
Abstract
The development of the prostate depends on a precise androgenic control, so sensible interferences may predispose this gland to develop prostatic diseases during life. These aspects are of interest and preoccupation, since human beings are exposed to a growing number of endocrine-disrupting chemicals with androgenic potential. Therefore, our aim was to evaluate the prostates of adult gerbils exposed to testosterone during intrauterine life. Serological, morphological, morphometric-stereologic, immunohistochemical and three-dimensional reconstruction analyses were used. We found that the testosterone effects were dose-dependent and more harmful to females, leading to the development of masculine characteristics, evidenced by an increased anogenital distance, and absence of vaginal opening and the ectopic development of prostatic tissue. Moreover, premalignant lesions, such as prostatic intraepithelial neoplasia, were observed in addition to inflammatory foci in the prostate. The results showed that the prenatal exposure to testosterone may affect the reproductive system, disrupting developmental processes and increasing susceptibility to the development of prostatic diseases in the Mongolian gerbil.
Introduction
Endocrine-disrupting chemicals (EDCs) are exogenous substances that can potentially disrupt the endocrine system by affecting the metabolism of hormones.1,2 Besides this peculiarity, these substances may even act directly on the organs of the reproductive system in both males and females by competing with endogenous steroids for active sites on specific receptors.
The number of EDCs with androgenic potential to which humans and wildlife are exposed has been increasing. Some of them, such as methyltestosterone, 3 trenbolone acetate 4 and methlydihydrotestosterone, 5 were demonstrated to be present in the environment. Besides environmental contamination, there are several cases in which pregnant women make use of androgenic compounds. These substances may act as EDCs, reaching the fetuses and increasing the susceptibility to the development of malformations of reproductive organs.
The number of studies evaluating the effects of EDCs with estrogenic potential on the reproductive tract of rodents, especially on the prostate, has increased recently.6–8 However, few studies have assessed the effects of intrauterine exposure to EDCs with androgenic potential on the prostate.
It has been suggested that the susceptibility to diseases, such as benign prostatic hyperplasia (BPH) and prostate cancer (PCa) initiates earlier in life, mainly due to abnormal androgen exposure, especially during development. 9 However, the mechanisms involved in the predisposition induced by abnormal androgen exposure and its relationship with the manifestation of prostatic diseases during adult and senile life are unknown. 10
Investigations with mice exposed to dihydrotestosterone during the prenatal phase have shown that gene expression programming, which is controlled by the androgen receptor (AR) pathway and is responsible for guiding prostate organogenesis, is also activated during prostatic cancer installation. 9 These aspects show that this gene expression programming is present either during processes that give rise to the initiation and progression of the PCa or during prostatic organogenesis. These studies have shown that AR-dependent genes that are associated with events characteristic of the carcinogenesis process, such as angiogenesis, apoptosis, migration, motility and proliferation, are similar to genes that are expressed during the prostatic development. 9 These aspects are important, especially because more than a hundred AR-dependent genes are expressed and finely regulated during prostatic organogenesis. 11
Thus, the detailed knowledge of how EDCs with androgenic potential may affect the prostate development may provide some important information about the susceptibility of this gland, when exposed to exogenous androgenic substances, to develop lesions throughout life. Moreover, these studies may even provide some idea of how earlier abnormal events during prostate organogenesis may be involved in the promotion of prostatic lesions throughout life.
Studies carried out in this laboratory have shown that the Mongolian gerbil (Meriones unguiculatus) is a good model for studies involving drugs and hormonal manipulation.12–14 This characteristic is due to the fact that the gerbil prostate is sensible to both endogenous and exogenous hormonal imbalances. Moreover, previous work 15 has shown that female gerbils possess prostatic tissue that is quite similar to the ventral prostate of male gerbils, which allows these kinds of studies with females of this species.
Studies on the female prostatic tissue (also known as Skene's gland) of several rodent species have been previously published in the literature.15–19 Although some studies have reported some female rodents with a low frequency of prostatic tissue,16,19 the Mongolian gerbil presents a high incidence of this gland between the females of this species. 15 This high incidence of prostate tissue in female gerbils simulates what occurs in women who present a high incidence of prostatic tissue, 20 a fact that has already been discussed in the literature.
Taking these aspects into account, our hypothesis is that the testosterone exposure during prostatic embryogenesis may disrupt the normal developmental process of the prostate leading to the development of prostatic lesions, such as BPH and prostatic intraepithelial neoplasia (PIN), in both male and female adult gerbils. Thus, our aim in this work was to evaluate the impact of testosterone cypionate exposure during the embryonic phase on the prostate of male and female adult gerbils using morphological, stereo-logic–morphometric, immunohistochemical and three-dimensional (3D) reconstruction analyses.
Material and methods
Animals and experimental design
The animals were provided by the São Paulo State University (UNESP) (São José do Rio Preto), were maintained in polyethylene cages under controlled conditions of light and temperature and were provided with filtered water and rodent food ad libitum. Animal handling and experiments were performed according to the ethical guidelines of the São Paulo State University (UNESP), following the Guide for Care and Use of Laboratory Animals (NIH, Bethesda, MD, USA).
Fifteen adult female and 15 adult male (90 days old) gerbils (M. unguiculatus, Muridae: Gerbillinae) were used. One male was matched with each female to form independent families. Pregnant females from these couples underwent different manipulations and their offspring formed the experimental groups as follows: C (control) group: offspring from non-manipulated pregnant females; LT (low testosterone during gestation) group: offspring from mothers exposed to a single subcutaneous injection of 100 μg of T (testosterone cypionate – Deposteron; EMS Sigma-Pharma, São Bernardo do Campo, São Paulo, Brazil) diluted in 100 μL corn oil during gestation; HT (high testosterone during gestation) group: offspring from mothers exposed to a single testosterone subcutaneous injection of 1 mg of T diluted in 100 μL of corn oil during gestation. Only the pups exposed to testosterone that were born six to eight days after treatment were employed in this study. All animals utilized in this study were killed within 120 days of birth. The protocol of T treatment was adapted from Wolf et al.
