF-1 released from MRC-5 cells under hypoxia, we supposed that the SDF-1/CXCR4 axis may contribute to the observed effects of VNR in our experiments. To provide direct evidence that the SDF-1/CXCR4 signal accounts for the effects of Hypo-CM and Hypo-CMV on 95D migration and angiogenesis, we additionally introduced a CXCR4 antagonist AMD3100 or recombinant human SDF-1. We found that pro-migration and pro-angiogenesis induced by Hypo-CM were significantly reversed by ADM3100 pretreatment; meanwhile, by adding exogenous SDF-1 to the Hypo-CMV, we were able to attenuate the inhibition effects of Hypo-CMV on HUVEC and 95D migration (Figures 6a and b) and angiogenesis (Figures 6c and d). We also performed Western blot analysis to evaluate the change in the SDF-1/CXCR4 signaling pathway in HUVECs and 95Ds after Hypo-CMV treatment. We first detected the expression of SDF-1 receptor (CXCR4) on HUVECs and 95Ds, and observed that Hypo-CMV treatment greatly decreased this receptor expression (Figures 7a and b). The downstream of CXCR4 is mainly related to pathways concerning the Akt and ERK signal. Therefore, we took further steps to investigate the alternation of those signals. The results in Figures 7a and b show the down-regulation of the expression of P-Akt and P-ERK1/2. In all cases, the total steady state protein levels of Akt and ERK1/2 remained unchanged. Taken together, these results implied that the SDF-1/CXCR4 axis mediated VNR inhibition on angiogenesis and 95D migration through its effects on MRC-5.
SDF-1/CXCR4 mediated the inhibition effects of CM from hypoxic MRC-5 pretreated with VNR. (a) SDF-1/CXCR4 mediated the inhibition effects of Hypo-CMV on HUVEC migration. HUVECs were pretreated with or without AMD3100 before being added into the upper chamber. SDF-1 was premixed into the hypo-CMV to increase the content of SDF-1 exogenously as mentioned in the corresponding Methods and Results sections. Bar graph shows results of quantitative analysis of HUVEC migration. Values are reported as mean ± SD from three independent experiments (*P < 0.05). (b) SDF-1/CXCR4 mediated the inhibition effects of Hypo-CMV on 95D migration. Cells were pretreated with or without AMD3100 before being added into the upper chamber. SDF-1 was premixed into the hypo-CMV to increase the content of SDF-1 exogenously as mentioned in the corresponding Methods and Results sections. Bar graph shows results of quantitative analysis of 95D migration. Values are reported as mean ± SD from three independent experiments (*P < 0.05, **P < 0.01). (c) SDF-1/CXCR4 mediated the inhibition effects of Hypo-CMV on tube formation. Cells were pretreated with or without AMD3100 before being added onto basement membrane to form tubes. SDF-1 was premixed into the hypo-CMV to increase the