
Abstract
Chronic renal failure (CRF) is associated with an increased incidence of cardiovascular diseases. Intensive research revealed a number of alterations in the heart during CRF; however, possible interventricular differences in CRF-induced cardiac remodeling have so far not been addressed. CRF was induced by two-stage surgical 5/6 nephrectomy (NX) in male Wistar rats. Cellular hypertrophy was quantified using immunohistological morphometric analysis. Contraction force and membrane potential were recorded in left and right ventricle papillary muscles with an isometric force transducer and high-resistance glass microelectrodes. Hypertrophy was present in the left ventricle (LV) of NX animals, but not in the right ventricle (RV) of NX animals, as documented by both ventricle/body weight ratios and cellular morphometric analysis of the cross-sectional area of myocytes. The contraction force was reduced in the LV of NX animals but increased in the RV of NX animals compared with sham-operated rats. Rest potentiation of contraction force was relatively more pronounced in the LV of NX rats. Fifty percent substitution of extracellular sodium with lithium significantly increased the contraction force only in the LV of NX animals. Action potential durations were shortened in both ventricles of CRF animals. Cardiac structural and contractile remodeling in CRF shows significant interventricular differences. CRF induces hypertrophy of the LV but not of the RV. LV hypertrophy was associated with a reduction of contraction force, whereas in the RV, the contraction force was enhanced. Partial recovery of contractile function of the LV by rest potentiation or lithium substitution indicates a role of the Na+/Ca2+ exchanger in this phenomenon.
Introduction
In chronic renal failure (CRF), the incidence of cardiovascular complications, including hypertension, left ventricle (LV) hypertrophy, both systolic and diastolic dysfunctions, autonomic neuropathy and arrhythmia, is on average about 20 times higher compared with the normal population. Cardiovascular diseases (mainly heart failure, ischemic heart disease or sudden cardiac death) are the leading causes of death in CRF patients. 1 The clinical findings are supported by experimental animal studies. Both structural and functional changes in the heart have been described due to experimental CRF induced by subtotal nephrectomy (NX). 2–4 The alterations of cardiac contractile functions and of calcium homeostasis, changes in potassium and calcium currents, diastolic dysfunction and impairment of the autonomic cardiac innervation were all shown in subtotally nephrectomized rats. 5–13
In the mammalian heart, the right ventricle (RV) and LV have different hemodynamic functions. The RV pumps blood into low-pressure, low-resistance pulmonary circulation while the LV pumps the same volume of blood into high-pressure, high-resistance systemic circulation. 14 Afterload, the major determinant of ventricular function is nearly six-fold higher in the LV. 15 The heart ventricle-specific functions are associated with interventricular differences in morphology, contractile and electrophysiological properties that are well documented in a number of experimental studies. In physiological myocardium, diverse Ca2+ handling of sarcoplasmic reticulum, sarcomere shortening, calcium transients and afterload isotonic relaxation were described in the LV and the RV. 16–18 In adult rats, the action potential duration (APD) at a fixed heart rate was shorter in the RV probably because of a higher density of potassium transient outward current in the RV. 19
Some pathological processes are known to affect the ventricles differentially and to modify RV–LV differences, for example the impact of experimental diabetic cardiomyopathy on electrophysiological characteristics of the rat heart or the negative effect on the contractility of the end-stage heart failure in spontaneous hypertensive rats are more expressed in the LV. 20,21 Furthermore, a malfunction of one ventricle usually unfavorably alters the function of the other, a result of the adaptation process. 22–24 With regards to CRF, the majority of the experimental studies were performed either on the whole heart level or focused on LV function, while only minimal attention was paid to the impact of CRF on RV function. A direct comparison of the LV and RV effects of CRF is, to the best of our knowledge, so far completely lacking. Therefore, in this study, the structural, contractile and electrophysiological remodeling of both the LV and RV induced by CRF was investigated and compared. CRF was induced by subtotal (5/6) surgical NX and the remodeling was studied at early (10 days) and late (10 weeks) stages of renal failure development.
Materials and methods
All experiments were conducted in accordance with European Directive for the Protection of Vertebrate Animals Used for Experimental and Other Scientific Purposes (86/609/EU), the relevant Guidelines of the Czech Ministry of Agriculture for scientific experimentation on animals and were approved by the Faculty Committee for Experiments on Laboratory Animals (Faculty of Medicine in Plzen, Charles University in Prague, Plzen, Czech Republic).
Experimental model
Four-month-old Wistar male rats (Velaz, Prague, Czech Republic) were randomly allocated to subtotal NX (n = 45) or sham-operation (n = 15) groups. After anesthesia with sodium pentobarbital (100 mg/kg, intraperitoneal; Sigma-Aldrich, Prague, Czech Republic), surgical 5/6 renal mass reduction in two stages or sham operation was performed. Creatinine and urea concentrations in serum were measured using a routine autoanalyzer system (Olympus AU 640, Mishima, Japan). The plasma concentrations of peptides (calcitonin gene-related peptide [CGRP], neuropeptide Y [NPY]) were determined by radioimmunoassay using commercial kits (Phoenix Pharmaceuticals, Burlingame, CA, USA). Blood pressure was measured by the tail-cuff method using an RTBP apparatus (Kent Scientific Co., Torrington, CT, USA). Animals were sacrificed and the tissues used for experiments 10 days or 10 weeks after subtotal NX or sham operation.