21
The overall experimental design, treatment windows for T and the age at which the animals were killed are shown in Figure 1.
Schematic representation of the experimental protocol employed in this study. The days (day 0 – day of mating and day 0 (P0) – day of birth) represent the gerbil gestational period. The length from day 9 to day 0 (P0) shows the countdown from the ninth day before the birth of the pups. Groups LT (low testosterone) and HT (high testosterone) were exposed between days 6 and 8 with a single injection of testosterone and the pups were born six to eight days later. P0–P120 (postnatal days). P120 represents the age of the animals at death. (A color version of this figure is available in the online journal)
All animals were killed by CO2 inhalation followed by decapitation. Only females (groups C and LT) in the proestrus phase were killed, according to Nishino and Totsukawa. The females of the HT group were not cycled due to the absence of a vaginal opening. Blood samples were also collected. The body, prostatic complex of males (UPT - urethra plus ventral, dorsal and dorsolateral prostate), prostatic complex of females (UPT - urethra plus prostate), testicles, ovaries and adrenal glands were all weighed. These fragments were dissected out using an Olympus SD-ILK stereoscopic microscope (Olympus Optical Co. Ltd, Tokyo, Japan) to remove adipose tissue and isolate the urethral segment plus the associated prostatic tissue.
Plasma total testosterone and estradiol dosage
Blood serum samples of female and male gerbils were collected after decapitation followed by the rupture of the cervical vessels. The serum was separated by centrifugation (3000 rpm) and stored at −20°C for subsequent hormone analysis. Circulating serum testosterone and estradiol concentration were determined via chemiluminescence immunoassay in a Vitros-ECi automatic analyzer (Johnson & Johnson, Orthoclinical Diagnostics Division, Rochester, NY, USA). The sensitivity was ≥ 0.02ng/mL for testosterone and ≥5 pg/mL for estradiol.
Light microscopy
UPTs from male and female gerbils were fixed by immersion in Karnovsky's solution (5% paraformaldehyde, 2.5% glutaraldehyde in 0.1 mol/L phosphate buffer, pH 7.2). After fixation, the tissues were washed with running tap water, dehydrated in an ethanol series, clarified in xylene and embedded in Historesin using an embedding kit (Leica, Nussloch, Germany). Tissue sections (3 μ m) were obtained with an automatic rotator microtome (Leica RM2155) and stained with hematoxylin-eosin (HE) for general morphological analysis. Prostatic reticular fibers were identified by Gömöri's reticulin. The specimens were analyzed with an Olympus BX60 light microscope (Olympus, Hamburg, Germany), and the images were digitalized using the software DP-BSW V3.1 (Olympus) and Image-Pro Plus version 6.1 for Windows (Media Cybernetics Inc., Silver Spring, MD, USA).
Morphometry and stereology
The stereological analyses were carried out using Weibel's multipurpose graticulate with 130 points and 10 test lines to compare the relative proportion (relative volume) of each component of the prostatic tissue (epithelium, lumen and stroma), as described by Huttunen et al. 24 Thirty microscopic fields were chosen at random from each experimental group (six fields for animal; N =5). In summary, the relative values were determined by counting the coincident points in the test grid and dividing them by the total number of points. Morphometric analysis also included the determination of epithelial cell height and smooth muscle layer thickness. All morphometric parameters were assessed using the software Image-Pro Plus version 6.1 for Windows (Media Cybernetics Inc.).
Immunohistochemistry
Prostatic tissues were fixed in 4% paraformaldehyde for 24 h and in metacarn (proportions: methanol 60%, chloroform 30% and acetic acid 10%) for three hours. After fixation, the tissues were washed in water, dehydrated in ethanol, clarified in xylene and embedded in paraffin (Histosec, Merck, Darmstadt, Germany). Tissue sections (thickness 5 μm) were subjected to immunohistochemistry for the detection of the androgen receptor (AR), as described in protocols applied to the prostate, estrogen receptor alpha (ERΑ), p63 protein, α-actin and proliferating cell nuclear antigen (PCNA). Primary antibodies reactive to AR (rabbit polyclonal IgG, N-20; Santa Cruz Biotechnology, Santa Cruz, CA, USA), ER-α (rabbit polyclonal IgG, MC-20; Santa Cruz Biotechnology), p63 (mouse monoclonal IgG2a, sc-843, 4A4; Santa Cruz Biotechnology), α-actin (mouse monoclonal IgG2a, sc-32251, IA4; Santa Cruz Biotechnology) and PCNA (mouse monoclonal IgG2a, sc-56, PC10; Santa Cruz Biotechnology) were employed at a dilution of 1:100. Peroxidase-conjugated specific antibodies (Sigma Chemical Co., St Louis, MO, USA) or polymers (Post Primary Block and polymer; Novocastra, Newcastle Upon Tyne, UK) were used as secondary antibodies for 45 min at 37°e sections were revealed with diaminobenzidine and counterstained with Harris's hematoxylin. The histological sections were then analyzed under an Olympus BX60 light microscope (Olympus).
3D reconstruction
3D reconstruction was made to determine the pattern of prostatic tissue distribution in both male and female gerbils from the C and HT experimental groups. In order to do this, 20 histological sections (3 μf the central region of the prostatic complex were stained with HE (as described above) and analyzed with a Leica M125 stereo-scopy microscope coupled to an AxioCam HRC 10-33 photographic camera (Zeiss-Jenaval, Jena, Germany). The images were digitalized using the software AxioVison Release 4.7.2 (Zeiss) for Windows. The software Reconstruct 25 aligned the urethra, vagina, ducts and prostatic alveoli of histological sections. Finally, an interface link between each section and a 3D model was created.
