
Abstract
Endometriosis, with a prevalence rate ranging from 6% to 10%, is the major contributor to pelvic pain and subfertility, and considerably reduces the quality of life in affected women. However, the pathogenesis of this disease remains largely unknown. The present study aimed to uncover the role of hyperperistalsis in the pathogenesis of endometriosis, by exploring the response of human endometrial stromal cells (ESCs) to the cyclic stretch in vitro. ESCs isolated from 18 different endometrium biopsies undergoing hysterectomy for myoma were subjected to uniaxial cyclic stretches with different magnitude and frequency using the Uniaxial Tension System. Expression of cyclooxygenase-2 (COX-2) and microsomal prostaglandin E2 synthase-1 (mPGES-1) in stretched and unstretched ESCs were assessed by realtime quantitative polymerase chain reaction and Western blot. Production of prostaglandin E2 (PGE2) in the culture medium was measured by enzyme-linked immunosorbent assay. The cyclic stretch mimicking hyperperistalsis in endometriosis (5% elongation at 4 cycles/min) stimulated quick up-regulations of COX-2 and mPGES-1 simultaneously on both transcriptional and translational levels, and delayed PGE2 overproduction was also noted in ESCs. As the stretch magnitude or frequency increased, so did overexpression of COX-2 and PGE2 (P < 0.05). By contrast, the cyclic stretch mimicking physiological peristalsis (3% elongation at 2 cycles/min) did not induce significant COX-2, mPGES-1 or PGE2production within 12 h. Both COX-2 and mPEGS-1 are PGE2 synthases, and the aberrant COX-2 and PGE2 production play important roles in the pathogenesis of endometriosis. Therefore, the present findings revealed that increased stretch stimuli from the hyperperistalsis of endometriosis were capable of causing the aberrant COX-2 and PGE2 expression in the endometrium by mechanotransduction, in a magnitude and frequency-dependent manner. It implied possible roles of hyperperistalsis in the pathogenesis of endometriosis, particularly in the view of COX-2 and PGE2.
Introduction
Endometriosis, defined as endometrial-like tissue outside the uterus, is a major contributor to pelvic pain and subfer-tility. Affecting 6–10% of women of reproductive age, 1 it considerably reduces the quality of life in affected women to a great extent. 2 However, most patients of endometriosis receive symptomatic treatments with a rather high recurrence rates, 3 because of the unclear pathogenesis.
Numerous theories have been proposed to elaborate the pathogenesis of endometriosis, but none of them have produced a complete explanation.4–9 Among these theories, the role of the eutopic endometrium has recently gained more attentions. 10 Based on the aberrant eutopic endome-trium theory, some researchers have suggested that the subendometrial myometrium might also be involved,11,12 because the subendometrial myometrium and the endome-trium are not only anatomically consecutive, but also of identical origin and are functionally associated.13–15 On account of the fact that hyperperistalsis and dysperistalsis of the subendometrial myometrium have presented in endo-metriosis patients throughout the menstrual cycle,16,17 it is apparent that the endometrial cells of the patients suffer more mechanical stretch stimuli than that of healthy women. However, whether and how the hyperperistalsis or increased mechanical stimulus is involved in the patho-genesis of endometriosis is still the fundamental issue, and to our knowledge, little evidence is yet available.
Given that cells can transduce mechanical signals into a cascade of biochemical signals, 18 we hypothesized that the hyperperistalsis of subendometrial myometrium might lead to the dysfunctional eutopic endometrium by mechanotransduction and contribute to the pathogenesis of endometriosis. The present study investigated the effect of uniaxial cyclic stretch on human endometrial stromal cells (ESCs), using an in vitro model mimicking physiological peristalsis (healthy women) and hyperperistalsis (endo-metriosis patients). In addition, we determined whether this mechanical stretch-mediated response was magnitude-or frequency-dependent. Cyclooxygenase-2 (COX-2) and microsomal prostaglandin E2 synthase-1 (mPGES-1) and prostaglandin E2 (PGE2) were chosen as indicators to test the hypothesis, because all of them have been reported to play crucial roles in the pathogenesis of endometriosis.19–22 COX-2 and mPGES-1 are PGE2 synthases, which can promote the production of PGE2. 23 Particularly, dysfunctional expression of COX-2 in the eutopic endometrium of endometriosis has been widely reported.20,24,25 By demonstrating the role of hyperperistalsis in the pathogenesis of endometriosis, the present study may broaden the understanding of the disease.