Morphometric analysis
After fixation in formalin, ventricles were cut into six transversal slabs and six tissue blocks (size 4×4×4 mm) were taken from every second slab, three of them representing the free wall of the LV and another three representing the free wall of the RV. The tissue blocks were cut into histological sections and the basal laminae surrounding the outer circumference of cardiac myocytes were visualized by immunohistochemistry using a polyclonal rabbit anti-rat laminin antibody (DakoCytomation, Glostrup, Denmark). Due to the random rotation of the tissue blocks performed prior to embedding, the section plane was also oriented randomly. In each section, an area occupied by transversally cut cardiac myocytes was found and photographed. Twelve counting frames were superimposed on each micrograph in a systematic random uniform manner and the cross-sectional area of the first 10 myocytes found inside the counting frames and containing cell nuclei was estimated based on a ‘live wire’ algorithm implemented in the LiveWire module of the Ellipse software (ViDiTo, Košice, Slovak Republic). For each animal, 30 cells in the free wall of the LV and 30 cells in the free wall of the RV were measured by an observer blinded to the animal's history.
Contraction and electrophysiology experiments
After anesthesia, the hearts were excised and weighed. The papillary muscles were dissected from both ventricles, placed into an experimental chamber and attached to an isometric force transducer F30 (Hugo Sachs, March-Hugstetten, Germany). The preparation was perfused with 36°C warm, oxygenated Tyrode solution at a constant flow rate (6–10 mL/min). After a stabilization period (>30 min), we recorded: (1) the steady-state contractions and action potentials at stimulation frequencies 0.5, 1, 2 and 3 Hz; (2) post-rest potentiation contraction – steady-state (1 Hz) stimulation interrupted by a period of rest (10, 30, 60, 120 and 300 s). Contractions were measured in arbitrary units (a.u.). Time course of contraction–relaxation cycle was characterized using time-to-peak (time from resting tension to the peak of contraction, TTP) and time-to-90%-relaxation (R90). The resting tension was taken as zero. The membrane potential was measured with glass microelectrodes (filled with 3 mol/L KCl, resistance >20 MΩ). APD was measured at 50% and 90% levels of repolarization (APD50, APD90). In each animal, both RV and LV papillary muscles were measured and the measurement sequence was randomly varied to exclude possible error due to preparation rundown. Data were recorded and analyzed using the National Instruments data acquisition hardware and software (National Instruments, Austin, TX, USA).
Solutions and chemicals
The composition of the Tyrode solution was as follows (in mmol/L): NaCl 137, KCl 4.5, MgCl2 1, CaCl2 2, glucose 10, HEPES 5; pH adjusted to 7.4 with NaOH. In some experiments, NaCl was partially (50%) substituted with LiCl. Chemicals were obtained from Sigma-Aldrich.
Statistical analysis
Data are presented as mean ± SEM. After testing for normality of distribution, statistical comparisons were made with the paired or unpaired Student's t-test and with one-way analysis of variance followed by Bonferroni test, using software OriginPro 8.5 (OriginLab Corporation, Northampton, MA, USA). Differences at P ≤ 0.05 were considered significant.
Results
Renal failure
Serum creatinine and urea concentrations were already significantly higher in the NX group 10 days after surgery and they remained high throughout the experiment (up to 10 weeks, Figure 1). Daily volumes of urine had already increased (from 15.25 ± 1.38 to 45.1 ± 3.1 mL) and creatinine clearance dropped (from 2.2 ± 0.15 to 0.65 ± 0.05 mL/min) 10 days after surgery and remained on a similar level throughout the experiment. A substantial regeneration of the remnant kidney was observed; the weight of the remnant kidney in NX rats 10 weeks after surgery was higher than the weight of the intact kidney in control animals (2.1 ± 0.2 versus 1.36 ± 0.1 g).
Renal failure and hypertension. Open squares, sham-operated rats (n = 7); filled circles, NX rats (n = 8). *Significantly different from sham-operated rats, P < 0.05. (a) Plasma concentrations of urea. (b) Plasma concentrations of creatinine. (c) Systolic and diastolic blood pressures. NX, nephrectomy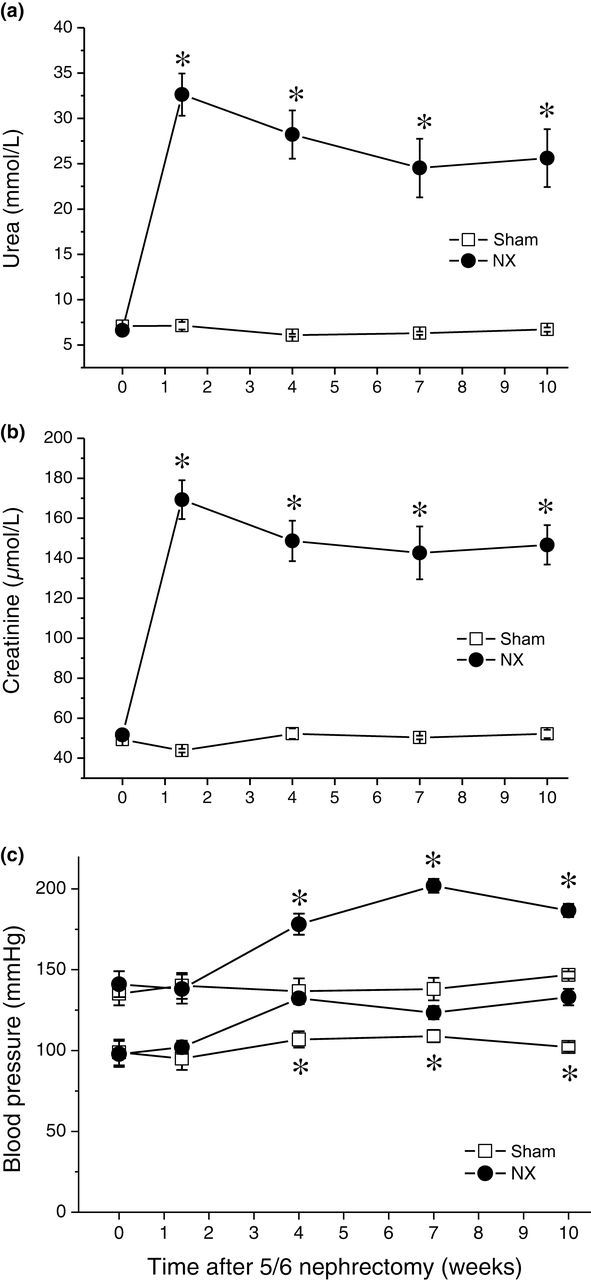
Cardiac hypertrophy
In NX rats, a pronounced hypertension developed (Figure 1). Both systolic and diastolic pressures were significantly increased in NX rats four weeks after surgery and further. Ten days after surgery, however, neither systolic nor diastolic pressures were different from control (Figure 1). Heart weight was significantly increased in NX rats and this increase was exclusively due to the LV (including septum); the weight of the RV was not different from control (Figure 2). The increase in the heart weight was already present 10 days after surgery and it persisted throughout the experiment, becoming even more pronounced 10 weeks after surgery (Figure 2). To verify cellular hypertrophy as the underlying cause of the heart weight increase, the cellular morphometric immunohistological analysis of cardiac tissue was performed in hearts 10 weeks after surgery. In accordance with the heart weight measurements, the cross-sectional area of cardiac myocytes was clearly increased in the LV of NX rats, whereas no difference between control and NX animals was observed in the RV (Figure 2).
