
Abstract
Neurotrophic factors such as glial cell line-derived neurotrophic factor (GDNF) and brain-derived neurotrophic factor (BDNF) are considered to contribute to the development, maintenance and survival of neurons, glia and oligodendrocytes. Astroglia are a major source of various neurotrophic factors. Thus, enhancement of astroglia-mediated neurotrophic factor release might hold promising potential for neurological diseases. Resveratrol, a natural non-flavonoid polyphenol found in grapes and red wine, has been recognized to be beneficial for health. Here, rat primary astroglia-enriched cultures were used to investigate the effects of resveratrol-mediated neurotrophic factor release and the related mechanisms. The cultures were treated with 25–100 μmmol/L resveratrol for 12–48 h. Results showed resveratrol increased BDNF and GDNF production in the culture medium. In addition, the production of BDNF in the supernatant of cultures was increased five-fold over control cultures 24 h after resveratrol treatment and then remained high 36 h later. Meanwhile, the production of GDNF was initially increased by up to four-fold 24 h after resveratrol treatment and continued to increase to six-fold at 36 h and remained at a high level till 48 h. Western blot analysis of BDNF and GDNF protein in astroglia at different time points after resveratrol treatment indicated similar increases. Furthermore, resveratrol significantly induced the phosphorylation of extracellular signal-regulated kinase 1/2 (ERK1/2) and cAMP responsive element-binding protein (CREB) in astroglia. Overall, resveratrol is effective in promoting astroglia-derived neurotrophic factor release, and this effect is mediated, at least in part, by the activation of ERK1/2 and CREB.
Introduction
Recent advances have demonstrated that glia (particularly astroglia and microglia) play an indispensable role in neurological disorders and have become the prime targets for therapy. 1,2 Astroglia serve many housekeeping functions, such as the maintenance of the extracellular environment and the stabilization of cell–cell communications in the central nervous system (CNS). 3 It is becoming increasingly evident that astroglia are a major source of various neurotrophic factors, such as glial cell line-derived neurotrophic factor (GDNF) and brain-derived neurotrophic factor (BDNF). 4 Furthermore, neurotrophic factors are considered to contribute to the development, maintenance and survival of neurons, glia and oligodendrocytes. 5 In contrast, lack of neurotrophic factors might lead to neuronal death and the pathogenesis of neurodegenerative diseases. 6,7 Thus, enhancement of astroglia-mediated neurotrophic factor release might hold promising potential for the treatment of neurological diseases.
Resveratrol (3, 4′, 5-trihydroxy-trans-stilbene), a natural non-flavonoid polyphenol found in grapes and red wine, has been recognized to be beneficial for health. 8 It is known to possess a large number of pharmacological properties such as cardioprotective, anti-inflammatory, anti-oxidant and anti-cancer effects. 9 Besides these beneficial actions, increasing interest has focused on its neuroprotective activities. Resveratrol could cross the blood–brain barrier 10 and produce neuroprotection against ischemia, seizure and neurodegenerative diseases. 11 Recent studies indicated that resveratrol improved hippocampal atrophy through enhancing neurogenesis and inhibiting apoptosis of granular cells in chronic fatigue mice. 12 However, the mechanisms underlying these effects still remain unclear. Our previous studies demonstrated that resveratrol protected dopaminergic neurons from lipopolysaccharide (LPS)-induced neurodegeneration via its anti-inflammatory properties. 13 To extend our understanding of resveratrol mediated neuroprotection, rat primary astroglia cultures were applied to investigate the effects of resveratrol on the production of neurotrophic factors from astroglia and further elucidate the related molecular mechanisms.
Materials and methods
Animals
Female Wistar rats (200–300 g) were purchased from the Experimental Animal Centre of the Third Military Medical University (Chongqing, China; Specific-pathogen free Grade II; Certificate No. scxk 2002003). Housing and breeding of the animals were performed in strict accordance with Animal Care and Use Guidelines in China.
Reagents
Resveratrol was obtained from Sigma Chemical Co. (St Louis, MO, USA). All the materials of cell cultures were purchased from Invitrogen (Carlsbad, CA, USA). Primary antibodies were obtained from Cell Signaling Technology (Beverly, MA, USA). The AntiVectastain avidin–biotin complex kit was obtained from Vector Laboratories (Burlingame, CA, USA). Enzyme-linked immunosorbent assay (ELISA) kits were obtained from Chemicon International Inc. (Temecula, CA, USA).