Materials and methods
Patients and endometrial tissue samples
The study was approved by the Ethics Committee of Peking Union Medical Collage Hospital (PUMCH), and signed informed consent was obtained from each woman. Eighteen women of reproductive age (20–45 y old) undergoing hysterectomy at the Department of Obstetrics and Gynecology in PUMCH between 1 October 2010 and 31 May 2012 were recruited. Considering that dysfunctional subendometrial peristalsis was more noticeable in the secretary phase of endometriosis, 26 we only included patients confirmed to be in the pathological secretory phase. Eligibility criteria required individuals to be diagnosed as intramural or subserous myoma (submucous myoma was excluded) by excluding endometriosis or adenomyosis with laparoscopy and pathology. Patients diagnosed with endometrial disease, such as endometrial carcinoma (or atypical hyperplasia), endometrial polyps and others, were excluded. None had received an intrauterine device, hormone therapy or gonadotropin-releasing hormone analog treatment within three months before the surgery. The endometrial tissue was collected under sterile condition during the surgery. Then, it was placed in a sterile tube containing Dulbecco's modified Eagle's medium (DMEM) supplemented with 100 U/mL penicillin, 100 mg/mL streptomycin and 2.5 μg/mL amphotericin B, and transported on ice to a laboratory within eight hours for the isolation and primary culture of ESCs.
Isolation and culture of ESCs
ESCs were isolated and cultured using the protocol reported by Ryan et al. 27 with minor modifications. Briefly, after rinsing in phosphate-buffered saline (PBS), endometrial tissue was minced into small pieces and digested in DMEM containing type III collagenase (1 mg/mL, 200 U/mg; Worthington Biochemical Corporation, Lakewood, NJ, USA) and deoxyribonuclease I (0.1 mg/mL, 2000 U/mg; Worthington Biochemical Corporation) for 60 min at 37°C with agitation. The dispersed ESCs were separated through a 40-μm sterile cell strainer, and then harvested in a 100-mm culture plate with DMEM containing 10% fetal bovine serum (FBS), 4 mmol/L L-glutamine, 100 U/mL penicillin, 100 mg/mL streptomycin and 2.5 μg/mL amphotericin B in humidified 5% CO2/95% air environment at 37°C. The medium was changed every other day, and cells were passaged following treatment with 0.05% Trypsin-ethylenediaminetetraacetic acid solution. All the ESCs used were between passage 1 and 3 in the study. All the regents used in the isolation and culture were purchased from Invitrogen Corporation (Grand Island, NY, USA).
Application of cyclic tensile strain using the Uniaxial Tension System
ESCs cultured on compliant membranes were subject to cyclic uniaxial tensile strain by the custom-built Uniaxial Tension System (patent number: 200810111785.4, China), which modified the stretch machine reported by Kurpinski et al. Beforehand, ESCs were harvested in a sterile silicone membrane (11.3 cm × 2.1 cm × 0.01 cm; Specialty Manufacturing Inc., Saginaw, MI, USA) coated with type I collagen (Sigma, St Louis, MO, USA) at a plating density of 2 × 105 cells per membrane. Cell-seeded membranes were cultured in DMEM with 10% FBS until 90% confluence and changed to serum-free medium at 12 h before application of cyclic strain. Afterwards, the membranes were clamped into grippers that were entirely placed inside a tank filled with serum-free culture medium. The mobile gripper was connected with a stepper motor, which was precisely manipulated by the computer. Therefore, the membranes were cyclic-stretched at an even pace with defined magnitude and frequency. ESCs were treated with four degrees of magnitude (3%, 5%, 8% and 10%) and frequency (2, 4, 6 and 8 cycles/min) to model the physiological peristalsis and hyperperistalsis (endometriosis) of the subendometrial myometrium according to the previous studies.16,17,29–31 Controls for all the stretch experiments consisted of ESCs seeded on membranes and treated identically, but not subject to cyclic stretch. All the experiments were carried out in a temperature-controlled incubator with humidified 5% CO2/95% air at 37°C.