Hypertrophy of the heart. Empty bars, sham-operated rats (n = 7); filled bars, NX rats (n = 8). *Significantly different from sham-operated rats, P < 0.05. (a) Absolute (left panel, BW, body weight, HW, heart weight, LV, left ventricle + septum weight, RV, right ventricle weight) and relative (right panel, HW, heart weight/body weight ratio, LV, left ventricle + septum weight/body weight ratio, RV, right ventricle weight/body weight ratio) weights 10 days after surgery. (b) Absolute (left panel, BW, body weight, HW, heart weight, LV, left ventricle + septum weight, RV, right ventricle weight) and relative (right panel, HW, heart weight/body weight ratio, LV, left ventricle + septum weight/body weight ratio, RV, right ventricle weight/body weight ratio) weights 10 weeks after surgery. (c) Left panel, microphotography of cardiac tissue. Immunohistochemistry against laminin, basal laminae surrounding the myocytes visualized. Right panel, cross-sectional areas of cardiac myocytes. #Significantly different from RV, P < 0.05. NX, nephrectomy. (A color version of this figure is available in the online journal)
The plasma concentrations of CGRP were similar in sham-operated and NX rats 10 days after surgery (126.2 ± 14.4 pg/mL in sham rats versus 145.3 ± 15.4 pg/mL in NX rats) but were significantly increased in NX rats 10 weeks after surgery (163.6 ± 11.1 pg/mL in NX rats versus 115.1 ± 8.7 pg/mL in sham rats, P < 0.05). A similar pattern was found for plasma concentrations of NPY: no difference between sham-operated and NX animals 10 days after surgery (5083.2 ± 802.4 pg/mL in sham versus 4318.8 ± 1022.7 pg/mL in NX rats), but a significant rise was present in NX rats 10 weeks after surgery (6204.5 ± 407.8 pg/mL in NX rats versus 4322.4 ± 465.1 pg/mL in sham rats, P < 0.05).
Cardiac contraction