Statistical analyses
The hypothesis tests employed to determine statistical significance were the Kruskal-Wallis test for non-parametric distributions and analysis of variance for parametric distributions. Further localization of the statistically significant differences between experimental groups was done using Dunn's test for non-parametric distributions and Tukey's test for parametric distributions. The hypothesis test employed for two samples of non-parametric data was Mann-Whitney. The data were analyzed using Statistica 6.0 (StarSoft, Inc., Tulsa, OK, USA) and BioEstat 5.0 (free statistical program) software (Instituto de Desenvolvimento Sustentável Mamiraua, Tefé, Amazonas, Brazil). The level of significance was set at 5% (P ≤ 0.05). Values were presented as mean ± standard error of mean (SEM).
Results
Biometric analyses
Biometric analyses (Table 1) showed no significant changes of body weight in males. The weight of the male prostatic complex decreased in treated animals, reaching the lowest values in the HT group. The difference in the relative weight of prostatic complexes of males in the treated groups (LT and HT) was statistically significant to those of the C group.
Biometrical and serological data of male and female gerbils (mean ± SEM*)
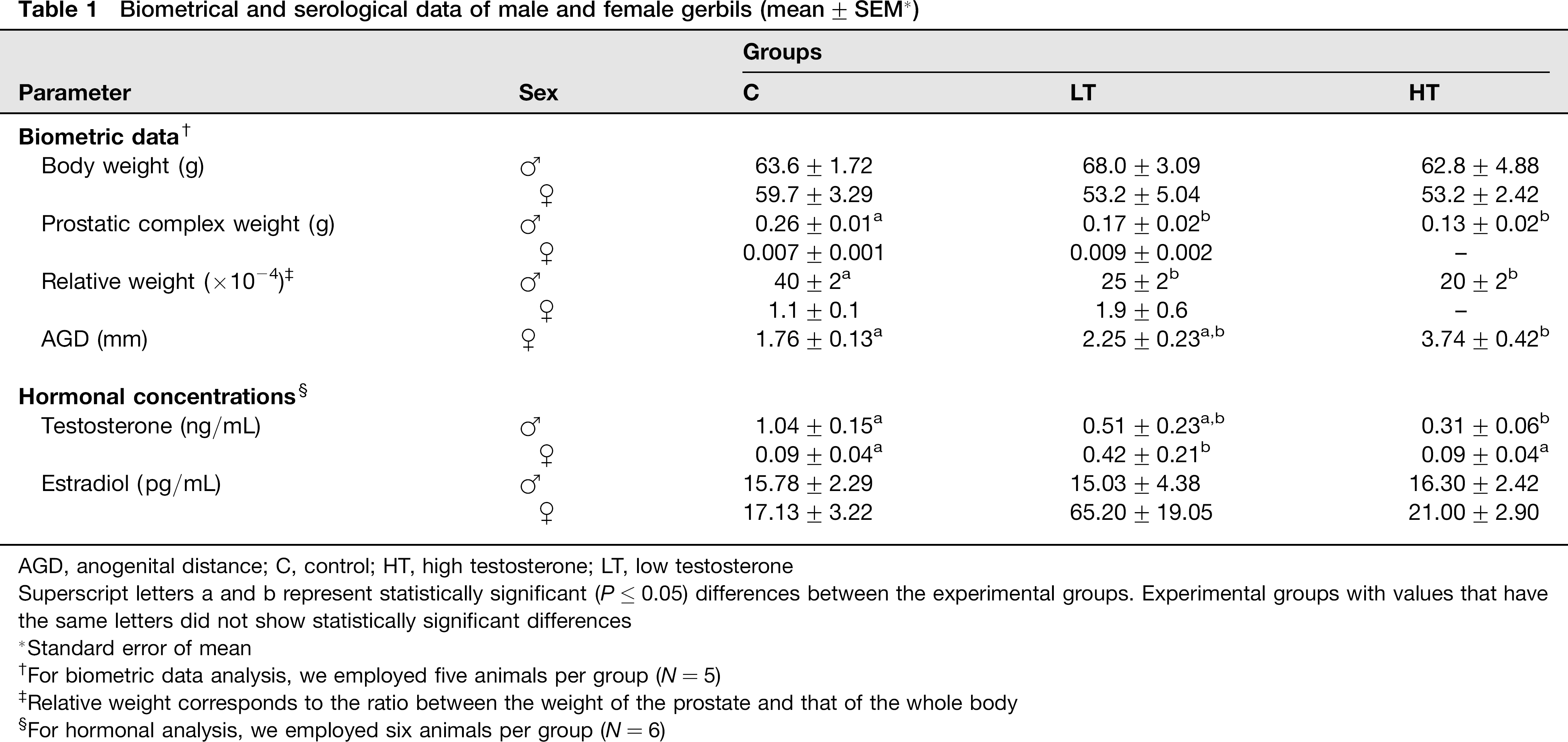
AGD, anogenital distance; C, control; HT, high testosterone; LT, low testosterone
Superscript letters a and b represent statistically significant (P ≤ 0.05) differences between the experimental groups. Experimental groups with values that have the same letters did not show statistically significant differences
Standard error of mean
For biometric data analysis, we employed five animals per group (N = 5)
Relative weight corresponds to the ratio between the weight of the prostate and that of the whole body
For hormonal analysis, we employed six animals per group (N = 6)
Biometric analyses of body weight, prostatic complex weight and relative weight did not show any differences among females of the experimental groups (Table 1). The anogenital distance (AGD) showed a statistically significant difference only for females in the HT group. The measurements of the prostatic complex weight and the relative weight of the HT females were not taken because the pro-static tissue showed a scattered distribution around the vaginal wall, impairing a precise isolation of the female pro-static complex (UPT).