Rat primary astroglia-enriched cultures
Primary astroglia-enriched cultures were prepared from the whole brains of one-day-old rat pups as described previously. 14 After a confluent monolayer of glial cells had been established, microglia were separated from astroglia by shaking the flasks at 180 rpm for 1 h. The remaining astroglia were detached with trypsin-ethylenediaminetetraacetic acid (EDTA) and seeded in the culture medium. After two or three consecutive passages, immunocytochemical analysis indicated that the cultures were composed of >98% astroglia.
Measurement of BDNF and GDNF by ELISA
The production of BDNF and GDNF in the culture supernatant was quantified using ELISA kits according to the procedures provided by the manufacturer.
Western blot analysis
The whole cells from primary astroglia-enriched cultures were washed with cold phosphate-buffered saline and lysed with radioimmunoprecipitation assay (RIPA) cell lysis buffer consisting of 50 mmol/L Tris–HCl (pH 7.4), 150 mmol/L NaCl, 1 mmol/L EDTA, 5 μg/mL aprotinin, 1 mmol/L phenylmethylsulfonyl fluoride, 5 μg/mL leupeptin, 1% Triton X-100, 1% sodium deoxycholate and 1 mmol/L Na3VO4.
13
The lysates were incubated on ice for 30 min and then centrifuged at 12,000×
Statistical analysis
Data are expressed as mean ± SEM from three independent experiments performed in triplicate. Statistical significance was assessed by analysis of variance (ANOVA) using GraphPad Prism software (GraphPad Software Inc., San Diego, CA, USA). When ANOVA showed significant differences, pairwise comparisons between means were tested by Bonferroni's post-test with correction. A value of P < 0.05 was considered statistically significant.
Results
Resveratrol increased BDNF and GDNF production in the culture medium
Rat primary astroglia-enriched cultures were treated with resveratrol (25–100 μmol/L). After resveratrol treatment for 24 h, the release of BDNF and GDNF in the supernatant was detected by ELISA. On the basis of ANOVA with Bonferroni's test analysis, resveratrol significantly increased BDNF (six-fold) and GDNF (four-fold) production in the culture medium as shown in Figure 1.
Resveratrol increased brain-derived neurotrophic factor (BDNF) and glial cell line-derived neurotrophic factor (GDNF) production in the culture medium. The release of BDNF (a) and GDNF (b) in the culture medium 24 h after treatment with resveratrol was detected by enzyme-linked immunosorbent assay. Data are expressed as a percentage of the control cultures and are the mean ± SEM from three independent experiments performed in triplicate. *P < 0.05 compared with the control cultures (analysis of variance with Bonferroni's test). (A color version of this figure is available in the online journal)
Resveratrol time-dependently induced BDNF and GDNF release
Rat primary astroglia-enriched cultures were treated with resveratrol (100 μmol/L). Twelve, 24, 36 and 48 h after resveratrol treatment, the release of BDNF and GDNF in the supernatant was measured by ELISA. On the basis of ANOVA with Bonferroni's test analysis, the production of BDNF was increased by up to five-fold of control cultures 24 h after resveratrol treatment and then maintained at a four-fold level 36 h later (Figure 2a). The production of GDNF was initially increased by up to four-fold 24 h after resveratrol application and continued to increase to six-fold at 36 h and was still five-fold higher 48 h later (Figure 2b).