Contraction force (CF) in control animals was significantly higher in the LV compared with the RV at stimulation frequencies 1, 2 and 3 Hz at both time points: 10 days as well as 10 weeks after surgery. In NX rats, the contractile remodeling was different in the LV and RV. In the LV, CF was reduced at all stimulation frequencies studied (0.5–3 Hz) and at both points in time (Figure 3). On the other hand, in the RV, the CF was increased, again at all stimulation frequencies and at both time points (Figure 3). Consequently, the relationship of LV and RV contractility was completely reversed; whereas in control animals LV CF was higher than RV CF, in NX animals, LV CF was lower than RV CF (Figure 3).
Steady-state contraction force (CF). (a) Representative examples. Left panel, contraction in left ventricle of sham-operated (n = 7) and NX (n = 8) rats. Right panel, contraction in right ventricle of sham-operated and NX rats. (b) Dependence of CF on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles) rats 10 days after surgery. *Significantly different from sham-operated rats, P < 0.05. (c) Dependence of CF on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles) rats 10 weeks after surgery. *Significantly different from sham-operated rats, P < 0.05. NX, nephrectomy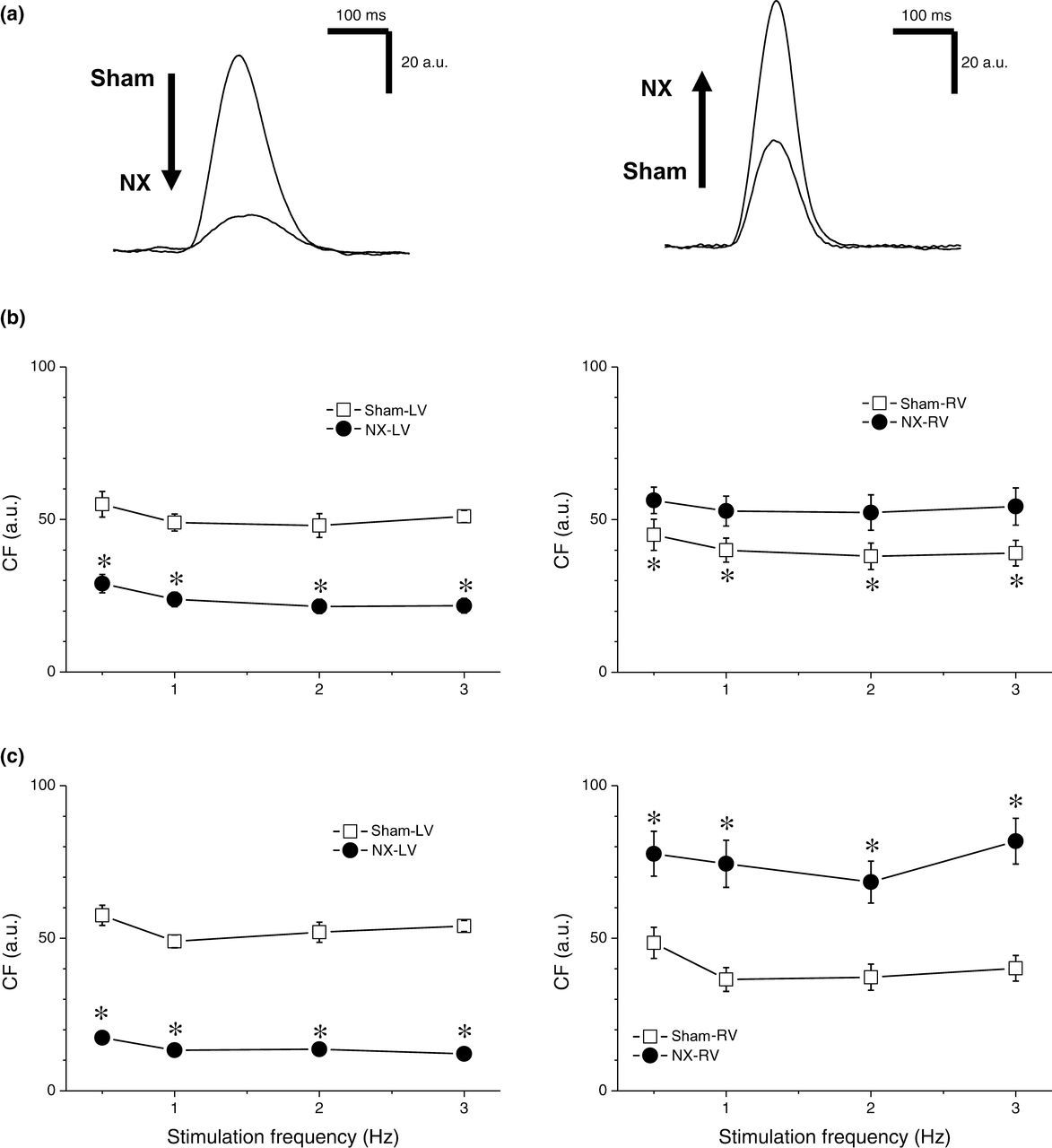
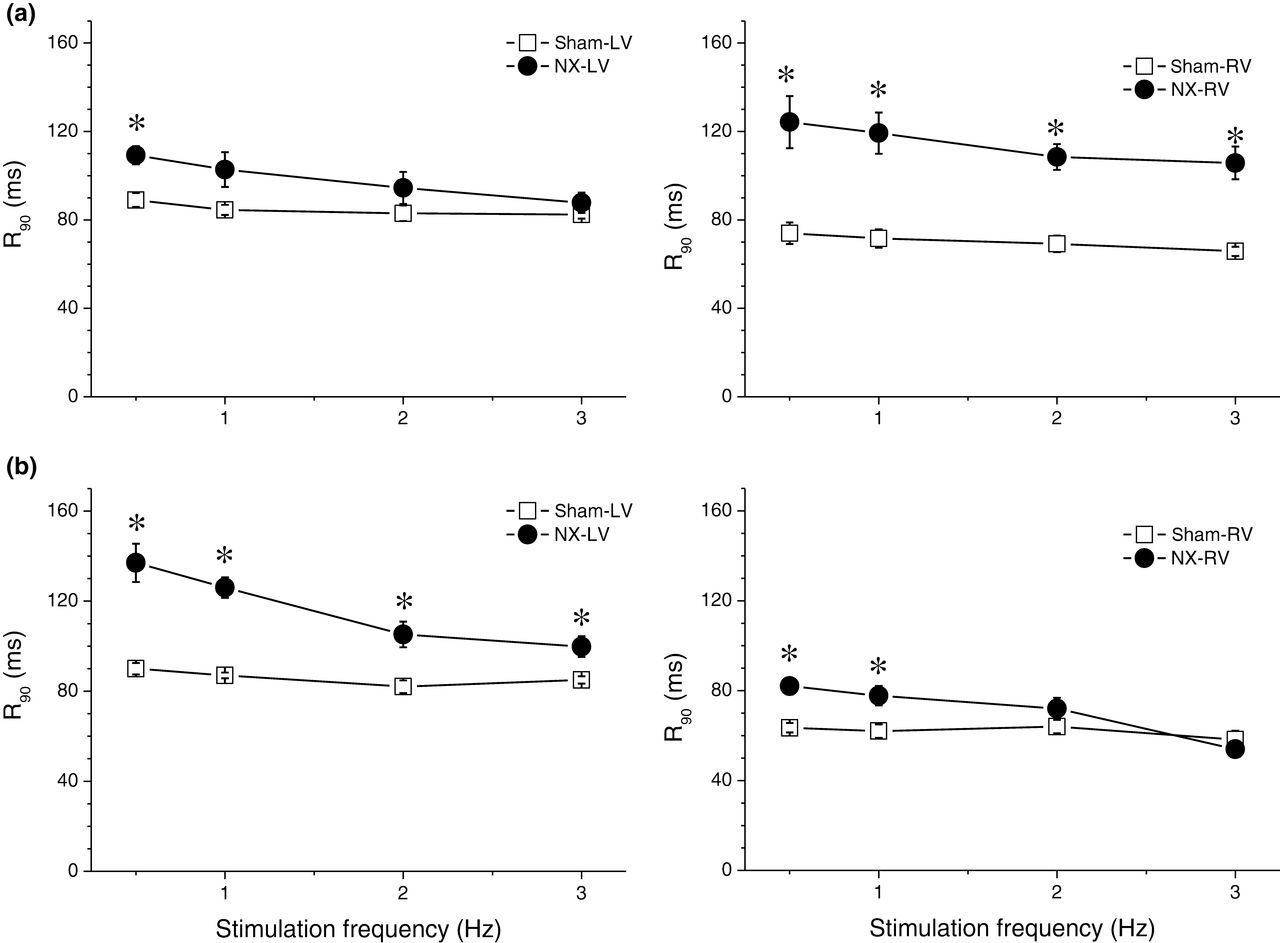
With regard to the kinetics of the contraction–relaxation cycle, TTP and time from peak of contraction to 90% relaxation (R90) were measured. In control rats, TTP and R90 were significantly shorter in the RV than in the LV at all stimulation frequencies tested both 10 days and 10 week after sham operation. NX induced prolongation of TTP and R90 in both ventricles, but the kinetics of the change was different. In the LV, 10 days after surgery, both TTP and R90 were prolonged only at low stimulation frequencies, while 10 weeks after surgery, the effect was more pronounced and TTP and R90 were prolonged at all stimulation frequencies tested (0.5–3 Hz, Figures 4 and 5). In contrast, in the RV, both TTP and R90 were already prolonged significantly at all stimulation frequencies tested 10 days after surgery. Ten weeks after surgery, the effect was reduced, leaving a significant prolongation of both TTP and R90 only at low frequencies (Figures 4 and 5).