Plasma total testosterone and estradiol dosage
The testosterone concentrations showed (Table 1) a decrease for both LT and HT in the male groups in comparison with the control, being statistically significant only for HT in comparison with the C group. However, there was no statistically significant difference between the LT and HT male groups. Regarding the females, only the LT group showed higher testosterone values than the C and HT groups. Hormonal dosage data for estradiol did not show any significant differences for the male or female experimental groups.
Anatomical aspects observed during animal dissection
No microscopic changes were observed in the reproductive system of male gerbils during dissection. However, females of the HT group showed an absence of vaginal opening, increased AGD, and hydrometrocolpos (data not shown).
Morphological characteristics of the male and female prostatic glands
Morphological analyses using HE staining (Figure 2) showed a normal pattern of epithelial and stromal tissue architecture for the prostatic glands of males from the C group (Figures 2a and b). On the other hand, the males of the LT (Figures 2e and f) and HT (Figures 2i and j) groups showed different prostatic changes distributed within the prostatic tissue. The formation of intraepithelial arcs (Figure 2f) and PIN (Figure 2j) were observed, as were inflammatory foci inside the luminal compartment (Figure 2f).
Morphological aspects of male gerbil prostatic tissue stained with hematoxylin–eosin showing general morphological aspects and by Gömöri's reticulin technique and immunohistochemistry reaction showing the stromal changes for the reticular fibers and smooth muscle α-actin, respectively. The hematox-ylin–eosin stain showed a characteristic morphological pattern of the ventral prostates of the C (a, b) and LT (e, f) groups, which were characterized by large acini with abundant secretory materials and a thin stromal compartment. On the other hand, the HT (i, j) group showed a ventral prostate with a reduced lumen and a stromal compartment more developed, besides some foci injured by PIN lesions. In relation to Gömöri's reticulin technique, the C group (c) showed a regular pattern of reticular fiber organization around prostatic acini, while we noticed the presence of regions with intense reshuffling of these fibrillar elements in the LT (g) and HT (k) groups. With the α-actin reaction, it was possible to observe the normal pattern of smooth muscle layer integrity in the C (d) and LT (h) animals and an altered pattern characterized by discontinuous regions of this layer in HT groups (l). Ac, acinus; Ep, epithelium; S, stroma; L, lumen; asterisk, inflammatory cells; large arrows, intraepithelial arc; PIN, prostatic intraepithelial neoplasia; arrows, reticular fiber; arrowhead, smooth muscle α-actin; short arrow, absence of smooth muscle layer; C, control; HT, high testosterone; LT, low testosterone. (A color version of this figure is available in the online journal)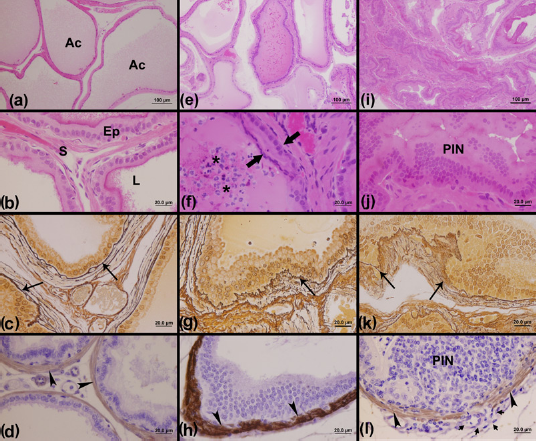
Similarly, the epithelial and stromal prostate components of the female C group showed a normal morphological pattern (Figures 3a and b). However, prostatic changes in females of the LT (Figures 3e and f) and HT (Figures 3i and j) groups were observed, especially in the HT group, which showed the most altered pattern of prostate tissue due to testosterone exposure. The most common prostatic changes observed were hyperplasia of the epithelial compartment (Figure 3f) and PIN (Figure 3j).
Histology of female prostatic tissue stained with hematoxylin–eosin showing general morphological aspects and by Gömöri's reticulin technique and immunohistochemistry reaction showing the stromal changes for the reticular fibers and smooth muscle α-actin, respectively. The hematoxylin–eosin stain showed a characteristic morphological pattern of the prostates of the C (a, b) and LT (e, f) groups, which were characterized by large acini with abundant secretory materials. On the other hand, the HT (i, j) group showed a prostate with a reduced lumen and a more developed stromal compartment when compared with the C and LT groups, besides some foci injured by PIN lesions (j). In relation to Gömöri's reticulin technique, the C group (c) showed a regular pattern of reticular fiber organization around the prostatic acini, while the presence of regions was noticed with intense reshuffling of these fibrillar elements in hyperplasic regions of the LT (g) and HT (k) groups. With the α-actin reaction, it was possible to observe the normal pattern of smooth muscle layer integrity around the acini in the prostates of C (d) and LT (h) animals and an altered pattern characterized by an intense reshuffling of this layer in some foci of the prostate in the HT group (l). Ac, acinus; Ep, epithelium; S, stroma; L, lumen; PIN, prostatic intraepithelial neoplasia; arrows, reticular fiber; arrowhead, smooth muscle α-actin; asterisk, inflammatory cells; C, control; HT, high testosterone; LT, low testosterone. (A color version of this figure is available in the online journal)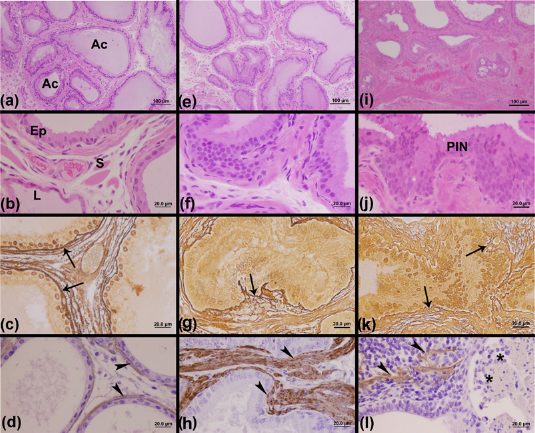
Gömöri's reticulin technique allowed the observation of the normal arrangement of the reticular fibers in the prostates of C males (Figure 2c) and a rearrangement of these elements in some regions of the prostates of the LT (Figure 2g) and HT males (Figure 2k). The immunoreaction for α-actin showed a pattern of distribution for the smooth muscle layer around the prostatic acini of the different experimental groups (Figures 2d, h and l). While the C (Figure 2d) and LT (Figure 2h) groups generally showed a normal pattern of distribution of the smooth muscle layer, in the HT group (Figure 2l) it was possible to observe regions of an interrupted smooth muscle layer surrounding the PIN lesions in the epithelium of HT male prostates.