Resveratrol time-dependently induced brain-derived neurotrophic factor (BDNF) and glial cell line-derived neurotrophic factor (GDNF) release. The release of BDNF (a) and GDNF (b) in the supernatant after treatment of 100 μmol/L resveratrol was measured by enzyme-linked immunosorbent assay. Data are expressed as a percentage of the control cultures and are the mean ± SEM from three independent experiments performed in triplicate. *P < 0.05 compared with the control cultures (analysis of variance with Bonferroni's test)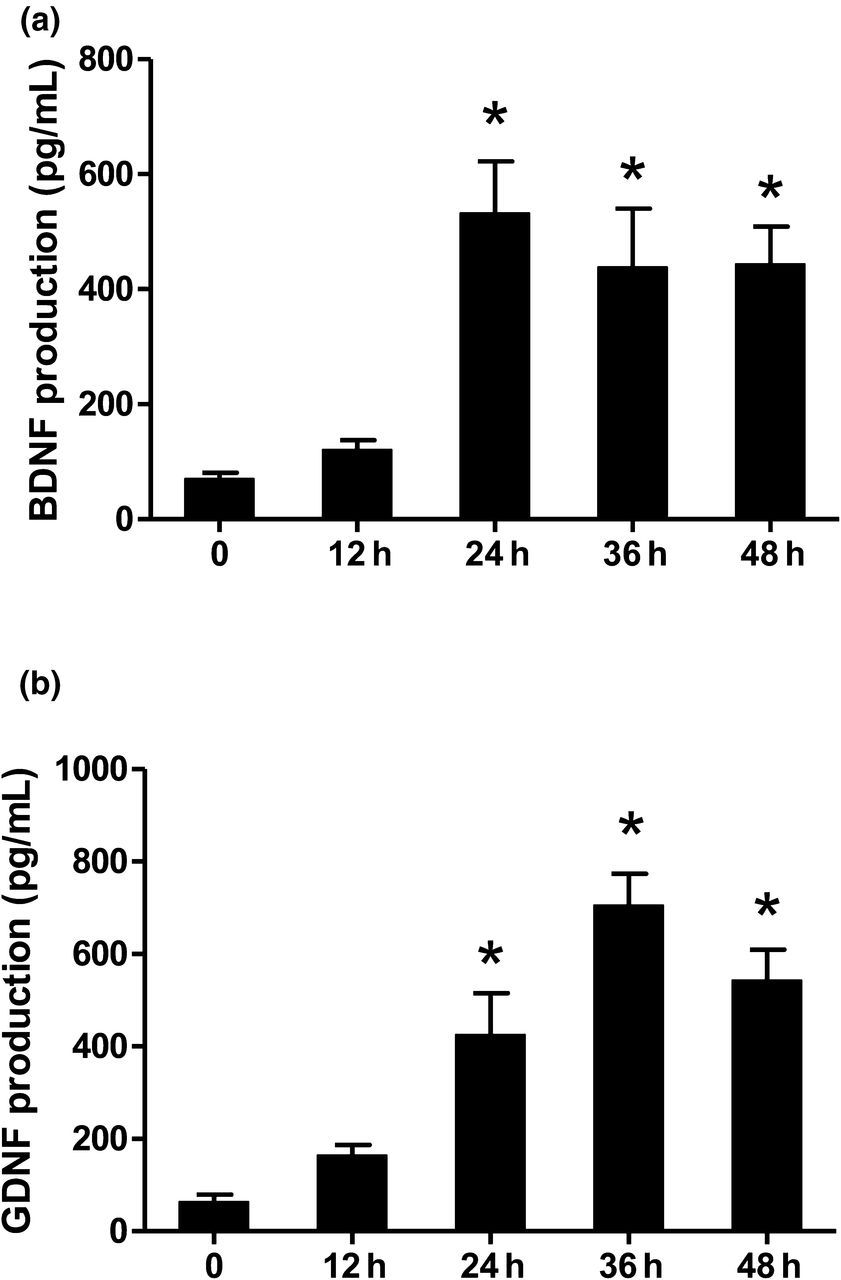
To further investigate the effects of resveratrol on BDNF and GDNF protein levels, total cell protein was harvested 12, 24, 36 and 48 h after resveratrol (100 μmol/L) treatment, respectively. Western blot analysis was applied to detect the protein expressions. The densitometry values of BDNF, GDNF and β-actin were analyzed and normalized to each respective control group. On the basis of ANOVA with Bonferroni's test analysis, consistent with the BDNF and GDNF production in the culture medium, astroglia-derived BDNF protein levels increased by up to 170% of control culture levels 24 h after resveratrol treatment and remained at 140% of control culture levels 36 h later (Figure 3a). The GDNF protein expression was increased by up to 140% of control cultures after resveratrol treatment for 24 h and peaked to 160% at 36 h and maintained at 130% of control cultures 48 h later (Figure 3b).
Resveratrol increased brain-derived neurotrophic factor (BDNF) and glial cell line-derived neurotrophic factor (GDNF) protein in astroglia cultures. The levels of BDNF (a) and GDNF (b) protein were determined by Western blot analysis after 100 μmol/L resveratrol treatment for different times. The densitometry values of target proteins (BDNF or GDNF) were normalized to β-actin of the same sample. Data are expressed as a percentage of the control cultures and are the mean ± SEM from three independent experiments performed in triplicate. *P < 0.05 compared with the control cultures (analysis of variance with Bonferroni's test)
Induction of CREB and ERK1/2 activation by resveratrol
Rat primary astroglia-enriched cultures were treated with resveratrol (25–100 μmol) for 60 min. The whole-cell lysates were analyzed by Western blotting assay. The ratio of densitometry values of phosphorylated CREB and ERK1/2 (p-ERK1/2) compared with total CREB and ERK1/2 was analyzed and normalized to each respective control group. On the basis of ANOVA with Bonferroni's test analysis, resveratrol induced the phosphorylation of CREB and ERK1/2 by up to 150% and 180% of control cultures, respectively, as shown in Figure 4.