Time to peak of contraction (TTP). (a) Dependence of TTP on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares, n = 7) and NX (filled circles, n = 8) rats 10 days after surgery. *Significantly different from sham-operated rats, P < 0.05. (b) Dependence of TTP on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles) rats 10 weeks after surgery. *Significantly different from sham-operated rats, P < 0.05. NX, nephrectomy Time from peak of contraction to 90% relaxation (R90). (a) Dependence of R90 on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares, n = 7) and NX (filled circles, n = 8) rats 10 days after surgery. *Significantly different from sham-operated rats, P < 0.05. (b) Dependence of R90 on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles) rats 10 weeks after surgery. *Significantly different from sham-operated rats, P < 0.05. NX, nephrectomy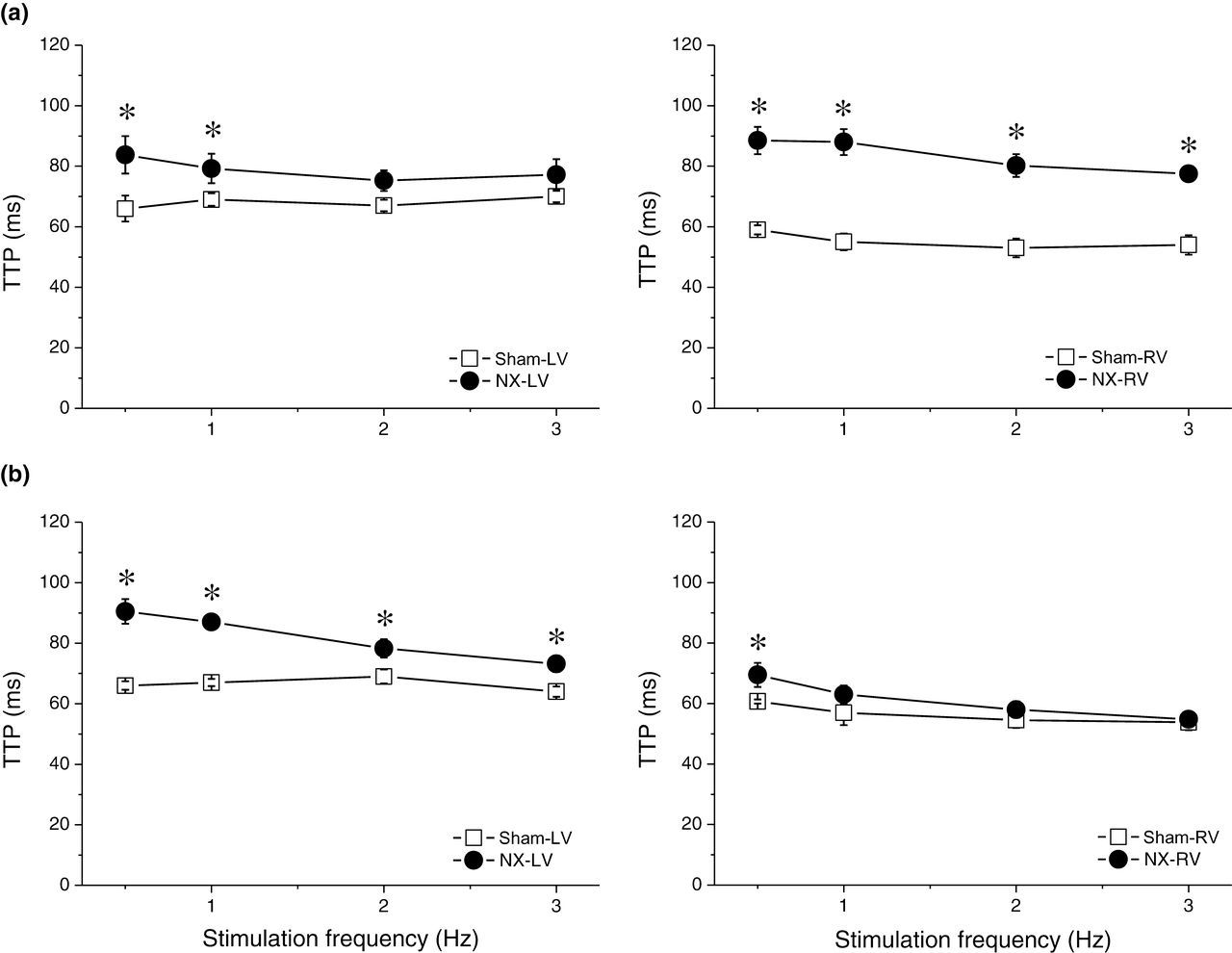
The rest-potentiation phenomenon was more pronounced in the LV than in the RV (Figure 6). NX further enhanced the relative rest potentiation (ratio of the first post-rest CF and of steady-state CF) in the LV, but suppressed it in the RV (Figure 6), regardless of the duration of period of rest (10–300 s). The rest-potentiation phenomenon, therefore, was able to either partially (LV) or fully (RV) compensate for the changes in steady-state CF induced by NX: enhanced rest potentiation for reduced steady-state contraction in the LV and reduced rest potentiation for increased steady-state contraction in the RV (Figure 6).