Cell viability
Before and after the stretch, ESC viability was assessed by trypan blue exclusion test. In brief, 200 μL of 0.4% trypan blue (Sigma) was added to the membrane. The percentage of viable ESCs (cells with unstained nuclei) was calculated by counting 100 cells in five random fields of view using a light microscope (TH4–200; Olympus, Tokyo, Japan).
RNA extraction, reverse transcription and realtime quantitative polymerase chain reaction
Total RNA was extracted from each membrane at the indicated hour using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. The quality and quantity of RNA was identified by NanoDrop 2000 (Thermo Fisher Scientific Inc., Wilmington, DE, USA). Aliquots (600 ng) of RNA were used for cDNA synthesis using the SuperScript® III First-Strand Synthesis system (Invitrogen). The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), COX-2 and mPGES-1 were determined by realtime quantitative polymerase chain reaction (PCR) using Applied Biosystems (Foster City, CA, USA) according to the manufacturer's instructions and a previous study. 32 In detail, the probes and primers were used as follows: TaqMan assay on-demand gene expression primer/probe sets for COX-2 (assay ID Hs00153133_m1), mPGES-1 (assay ID Hs00610420_m1) and the primer/ probe set for GAPDH (human GAPDH endogenous control [VIC/TAMRA probe, Primer Limited; Applied Biosystems]). Realtime quantitative PCR was performed under the following conditions: 95°C for 20 s, 40 cycles of 95°C for three seconds, and 60°C for 30 s. Cycle threshold (Ct) values and the results were determined by ABI Sequence Detection System (Software SDS version 1.3.1; Applied Biosystems). All mRNA Ct values for each sample were normalized by the Ct value of GAPDH in the same sample as follows: ΔCt (sample) = Ct (sample) - Ct (GAPDH). The relative mRNA level was expressed as the value of 2 - ΔCt (sample).
Western blot analysis
ESCs on the membrane were lysed with 100 μL cell lysis buffer containing 0.1 mmol/L phenylmethyl sulfonylfluor-ide (Beyotime Institute Biotechnology, Haimen, Jiangsu, China) and stored at -70°C until further analysis. After concentration testing by a BCA protein assay kit (Pierce, Thermo Fisher Scientific, Rockford, IL, USA), 30 μg total protein for each sample was subject to sodium dodecyl sulfate polyacrylamide gel electrophoresis on a 12% Tris-HCl linear gradient gel with subsequent electroblotting transfer onto poly(vinylidene difluoride) membrane (Millipore, Bedford, MA, USA). The membrane was then blocked with 5% bovine serum albumin for two hours at room temperature, and hybridized overnight with the rabbit anti-human COX-2 (dilution 1:1000; Cell Signaling, Beverly, MA, USA), rabbit anti-human mPGES-1 (dilution 1:250; Cayman Chemical, Ann Arbor, MI, USA) and rabbit anti-human GAPDH (dilution 1:500; Goodhere Biotechnology Corporation, Hangzhou, Zhejiang, China) antibody. It was washed in phosphate-buffered saline with 0.2% Tween 20 before and after incubation with horseradish peroxidase-conjugated goat anti-rabbit IgG (dilution 1:5000; Zhongshan Goldenbridge Biotechnology, Beijing, China), and chemiluminescence was detected with Super Enhanced chemiluminescence detection reagent (Applygen Technologies Inc., Beijing, China) according to the manufacturer's instructions. The density of the bands was analyzed using Quantity One software, version 4.6 (Bio-Rad, Hercules, CA, USA) and all immunoblot data shown are representatives of independent experiments.