Resveratrol induction of cAMP responsive element-binding protein (CREB) and extracellular signal-regulated kinase 1/2 (ERK1/2) in astroglia cultures. The CREB (a) and ERK1/2 (b) protein expressions were detected by Western blot assay 60 min after 100 μmol/L resveratrol treatment at different concentrations. Data are expressed as a percentage of the control cultures and are the mean ± SEM from three independent experiments performed in triplicate. *P < 0.05 compared with the control cultures (analysis of variance with Bonferroni's test)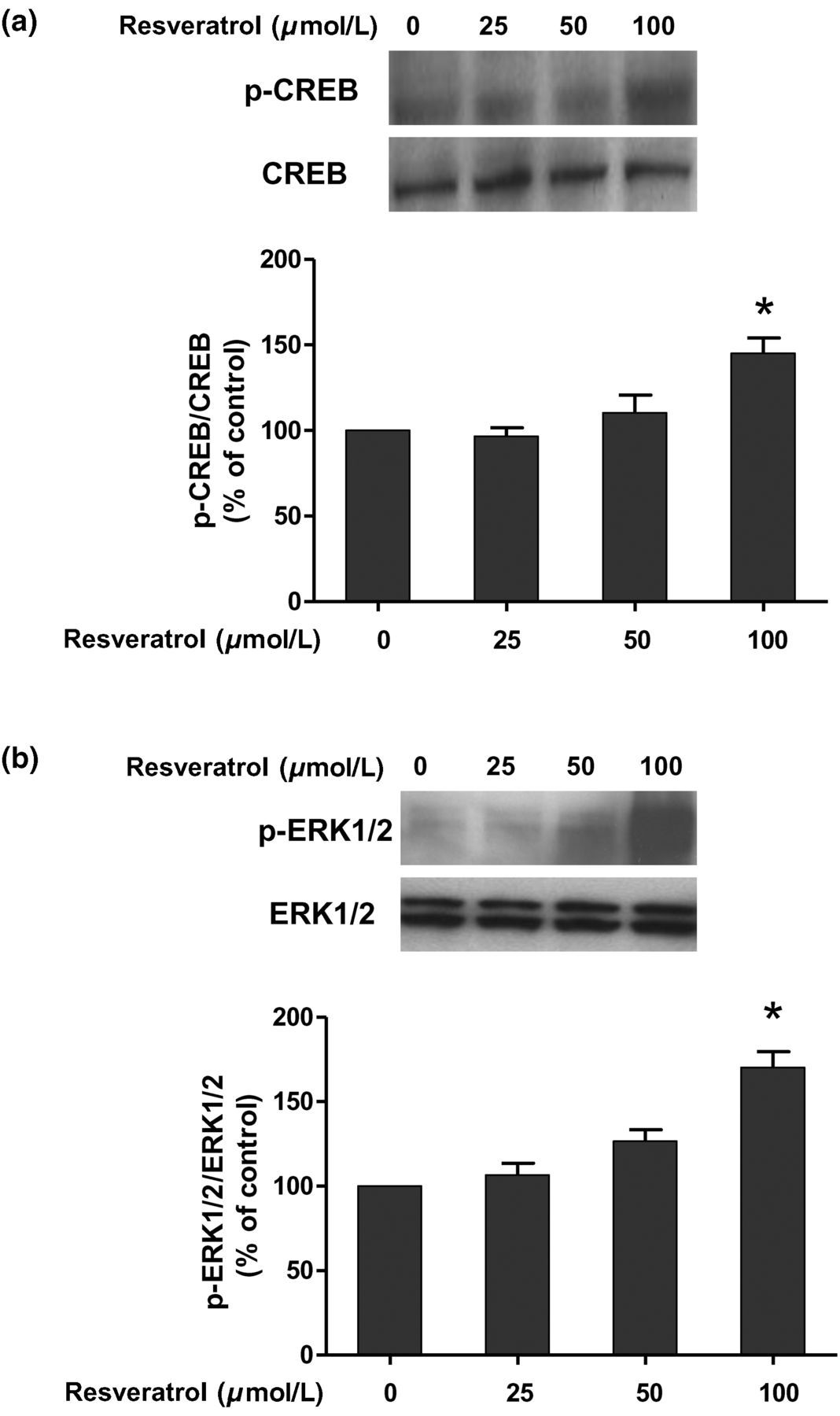
Discussion
This study indicated that resveratrol promoted the release of BDNF and GDNF neurotrophic factors from astroglia, as evidenced by ELISA of the culture medium and by Western blot analysis of astroglia protein. Astroglia and microglia are becoming the targets of neuroprotection. 1,7,9 Our previous studies have shown that microglia are involved in protection against LPS-induced neuroinflammation and dopaminergic neuronal damage by resveratrol 9,13 and other neuroprotective compounds. 15,16 Here, we further demonstrate that the neuroprotective effects of resveratrol also involve the activation of astroglia, with increased production of neurotropic factors, probably mediated through the activation of the CREB and ERK1/2signaling pathways.