Rest potentiation of contraction. (a) Representative examples. Left panel, steady-state contraction and the first post-rest contraction in the left ventricle of sham-operated (left, n = 7) and NX (right, n = 8) rats. Right panel, steady-state contraction and the first post-rest contraction in the right ventricle of sham-operated (left) and NX (right, 10 weeks after surgery) rats. (b) Dependence of rest potentiation (ratio of the first post-rest contraction and the steady-state contraction) on duration of rest period in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles, 10 weeks after surgery) rats. *Significantly different from sham-operated rats, P < 0.05. (c) Dependence of CF of the first post-rest contraction on duration of rest period in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles, 10 weeks after surgery) rats. *Significantly different from sham-operated rats, P < 0.05. NX, nephrectomy; CF, contraction force; RP, rest period
Fifty percent substitution of extracellular sodium with lithium, an intervention enhancing the reverse mode of sodium–calcium exchanger, elicited a positive inotropic effect only in the LV of NX animals (Figure 7). The effect was more pronounced at low stimulation frequencies. In the LV of sham-operated rats and in the RV of both sham-operated and NX rats, no significant effect was observed despite the positive tendency (Figure 7).
Effects of partial (50%) substitution of extracellular Na+ with Li+ on contraction. (a) Representative examples of contraction in control Tyrode solution and in solution with Li+ substitution in the left ventricle (LV) from sham-operated rats (n = 7), LV from NX rats (n = 7), right ventricle (RV) from sham-operated rats and RV from NX rats (from left to right), all 10 weeks after surgery. Li+ substitution tended to increase CF in all preparations; however, only in the LV from NX rats, the effect was significant. *Significantly different from contraction in control Tyrode solution, P < 0.05. (b) Dependence of CF on stimulation frequency in the LVs (left panel) and RVs (right panel) of sham-operated (squares) and NX (circles, 10 weeks after surgery) rats in control Tyrode solution (open symbols) and in solution with 50% Li+ substitution (filled symbols). *Significantly different from contraction in control Tyrode solution, P < 0.05. (c) Normalized representative examples (same as in a) of contraction in control Tyrode solution and in solution with Li+ substitution in the LV from sham-operated rats, LV from NX rats, RV from sham-operated rats and RV from NX rats (from left to right), all 10 weeks after surgery. *Significantly different from contraction in control Tyrode solution, P < 0.05. (d) Relative effect of Li+ substitution on CF (ratio of CF in solution with 50% Li+ substitution and in control Tyrode solution). Dependence of CF ratio on stimulation frequency. *Significantly different from RV, P < 0.05. NX, nephrectomy; CF, contraction force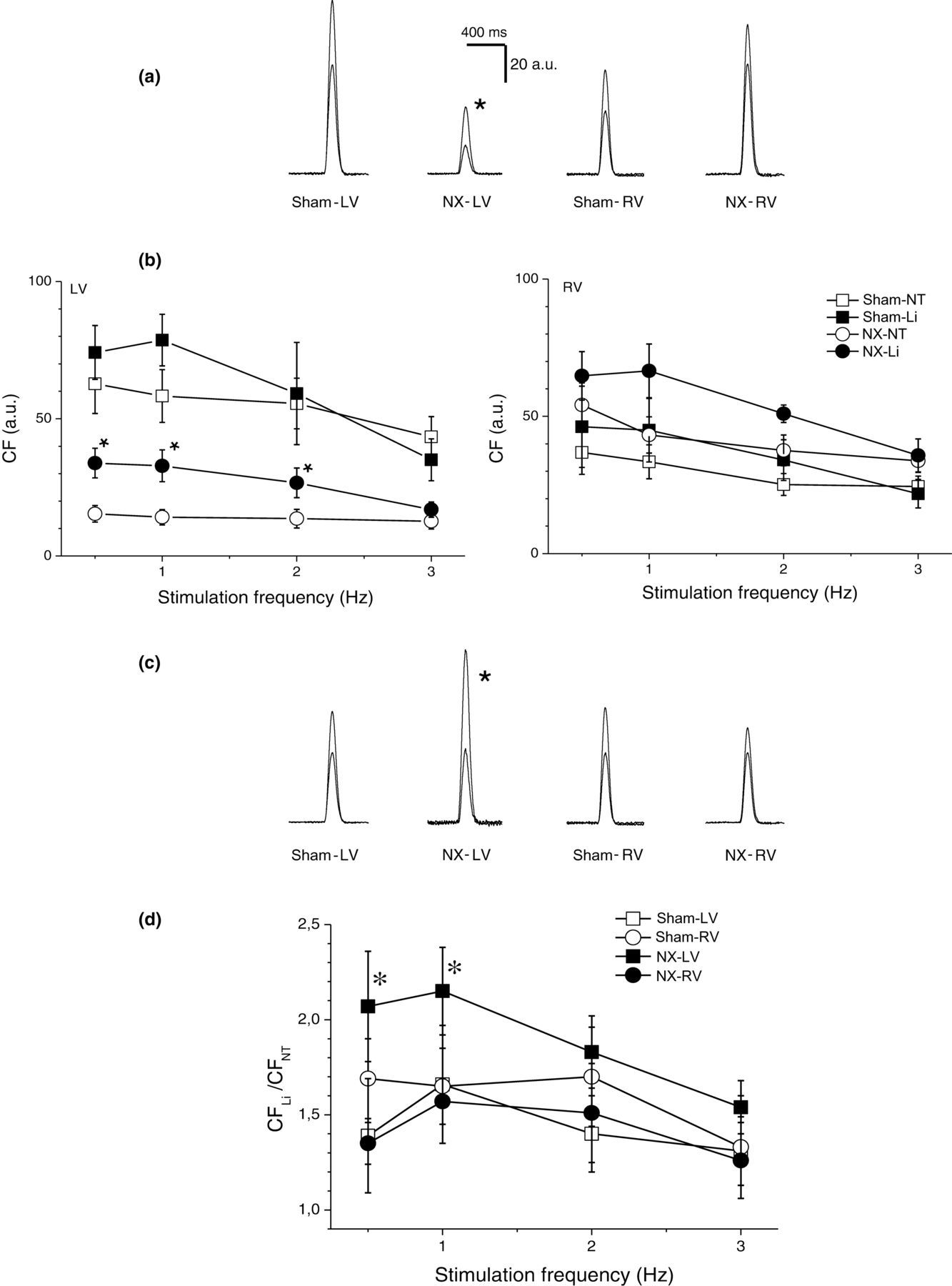
Cardiac electrophysiology
APD90 was significantly shorter in the RV than the LV in both experimental groups (Figure 8). NX led to a shortening of APD90 in both the RV and LV that only became significant 10 weeks after surgery (Figure 8). The RV versus LV difference in APD90 was not influenced by NX, for example LV-RV difference of 39.3 ± 3.8 ms in sham-operated animals and of 36.3 ± 2.9 ms in NX rats. Similar results were obtained also for APD50 (not shown).
Action potential duration (APD). (a) Representative examples of action potentials from sham-operated (upper panels, n = 7) and NX (lower panels, n = 8) rats in left (left panels) and right (right panels) ventricles. Horizontal bars, 0 mV. (b) Dependence of action potential duration at 90% level of repolarization (APD90) on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles) rats 10 days after surgery. (c) Dependence of APD90 on stimulation frequency in left (left panel) and right (right panel) ventricles of sham-operated (open squares) and NX (filled circles) rats 10 weeks after surgery. *Significantly different from sham-operated rats, P < 0.05. NX, nephrectomy
Discussion
In this model of renal failure, the RV and LV were remodeled completely differently. The hypertrophy of myocytes occurred in the LV, but not in the RV. The hypertrophy in the LV was associated with a reduction of contractile force, whereas in the RV, an improved contractility was observed. The impaired contractility in the LV could be compensated by interventions enhancing the contribution of the sodium/calcium exchanger to the contraction, thus indicating an important role of this transporter in the phenomenon. In contrast to structural and contractile remodeling, the electrophysiological remodeling was similar in both ventricles, thus maintaining the interventricular electrophysiological heterogeneity.