With regard to females, the C (Figure 3c) and LT (Figure 3g) groups showed a normal pattern of reticular fiber distribution, while the HT females (Figure 3k) showed regions of intense rearrangement of these fibers in the stromal compartment. The immunoreaction for α-actin allowed the observation of a normal pattern of smooth muscle layer distribution in C (Figure 3d) and LT female prostates (Figure 3h) and an intense stromal reshuffle in areas that were injured with prostatic changes in the HT female prostates (Figure 3l).
Stereological data of male and female prostates
Although both the LT and HT male groups showed an increased epithelial compartment, a statistically significant increase was observed only in the prostates (Table 2) of the HT male group in comparison with the C group. The LT male group did not show any statistically significant differences in comparison with the C and HT male groups. The luminal compartment underwent an expressive reduction in animals of the HT male group when compared with the C group, in a manner that was different from the stroma, which increased, although not significantly, reaching the highest stereological value in this group.
Stereological and morphometrical data (mean ± SEM)

C, control; HT, high testosterone; LT, low testosterone
Superscript letters a, b and c represent statistically significant differences (P ≤ 0.05) between the experimental groups. Experimental groups with values that have the same letters did not show statistically significant differences
n = 200 measurements in five animals per group
For females, the main changes occurred in the HT group (Table 2), which showed an increase of the epithelium followed by a significant decrease of the prostatic luminal compartment. Moreover, the most significant changes were the intense increase of the stromal area in the HT group in comparison with the C group.
Morphological aspects of male and female prostates
The prostatic epithelium height of the male LT and HT groups (Table 2) increased significantly. With regards to the smooth muscle layer, only males of the LT group had a mean value of smooth muscle layer thickness that was significantly different from the other groups.
All females of the experimental groups exposed to testosterone (LT and HT) during the embryonic phase showed a significant height increase of the prostatic epithelium. The highest values of smooth muscle layer thickness were observed in females of the LT and HT groups.
3D reconstruction
3D reconstruction allowed the pattern of prostatic tissue distribution over the reproductive tract to be observed in both males and females of the C and HT groups (Figure 4). Males of two groups (C and HT) presented a characteristic pattern of prostatic distribution on the ventral portion of the urethra (Figures 4a and b). However, the male HT group showed a prostatic gland with acini that were generally more reduced than those found in the prostates of the C group. The females of the C group showed a characteristic pattern of the prostatic tissue in a paraurethral position (Figure 4c). In females of the HT group, although the prostate had also presented with a typical paraurethral localization, it was possible to observe an atopic distribution of prostatic tissue around the vaginal wall (Figure 4d), which is completely abnormal in wild animals.
Three-dimensional reconstruction of the prostate from C (control) and HT (high testosterone) male and female groups. Panels a and b show a typical pattern of ventral prostatic tissue distribution in males. Panel b shows a decrease in the acini diameter in the ventral prostate of the HT male group. In the females of the C group (c), it was possible to observe a typical pattern of paraurethral localization of the prostatic tissue, while in the females of the HT group (d), an abnormal distribution of prostatic tissue around the vaginal wall was observed. Structures in red indicate prostatic tissue; yellow, urethra; blue, urethral lumen; light gray, vagina – showed only in (d). Ca, caudal; Cr, cranial. Inset represents the approximate angle (45°) of the horizontal rotation of the images. (A color version of this figure is available in the online journal)
Immunohistochemistry (AR, ERΑ, p63 and PCNA)
AR
The immunohistochemistry assay for the AR showed a similar pattern of nuclear reaction in the male prostatic tissue of all of the analyzed experimental groups: C (Figure 5a), LT (Figure 5e) and HT (Figure 5i). Besides the secretory epithelial cells, stromal cells, such as fibroblasts, and smooth muscle cells were found to be positive for AR.