A growing body of evidence indicates that astroglia play a critical role in the regulation of neurogenesis, synaptogenesis and neurotransmitter environments of the CNS. 17 By releasing various neurotrophic factors, astroglia also provide neuroprotection against neuronal damage. 18 BDNF is up-regulated in astroglia in the vicinity of injury sites in humans and experimental animals. 19 Upon activation, astroglia release BDNF to rescue neuronal cells from neuritic degeneration and neuronal death. 20,21 Several lines of evidence indicate that BDNF has beneficial effects on cognition, learning and memory formation by modulating synaptic plasticity. 22 Studies also show that the decreased level of BDNF was discerned in neurodegenerative diseases such as Alzheimer's disease, Huntington's disease, Parkinson's disease, depression, epilepsy and chronic pain. 23 In addition, GDNF has been recognized to play a crucial role in maintaining the physiological functions of the brain and pathological features of the CNS. 24 GDNF produced neuroprotection against ischemia, drug abuse and neuropathic pain. 25 Moreover, astroglia were reported to protect neurons through secreting various neurotrophic factors such as GDNF. 26 Besides BDNF, increasing evidence demonstrated that the alterations of the GDNF levels were detected in the peripheral blood and postmortem brain tissue of patients with mood disorders. 27,28 Therefore, promotion of BDNF and GDNF release is expected to have therapeutic potential for neurological disorders. This study is among the first to reveal that resveratrol significantly induced BDNF and GDNF production in astroglia cultures. This finding was consistent with a previous study in which resveratrol increased BDNF gene expression in the hippocampus of rat brain, 19,29 and extended this finding to astroglia, an important brain cell type in neuroprotection.
As rapid activation of mitogen-activated protein kinase kinase (MEK)/ERK/CREB pathways is critically involved in neurotrophic factor production in C6 glioma cells, 30,31 we next examined the role of ERK1/2 and CREB-signaling pathways in resveratrol-mediated BDNF and GDNF release. It has been reported that the ERK1/2 signaling pathway participates in the phosphorylation of CREB in cultured astrocytes. 32 Activation of CREB is observed to promote GDNF production in cultured astrocyte cells through CRE locating in the promoter sequence of the GDNF. 33,34 In addition, CREB siRNA can block the GDNF expression in astrocytes. 35 Therefore, the CREB signaling pathway represents a potential target responsible for inducing neurotrophic factor release.
Data from earlier clinical experiments indicate that GDNF was tested for its potential therapeutic treatment of Parkinson's disease by direct infusion into the brain of patients. However, this trial was terminated due to the occurrence of severe side-effects. 35–38 Thus, this study might provide an alternative approach for the treatment of neurological diseases by application of natural substances such as resveratrol to promote neurotrophic factors release.
Overall, this study clearly demonstrates that resveratrol is effective in promoting the release of neurotrophic factors BDNF and GDNF in rat astroglia cultures and that the ERK1/2 and CREB pathways might participate in resveratrol-induced neurotrophic effects.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by the National Natural Science Foundation of China (81102433 and IRT1197), National Key Basic Research Development Program (973 Program) Sub-project (2011CB512009), Science and Technology Foundation of Guizhou Province (20107030 and 20112316) and Doctor Foundation of Zunyi Medical College.