Cardiac hypertrophy is a common finding in CRF. 25 Most studies, however, were focused on the LV, hypertrophy of which was shown in experimental (e.g. ref. 26 ) as well as clinical studies (e.g. ref. 27 ). The remodeling of the RV was investigated to a much lesser extent. Our data indicate that the response of the RV to NX is different from that of the LV: whereas a clear increase in tissue weight and cardiac myocyte dimensions was present in the LV, in the RV, no significant changes in either tissue weight or cellular dimensions were found. Similarly, the histological analysis in NX rats four weeks after surgery revealed an increase in LV tissue cross-sectional area but no change in RV tissue area. 6 Experimental acute kidney injury by subtotal NX induced in 10 days a significant increase in the weight of both the LV and RV, although the increase in the weight of the LV was much more pronounced. 28 A subtle methodological difference could contribute to this discrepancy: whereas in our study the partial NX of one kidney (beside the complete resection of the other kidney) was done by surgical resection of kidney poles, in the study by Burchill et al., 28 it was done by ligation of renal artery branches.
Development of the hypertrophy was fast; a significant hypertrophy was already present 10 days after surgery. This corresponds well with the study of Burchill et al., 28 in which a significant hypertrophy was also observed 10 days after subtotal NX. Such a fast development of hypertrophy and comparison with the time course of development of other phenomena (hypertension) provides interesting insights into the pathogenesis of the disease. It is generally believed that the LV hypertrophy associated with chronic renal disease results from both pressure and volume overload together with humoral factors. 29–32 Pressure overload is assumed to be secondary to hypertension and loss of elasticity of arteries. 32 Comparison of time courses of development of LV hypertrophy and hypertension in this study, however, documents that the hypertrophy precedes the hypertension. At day 10 after surgery, the LV hypertrophy was already present, which was not the case for the hypertension. Similarly, in another study, four weeks after NX, hypertrophy of LV was present but there was no evidence of hypertension. 6 On the other hand, structural changes in aorta were consistent with hypertension development. Significant structural changes in the aortic wall were only found 10 weeks after NX, but they were not present at the early time point (10 days after surgery). 33 Taken together, these data argue against a significant role of pressure overload in the (at least early) development of LV hypertrophy in NX rats. Volume overload should affect both the LV and RV. In a well-described non-renal model of volume overload, dog with chronic atrioventricular block, the chronic volume overload led to biventricular hypertrophy, confirmed on a cellular level. 34,35 The absence of hypertrophy in the RV may suggest that the volume overload is not a critical contributor to the pathogenesis of LV hypertrophy in NX rats. That leaves the humoral hypothesis as the most likely alternative, and the fast and early rises in urea and creatinine plasma concentrations documented in this study are in agreement with this hypothesis. Although several substances were already suggested as possible contributors to the development of cardiovascular complications in renal disease (e.g. parathyroid hormone and vitamin D), 36 it may be assumed that the majority of humoral modulators involved in the pathogenesis of cardiorenal syndrome still remains obscure. From a vast number of possible candidates, the peptides CGRP and NPY were reported to induce hypertrophy of ventricular cardiomyocytes. 37,38 Furthermore, enhanced neuronal expression of CGRP was found in a mineralocorticoid-salt hypertension rat model that included left NX. 39 Serum NPY concentration was increased in rats with CRF and higher plasma NPY concentrations were found in patients with left ventricular hypertrophy and end-stage renal disease. 40,41 In this study, the plasma concentrations of both CGRP and NPY were found to be increased. The increase, however, was delayed, being significant 10 weeks after NX but not at the early time point. Since a significant hypertrophy was already present 10 days after NX, a significant role of CGRP and/or NPY in (early) development of hypertrophy is not likely. The modern high-throughput methods (proteomics) will have to be employed to screen for critical regulators in both plasma and tissues involved and to identify new therapeutic and diagnostic targets.