Imunohistochemistry for the androgen receptor (AR), p63, estrogen receptor alpha (ERΑ) and proliferating cell nuclear antigen (PCNA) of the male ventral prostate. Immunohistochemical reaction for the AR showed the pattern of nuclear marking for ARs in epithelial and stromal cells for all of the analyzed groups: C (control) (a), LT (low testosterone) (e) and HT (high testosterone) (i). Panels b and f show the immunohistochemical reaction for p63, which is characterized by a distribution of basal cells in the basal layer of the prostatic epithelium of the C and LT groups, respectively. On the other hand, this technique showed some foci of altered expression for p63 in the HT group (j). In relation to immunohistochemical reaction for the ERΑ, rare stromal cells positive to ERΑ were observed in the stromal compartment of the LT group (g) and an absence of immunoreactivity in the prostate of group C (c). However, the presence of this receptor was observed in both epithelium and stromal compartments in the prostate of the HT group (k). The immunohistochemical reaction for PCNA allowed the observation of few proliferating cells in the epithelium of the C (d) and LT (h) groups, especially in the prostates of the HT group (l). Short arrow, stromal cells positive to AR; arrows, cells with positive marking for p63; PIN, prostatic intraepithelial neoplasia; arrowhead, cells with positive marking for ERΑ; wide arrows, epithelial cells marked for PCNA. (A color version of this figure is available in the online journal)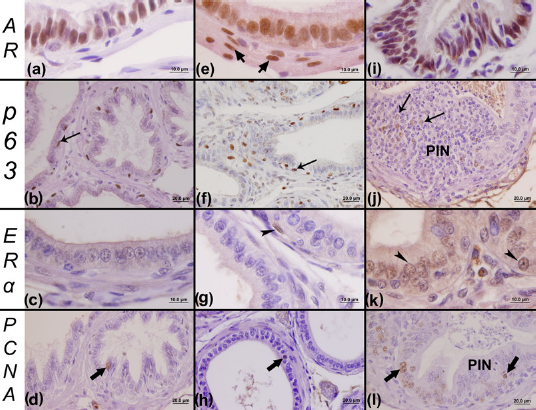
The AR reaction for all females also had a similar pattern of immunoreactivity for groups C (Figure 6a), LT (Figure 6e) and HT (Figure 6i). Moreover, there were few stromal cells or smooth muscle cells with positive reactions for AR (Figure 6e).
Imunohistochemistry for the androgen receptor (AR), p63, estrogen receptor alpha (ERΑ) and proliferating cell nuclear antigen (PCNA) of the female prostates. Positive reactions for the AR showed the pattern of nuclear marking for ARs in the epithelial compartment of all analyzed groups: C (control) (a), LT (low testosterone) (e) and HT (high testosterone) (i). Some rare stromal cells that were positive to AR could also be visualized (e) in the female prostate. Panels b and f show immunohistochemical reactions for p63 for the C and LT groups, respectively, which were characterized by a typical distribution of basal cells adjacent to the secretory cells. Moreover, this technique showed some foci of altered expression for p63 in the HT group (j), which was characterized by a multilayer pattern of the basal cells. In relation to the immunohistochemical reaction for the ERΑ, epithelial and stromal cells positive to ERΑ were observed in the prostate of all groups: C (c), LT (g) and HT (k). However, a more intense reaction of this receptor was observed in the epithelium of the prostates of the HT group (k), especially in hyperplasic or regions injured by PIN lesions. The immunohistochemical reaction for PCNA allows the observation of proliferating cells in the epithelial of all of the analyzed female groups, especially in the prostates of the HT group (l), which showed several lesion foci in comparison with the C (d) and LT (h) groups. Short arrow, stromal cells positive to AR; arrows, basal cells; arrowhead, cells with positive marking for ERΑ; PIN, prostatic intraepithelial neoplasia; wide arrows, epithelial cells marked for PCNA. (A color version of this figure is available in the online journal)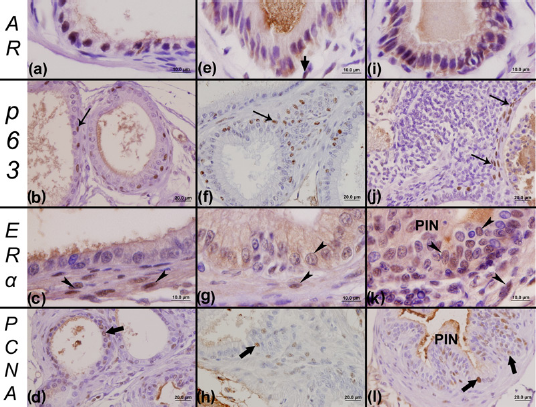
p63
A similar pattern of immunoreactivity was found for basal cell markers (p63) in both C and LT male prostates (Figures 5b and f). However, areas of prostatic lesions in the prostates of the HT male group were characterized by abnormal immunolocalization of p63 (Figure 5j). With regard to females, basal cells were normally disposed in the prostates of the C (Figure 6b) and LT (Figure 6f) groups. However, in the prostates of the HT group, we noticed some abnormal aspects as well as an increase in basal cells in regions with inflammatory foci (Figure 6j).
ERΑ
We observed a stromal reaction for ERΑ only for the LT (Figure 5g) and HT (Figure 5k) male groups. A few prostatic stromal cells, such as fibroblasts, and smooth muscle cells with a positive reaction for ERΑ (Figures 5g and k) were identified in males. However, an intense epithelial reaction was observed for ERΑ in the prostates of the HT male group in hyperplasic regions (Figure 5k), which was not observed for the C (Figure 5c) and LT (Figure 5g) groups.
On the other hand, there was a higher immunoreaction for ERΑ, in terms of visualized positive cells, in both the stroma and epithelium of prostatic tissue in all of the female groups (Figures 6c, g and k) in comparison with the male groups. Also, the identification of stromal cells and epithelial secretory cells marked for ERΑ was more frequent in the female prostatic tissue than in the male tissue. Moreover, an intense epithelial and stromal reaction was observed for ERΑ in the prostates of the HT female group (Figure 6k) in regions with lesions, such as PIN (Figure 6k), that are characteristically frequently found in this group yet are not commonly found in the C (Figure 6c) and LT (Figure 6g) groups.
PCNA
A similar pattern of immunolocalization for PCNA was found in both males in the C (Figure 5d), LT (Figure 5h) and HT (Figure 5l) groups, and in females in the C (Figure 6d), LT (Figure 6h) and HT (Figure 6l) groups. However, in the prostate of males (Figure 5l) and females (Figure 6l) of the HT group, a high number of proliferating cells were found in regions with PIN, which were commonly found in the animals of this group, especially in females.