content of SDF-1 exogenously as mentioned in the corresponding Methods and Results sections. Bar graph shows results of quantitative analysis of tube formation. Values are reported as mean ± SD from three independent experiments (*P < 0.05, **P < 0.01). (d) SDF-1/CXCR4 mediated the inhibition effects of Hypo-CMV on rat aortic ring microvessel sprouting; aortic rings were treated with or without AMD3100 during the assay. SDF-1 was premixed into the hypo-CMV to increase the content of SDF-1 exogenously as mentioned in the corresponding Methods and Results sections. Bar graph shows results of quantitative analysis of rat aortic ring microvessel sprouting. Values are reported as mean ± SD from three independent experiments (*P < 0.05, **P < 0.01). VNR, vinorelbine; CM, conditioned medium; HUVEC, human umbilical vein endothelial cell; SDF-1, stromal cell-derived factor 1 Effects of hypoxic MRC-5 CM trigged by VNR on the activities of SDF-1/CXCR4 related signaling pathways. (a) Effects of hypoxic MRC-5 CM trigged by VNR on SDF-1/CXCR4-related signaling pathways in HUVECs. After each CM treatment for six hours, cell lysates were utilized to detect the CXCR4, and the phosphorylated protein levels of Akt and ERK1/2 in HUVECs by Western blot. (b) Effects of hypoxic MRC-5 CM trigged by VNR on SDF-1/CXCR4-related signaling pathways in 95Ds. Western blot assays were used to examine CXCR4, and the phosphorylated protein levels of Akt and ERK1/2 after different CM treatments for eight hours. VNR, vinorelbine; CM, conditioned medium; HUVEC, human umbilical vein endothelial cell; SDF-1, stromal cell-derived factor 1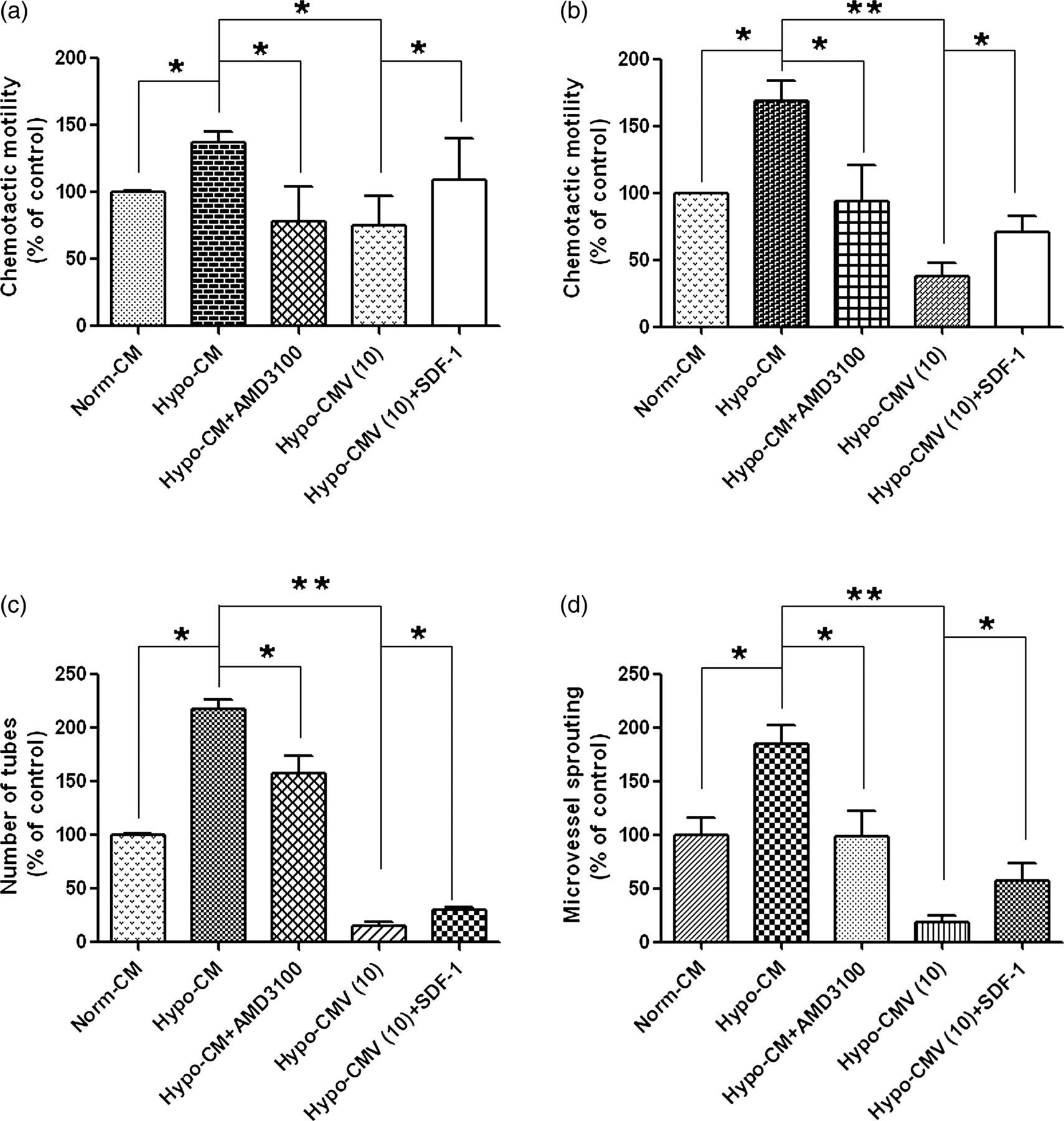
Discussion
Current anticancer therapy has received great achievement, mainly aimed at inducing tumor cell differentiation and apoptosis, inhibiting tumor cell proliferation and migration. As well as the above, drug research has already expanded to other aspects, including drug resistance, tumor angiogenesis, immune destruction, inflammation and so on. With the general investigation for tumor development, the roles of tumor stroma have been greatly proved and emphasized. Reliance of tumor on stroma tightly refers to the entire process of carcinogenesis, growth, and metastasis. 25 Thus, therapy against stroma has gradually been considered as a way to control cancer. 26,27 Although the interaction between stroma and tumor, as well as the associated mechanism, has been exposed in recent years, little is known about the interruption to stroma–tumor interaction under drug treatment. 28–30 Our present study revealed a novel role of VNR, a widely used anticancer drug, in tumor angiogenesis and migration regulated by stroma fibroblasts. This finding underlies the importance of drug treatment aiming at tumor stroma.