The most striking finding of the study was the differential contractile remodeling. Impaired contractility in the LV contrasted with a rather improved contractility in the RV. The reduced contractility in the LV could be partially compensated by two interventions: rest-potentiation and partial (50%) substitution of extracellular sodium with lithium. The rest potentiation, i.e. increase in CF after period of rest, is a phenomenon characteristic for rat myocardium and it reflects the function of the Na+/Ca2+ exchanger. 42–45 Based on a high intracellular concentration of Na+ in rat myocardium, a net Ca2+-influx occurs during rest leading to an increased sarcoplasmic reticulum Ca2+-loading and consequently to potentiation of the post-rest contraction. The enhanced rest potentiation in the LV from NX rats therefore indicates a more pronounced role of the Na+/Ca2+ exchanger compared with the other preparations. To further confirm this hypothesis, experiments with partial (50%) substitution of extracellular Na+ were performed. Reduction of extracellular Na+ shifts the balance of the Na+/Ca2+ exchange transport in favor of the reverse mode (Ca2+ inward, Na+ outward) and therefore a positive inotropic effect should occur. Again, a significant positive inotropic effect was observed only in LV preparations from NX rats, supporting an increased role of the Na+/Ca2+ exchanger in contraction of these preparations. Furthermore, this positive inotropic effect was only present at low stimulation frequencies (0.5–2 Hz), at which the diastolic Ca2+ loading via the exchanger should be more prominent. Consistent with this line of thinking, an increased expression of Na+/Ca2+ exchanger protein was described in the LV of NX rats. 7 In the same study, decreased enzymatic activity and protein expression of the Na+/K+-ATPase were also reported. 7 This could result in an increase in intracellular Na+ concentration that would again enhance the reverse mode of the Na+/Ca2+ exchanger.
The electrophysiological remodeling was similar in both ventricles: a shortening of APD90 was observed. In contrast to this finding, in patients with CRF, a prolongation of the QT interval was reported, both pre- and post-dialysis. 46 This discrepancy probably reflects an interspecies difference since other studies in the rat model of renal failure also showed an APD shortening that was associated with alterations in transient outward potassium current and/or L-type calcium current. 8,9 In general, Ca2+ influx during action potential initiates the contraction by Ca2+ release from sarcoplasmic reticulum and APD is known to be one of the most important inotropic factors. 47,48 Obviously, the electrophysiological remodeling being similar in both ventricles cannot explain the substantial differences in contractile remodeling of the ventricles.
Limitations of the study
The data that suggest a possible role of the Na+/Ca2+ exchanger in the differential contractile remodeling represent only indirect evidence. With regards to the role of the Na+/Ca2+ exchanger in rest potentiation, it should be mentioned that there is also a significant body of evidence that suggests that the rest potentiation may not depend on the sarcoplasmic reticulum Ca2+ content (i.e. on the Na+/Ca2+ exchanger), but rather may be attributable to recovery of the excitation–contraction coupling and to increased fractional release of Ca2+ from the sarcoplasmic reticulum. 49,50 Therefore, the function of the Na+/Ca2+ exchanger must be carefully analyzed in the context of all contraction processes that, in the setting of CRF, still need to be scrutinized in more detail.
The blood pressure was only measured in systemic circulation. To elucidate more the relationship between hypertension and hypertrophy, it would be worthwhile to measure also the blood pressure in pulmonary circulation. The lack of hypertrophic response in the RV, however, argues against a significant pressure challenge in the pulmonary circulation.
Two possible regulators of the hypertrophic response, CGRP and NPY, were studied and the data indicate that they are not the critical determinants of the hypertrophy induction. A thorough screening of the tissues affected and the plasma using the modern high-throughput methods will be necessary to narrow the search for signaling molecules and to define new therapeutic targets. Such a tremendous task, however, is beyond the scope of this study.
Conclusions
In rats with subtotal NX, a significant structural, contractile and electrophysiological remodeling of the heart occurs. The structural and contractile remodeling shows substantial interventricular differences; electrophysiological remodeling was similar in both ventricles. Hypertrophy of cardiac myocytes only occurred in the LV, but not in the RV. Contractility was impaired in the LV but rather improved in the RV. The Na+/Ca2+ exchanger is probably involved in the differential contractile remodeling of the ventricles.
Footnotes
ACKNOWLEDGEMENTS
The authors thank Ivana Hajdúková for excellent technical assistance.
The study was supported by the Research Project MSM 0021620819: Replacement of and Support to Some Vital Organs awarded by the Ministry of Education, Youth and Sports of the Czech Republic.