Discussion
The present study showed that intrauterine exposure to testosterone cypionate alters the morphophysiology of the pro-static gland of adult male and female Mongolian gerbils, leading to a pathological state characterized by a more hyperplasic epithelium and by the presence of lesions such as PIN. Furthermore, the excessive testosterone exposure severely affected females of the HT group, such that these animals developed an increased AGD, an absence of vaginal opening, the atopic formation of injured prostatic tissue around the vaginal wall and hydro-metrocolpos, which is defined as a distension of the vagina and uterus, either with mucous content, produced by uterine and cervical glands, or by the accumulation of urine in the presence of a vaginal obstruction. 26
Our results suggest that the significant decrease in the prostatic complex weight in males of the LT and HT groups may have occurred in response to the interference of exogenous testosterone during intrauterine life. However, further studies are necessary to reveal the mechanisms involved in the changes of overall prostate weight.
The most significant changes occurred in the females of the HT group, which showed several prostatic lesions and masculinized characteristics, due to abnormal testosterone exposure during intrauterine life. The more evident changes were the increased AGD, the appearance of pro-static tissue around the vaginal wall, the development of hydrometrocolpos and PIN lesions. A previous study showed that the exposure of Sprague–Dawley female rats to testosterone propionate during development (14–19 days of gestation) led to the appearance of hydrometrocolpos. 21 Some of these changes had already been described in other studies4,10,21,27 conducted with other rodents, especially rats. However, few studies evaluated the late impacts of abnormal androgenic exposure during the intrauterine phase on the physiology of the prostate. Moreover, no reports are available regarding the effects of the EDCs with androgenic potential on the development of the female prostate (Skene's gland) and its effects during adult or senile life.
A 3D reconstruction showed that females exposed to a high testosterone dosage during intrauterine life (HT group) presented an abnormal pattern of prostatic tissue localization around the vaginal wall. These results reinforce the essential role of androgens during prostatic organogenesis, influencing the pattern of formation and the branching of prostatic buds, as demonstrated by other studies with rodents.28,29 On the other hand, although the males did not show atopic prostatic tissue, as observed in HT females, the HT male group presented prostates characterized by a reduced acini diameter.
The severe increase of the AGD in females of the HT group was dose-dependent in relation to testosterone. Studies carried out with Sprague–Dawley rats showed that newborn females exposed to a dosage of 1 mg/day of testosterone propionate during the 14–19 gestational days had an increased AGD, whereas the males did not have any changes, even when exposed to a higher concentration of this androgen. 21 Studies of female Wistar rats also demonstrated that fetal exposure to androgens promoted an increase in AGD. 30 However, those authors adopted a different methodology, in which the androgens were administered on a daily basis for a prolonged period, while the current study showed that a single dose of 1 mg was sufficient to promote an increase in the AGD in females of the HT group.
A dose-dependent response could be explained when the placenta is seen as a barrier. Studies of Sprague–Dawley rats showed that the placenta may block the passage or even metabolize androgens in other steroids, modulating the concentration of fetal hormones. 28 One of these events is the aromatization of testosterone (by p450 or aromatase enzymes) into 17-b estradiol, which makes the determination of the exact concentration of testosterone that would affect the fetuses difficult. 30
Stromal changes were studied with Gömöri's reticulin and immunohistochemistry for smooth muscle alpha-actin. The main morphological changes were recorded in the prostate of both males and females of the HT group in contrast to the C and LT groups. We found regions with damaged reticular fibers adjacent to the epithelium. Smooth muscle alpha-actin immunohistochemistry showed areas with a discontinued smooth muscle layer, which is characteristic of regions with intense stromal remodeling due to cellular proliferation and the infiltration of inflammatory cells at adjacent stroma.
A significant increase in the prostatic epithelial height was found in both males and females exposed to testosterone during gestation. These aspects suggest that prostatic cells were affected by the abnormal exposure to testosterone during embryogenesis. However, the mechanisms involved in this androgenic imprint are still unknown, making more studies necessary to reveal the activated pathways during this process.
Generally, the prostates of males and females of the HT groups showed an increase in the area corresponding to epithelium, a decrease in the luminal volume and an increase in the stromal compartment. The most significant aspect was the increase in the stromal area in females of the HT group, who presented prostate glands composed mainly of stromal tissue. Besides the stereology, morphometric analyses also showed a more developed stromal compartment, characterized by the increase of the smooth muscle layer in females of the LT and HT groups. Recent studies have suggested that a differential trait of BPH is the transition of epithelial cells to mesenchymal cells and later its differentiation into myofibroblasts, resulting in a significant increase of the glandular stromal area. 31 In relation to this, our data suggest that the exposure to high testosterone dosage during the prenatal period may lead to the development of BPH in the female animals, which were the most affected by the androgens.
Regarding the AR, both male and female groups had a similar pattern of nuclear reaction. Although both male and female prostates showed stromal reactivity to AR, the presence of positive cells in the prostates of females was rare, while the males presented an intense positivity to AR in the stromal compartments.