The importance of the tumor stroma in tumor progression has been recognized for many years and has been highlighted in several reviews. 8,31,32 Multiple cell types constitute tumor stroma, among which fibroblasts are considered to be one of the most important members. As we all know, normal fibroblasts can be transformed into CAFs, and CAFs are involved crucially in tumor development via secretion of extracellular matrix components, growth factors and proteases, all of which facilitate tumor angiogenesis and metastasis. 33,34 Previous studies have demonstrated that hypoxia can activate fibroblasts, inducing their expression of α-SMA, which is a marker of myofibroblasts. In our present study, we cultured MRC-5 cells under hypoxia for 12 h and confirmed the increase in the expression of α-SMA in MRC-5 cells, suggesting that hypoxic MRC-5 cells have a myofibroblastic phenotype. This allowed us to use hypoxic MRC-5 cells to mimic CAFs to do further research.
Tumor angiogenesis, a complex multistep process involving extracellular matrix remodeling, endothelial cell migration and proliferation and capillary tube formation, is essential for tumor growth and metastasis. 35 To clarify the influence of fibroblasts on tumor angiogenesis and migration, we adopted the CM from hypoxic MRC-5 cells, analogous to CAFs, to explore the potency of CM on HUVEC proliferation, tube formation and migration, as well as aorta sprout outgrowth. Our results showed that Hypo-CM significantly enhanced the above processes except the proliferation of HUVECs at 12 h, compared with Norm-CM. In line with our results, previous work also provides strong support to the idea that the fibroblasts, involved in invasion and angiogenesis, are extremely crucial in tumor microenvironment. For instance, the group of Charlotta J Olsen and José Moreira demonstrated that co-culturing of human tumor cells and fibroblasts in 3D Matrigel in the absence of direct cell contacts stimulated the production of a complex network of molecules that induce invasion of both cell types, generating a more aggressive phenotype. 31 Therefore, considering the critical role of fibroblasts on tumor angiogenesis and invasion, this field is gaining more and more attention. 2,36
Despite the increasing appreciation of the importance of fibroblasts in progression of cancer, little is known about the factors regulating the interaction between tumor and stroma. Hence, to explore the mechanisms of how the Hypo-CM affects angiogenesis and 95D migration, we chose cytokine VEGF and SDF-1 for further study. VEGF, one of the key mediators of angiogenesis, was described to regulate most of the steps in angiogenic cascades. 37 In our experiments, the VEGF released by MRC-5 was up-regulated under hypoxia. We also inspected the possible involvement of the SDF-1 signal. Previous reports have shown that the SDF-1/CXCR4 axis plays a central role in the promotion of tumor growth, angiogenesis and metastasis. 9,38,39 Recently, there has been increasing evidence that SDF-1 enhanced pancreatic cancer cell invasiveness and promoted HUVEC tube formation in vitro through binding to CXCR4 in a paracrine manner. 40,41 Under a hypoxic condition, it has been shown to not only up-regulate total CXCR4 but also cell surface CXCR4 expression in various cancer cells 11,12,42–45 and stromal cells 11,46,47 via HIF-1α, which has also been demonstrated to enhance SDF-1 secretion. 48–51 This improved activity of the SDF-1/CXCR4 signal under hypoxia has been widely confirmed to contribute to the metastatic potential of tumors. The observation in our study was that SDF-1 produced by hypoxic MRC-5 cells was up-regulated compared with the normoxic MRC-5, which was in accordance with the increased expression of HIF-1α in hypoxic MRC-5 cells. Meanwhile, we also showed that CXCR4 expression was enhanced on HUVECs and 95Ds that were treated with the CM from hypoxic MRC-5 cells. Collectively, these data imply a potential mediation of SDF-1 and VEGF in the influence of hypoxic fibroblasts on HUVECs and 95Ds.
To date, considering the tolerability and proven efficacy, previous clinical reports have highlighted that VNR remains a vital agent in the treatment of non-small cell lung cancer. Deep investigations have already observed that VNR produced apparent antiangiogenic roles by direct cytotoxicity to endothelial cells, while little is known about the effect of VNR on the fibroblasts, which alsoregulated angiogenesis. Meanwhile, conventional anticancer drug screening is typically performed in the absence of fibroblasts of the tumor microenvironment, which might profoundly influence antitumor drug activity. 52,53 In the present study, we first observed that pretreatment of MRC-5 with VNR dramatically attenuated Hypo-CM-induced endothelial migration and tube formation. Furthermore, CM from hypoxic MRC-5 pretreated by VNR suppressed the microvessel sprouting from vascular tissues of rat aorta. This CM also inhibited the migration of 95Ds. Subsequently, we investigated the molecular mechanism by which VNR exerts its above effects. Unexpectedly, although the Hypo-CM promoted the secretion of VEGF, we did not observe any additional change after VNR pretreatment, indicating that VEGF is not involved in the above effects of VNR. However, the results showed that VNR inhibited hypoxic MRC-5 cell SDF-1 secretion, which then attenuated the capabilities of HUVECs in angiogenesis and 95D migration. VNR also decreased α-SMA expression by hypoxic MRC-5 cells, suggesting that VNR, to some extent, prevented the conversion of fibroblasts to myofibroblasts. More than this, the CXCR4 expression was down-regulated in HUVECs and 95Ds in response to Hypo-CMV. To prove that the SDF-1/CXCR4 signal is crucial for the observed effects, we blocked SDF-1/CXCR4 function with AMD3100, and identified that the promoting effects of hypoxic fibroblasts on cell migration and angiogenesis were mediated by the increased activity of the SDF-1/CXCR4 signal, while VNR strongly inhibited angiogenesis and cell migration induced by hypoxic fibroblasts, which are also SDF-1/CXCR4 signal-related, because adding exogenous SDF-1 to the Hypo-CMV was able to attenuate the inhibition effects of Hypo-CMV. Altogether these observations indicate that VNR could inhibit angiogenesis and cancer cell migration via interrupting fibroblast functions, and the mechanism responsible for the observed effects of VNR may partially be concerned with the SDF-1/CXCR4 axis. Furthermore, in order to investigate the specific effects of VNR on fibroblast under a non-toxicity condition, we adapted the time point of 12 h to collect the CM from MRC-5. However, it is also worth noting that 10 μmol/L VNR treatment of MRC-5 for 48 h exhibited an observed inhibition on MRC-5 proliferation, which implied another possible way of VNR to exert anticancer activity through directly depressing the growth of CAFs. The comprehensive mechanisms of VNR on angiogenesis and cancer cell migration through its action on stroma need further research.
Although the importance of fibroblasts in tumor development is now supported by clinical studies and other experimental models, the mechanism of tumor–stroma interaction is still not fully understood. 26 Our current data provides supplemental evidence that activated fibroblasts affect tumor angiogenesis and cancer cell migration. Moreover, based on the above, we demonstrate that VNR could affect the function of stromal fibroblasts, and alter the tumor stroma–tumor interaction by reducing the activation of the SDF-1/CXCR4 signal pathway. In this direction, the effects of anticancer drugs on fibroblasts should be taken into consideration, and advance in fibroblast research is proposed to initiate novel targets in anti-cancer therapy. 54
Footnotes
ACKNOWLEDGEMENTS
This work was financially supported by the Standardized Platform Construction and Scientific Application in New Technologies for New Drug Screening (no. 2009ZX09302-002), the Study of Saponin Monomer of Dwarf Lilyturf Tuber (DT-13): A New Natural Anti-metastatic Drug Candidate (no. 2009ZX09103-308) and the Research on Anti-tumor Metastasis Effect of DT-13 (no. 2011ZX8110285).