A differentiated pattern of immunolocalization was found for p63 in the prostate of males of the HT group, which was characterized by the loss of the basal cell layer in regions with PIN. In the prostates of females of the HT group, regions with an increase of basal cells were observed near inflammatory foci. These findings are in agreement with data from the literature. According to Grisanzio and Signoretti, 32 there is a progressive loss of the basal cell layer in high-grade PIN, causing a differential expression of p63. The use of p63 has been an important tool in pro-static cancer diagnostic, taking into account the fact that this protein regulates the differentiation, development, proliferation and apoptosis of prostatic epithelial cells. However, the role of p63 in tumorigenesis is not well known, making new approaches to understand the mechanisms of the p63 regulation necessary. 32
The immunohistochemistry data for ERΑ showed that there was a higher immunoreaction, in terms of the positive cells visualized, for ERΑ in both the stroma and epithelium of prostatic tissue of all of the female groups in comparison with the male groups. Moreover, a stronger ERΑ reactivity was observed in regions of the prostate affected by PIN, both in males and females of the HT group. In these cases, an intense positivity of epithelial cells to ERΑ was seen, which was not common in the prostate of the C and LT groups. These results are in agreement and reinforce the role of ERΑ in promoting proliferation and inflammation, in addition to being an inductor of premalignant lesions. Recent studies have compared the different roles of the estrogenic receptors ERΑ and ERΒ in the prostate.31–35 According to the literature, the activation of the ERΑ pathway involves some processes, as well as aberrant cellular proliferation, inflammation and the development of premalignant lesions, while the ERΒ pathway is involved in critical functions, stroma–epithelial signalization and in the mediation of antiproliferative effects of the estrogens on the prostatic epithelium. 33
Besides p63 and ERΑ, PCNA is also used to study the pattern of cellular proliferation in investigations with therapeutic intervention. 36 An increase in proliferative cells was seen, especially in regions affected by PIN, in both males and females of the HT group, confirming the presence of proliferative lesions in these cases. Conversely, in other experimental groups, either for males or females, few proliferating cells were marked for PCNA in the prostate.
It is believed that abnormal situations during prostatic organogenesis, as well as exposition to EDCs and exogenous hormones, polycystic ovaries and hyperandrogenism may cause epigenetic changes in the pattern of prostate development, favoring the development of premalignant neoplastic lesions during adult life.
Considering the employment of exogenous testosterone as a model of endocrine disruption, it is concluded that the action of EDCs with androgenic potential is dosage-dependent, as mentioned in other studies with androgens.21,28 This fact was observed throughout the experiment, with the most significant changes being more evident in the HT group. Furthermore, this paper showed that the prostates of females, mainly in the HT group, were more affected, since androgens had a masculinizing effect during the development of the reproductive system of these animals. In relation to prostatic lesions, such as PIN, although the incidence of these lesions in glands has not been assessed by a quantitative method, the evidence clearly suggests that exposure to testosterone during the prenatal phase increases the susceptibility of the prostate developing these lesions, especially in females.
This differential response to intrauterine androgenic exposure in females deserves attention, especially when monitoring the prostate. According to Santos and Taboga, 15 80% of female gerbils have a prostate gland, a frequency that is very similar to that found in women. 20 These aspects support gerbils as an excellent model for studies of the prostatic gland. Moreover, it is important to highlight the fact that the typical hormonal environment in females simulates approximately what happens in men during senescence, which is a moment in which the estrogen/testosterone ratio falls drastically. This period of hormonal imbalance coincides, more frequently, with the beginning of prostatic lesions, such as BPH and PCa.
The concern about the exposure to EDCs has grown since the employment of these substances has increased over the years due to the development of society. Accordingly, humans are exposed to a great variety of these chemical agents, with either estrogenic6–8 or androgenic4,27 potential. The trenbolone acetate, for example, is a synthetic androgen that is often used to improve cattle growth. Furthermore, this compound is widely employed in papermills, being responsible for the pollution of effluents of rivers. 4 Besides exposure to EDCs, there are other risk factors that can affect the development of the prostate, such as polycystic ovaries, 37 hyperandrogenism and the abuse of steroids by athletes and women seeking hormonal replacement.
All of these aspects are of great concern, taking into account the fact that the development of the reproductive system is composed of events that are finely regulated during embryogenesis, and small interferences are sufficient to cause permanent damage that will be carried by the organism throughout the entire life.
Recent studies have reinforced the importance of the embryonic period as a critical moment of development of the prostatic gland. According to Schaeffer et al., 9 PCa may originate from deregulated cellular programs that are activated during embryogenesis. It is well known that the AR pathway regulates the expression of more than a hundred genes in the normal prostate and also in lineages of cancerous prostatic cells. 11 The initial developmental program of the prostate is very similar to the program that controls the development of prostatic cancer, which is involved in the regulation of genes related to the acid phosphatase and Wnt pathways, as well as other processes, such as angiogenesis, apoptosis, migration and cellular proliferation. 9
Considering that little is known about the origin of the PCa, studies involving EDCs with androgenic action can contribute to the understanding of mechanisms that regulate the AR pathway. This may highlight how this nuclear receptor, which is essential during prostatic embryogenesis, behaves in altered hormonal environments and what this may mean for prostate health during the life.
This study shows that an endocrine-disrupting chemical with androgenic action may act as a dangerous substance for the health, especially during critical ontogenetic periods of life, such as the embryonic phase. The presence of serious malformations in adult animals, such as hydro-metrocolpos, increased AGD and even the development of premalignant prostatic lesions, such as PIN, show the potential that these substances may have for health. All of these lines of evidence should be considered since the variety and quantity of these substances in our environment has been increasing.
Although the present study has shown some limitations regarding the methodology in manipulating gerbils during the prenatal phase, which have already been reported in scientific literature,38–40 the present work allows us to develop new and more precise procedures in manipulating this model for future studies involving prostate morphogenesis. Therefore, these experiences demonstrate that gerbils are an excellent model for these approaches, which opens several perspectives for future research focusing on the influence of a disrupted prenatal environment and its relationship with the development and aging of the prostate gland.
Footnotes
Acknowledgements
The authors thank Luiz Roberto Falleiros Júnior as well as other researchers of the Laboratory of Microscopy and Microanalysis for their technical assistance.
This paper was supported by grants from the Brazilian agency FAPESP (São Paulo Research Foundation, Procs. No. 2007/06862-3) and CNPq (Brazilian National Research and Development Council, Procs. No. 301111/05-7, 473626/2010-1, 300163/2008-8 to SRT).