Enzyme-linked immunosorbent assay
The concentration of PGE2 in the culture media collected from unstretched and stretched ESCs was measured using a PGE2 enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
Statistical analysis
Parametric data, as determined by the Shapiro–Wilk test, were analyzed using analysis of variance followed by Tukey's test or Dunnett's test for multiple comparisons appropriately with the software package PASW Statistics, version 18.0 (IBM SPSS, Chicago, IL, USA). Spearman correlation was used to reveal the trend when needed. P < 0.05 was considered to be statistically significant.
Results
Cell viability and baseline expression of COX-2, mPGES-1 and PGE2
ESCs isolated from the endometrial tissue remained viable (>90%) throughout the culture period and without change in viability in the following application of cyclic stretch. Low but detectable COX-2, mPGES-1 and PGE2 (<100 pg/mL) baseline expression were determined by Western blot or ELISA method. Though the baseline expressions fluctuate among women, there was no significant difference on the baseline expression of COX-2, mPGES-1 and PGE2 between passage 1 and 3 in ESCs from the same patient (data not shown).
Uniaxial cyclic stretch mimicking the hyperperistalsis in endometriosis induced COX-2, mPGES-1 and PGE2 overexpression in ESCs
At first, we conducted time-course experiments to determine the effect of uniaxial cyclic stretch on COX-2, mPGES-1 and PGE2 expression in ESCs. The cyclic stretch of 3% elongation at 2 cycles/min and 5% elongation at 4 cycles/min were chosen to imitate the physiological peristalsis and hyperperistalsis in endometriosis, respectively, according to the previous publications.16,30,31,33,34
Figure 1 illustrates the stretch-induced COX-2, mPGES-1 and PGE2 expression in ESCs. Applications of cyclic stretch mimicking physiological peristalsis did not induce significant COX-2 expression or PGE2 production within 12 h (Figure 1). However, cyclic stretch mimicking hyperperistalsis in endometriosis stimulated quick up-regulations of gene expression of COX-2 and mPGES-1 simultaneously (Figure 1a and b). Delayed PGE2 overproduction was also found, responding to the cyclic stretch mimicking hyperperistalsis in endometriosis (Figure 1c). Under the cyclic stretch of 5% elongation at 4 cycles/min, the expression of COX-2 mRNA came to the peak at four hours (3.67 ± 0.79 fold, P < 0.05) and the expression of mPGES-1 mRNA came to the peak at eight hours (2.32 ± 0.47 fold, P < 0.05). They smoothly decreased to the platform about two-fold over control at 12h (Figure 1a and b). The protein level of COX-2 and mPGES-1 production was in line with the mRNA expression. Representative results are shown in Figure 1d. Significant increased PGE2 production was noted at four hours, and increased steadily as the stretch time increased within 12 h.
Effect of the uniaxial cyclic stretch on COX-2, mPGES-1 and PGE2 expression in ESCs. ESCs were mechanically unstimulated or stimulated with cyclic stretch of 3% elongation at 2 cycles/min (white) and 5% elongation at 4 cycles/min (black), and then harvested immediately for the indicated hours. Gene expression of COX-2 and mPGES-1 were determined relative to GAPDH and normalized to the unstimulated control by realtime quantitative PCR (a, b). The production of PGE2 was assessed by ELISA (c). Typical examples of COX-2 and mPGES-1 protein production were determined by Western blot (d). Values are the mean ± SEM of samples from four subjects per group. *Statistically significant difference versus unstimulated control. COX-2, cyclooxygenase-2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; mPGES-1, microsomal prostaglandin E2 synthase-1; PGE2, prostaglandin E2; ESC, human endometrial stromal cell; PCR, polymerase chain reaction; ELISA, enzyme-linked immunosorbent assay
After the treatment with cyclic stretch of 5% elongation at 4 cycles/min for four hours, inducible COX-2, mPGES-1 and PGE2 expression were observed (Figure 2). Interestingly, overexpressed COX-2 and mPGES-1 expression persisted within the first few hours at the mRNA level (Figure 2a and b). Persistent PGE2 production was also present within the first four hours (2.28 ± 0.42 fold at four hours, P < 0.05) and then decreased to the baseline level gently at 24 h (Figure 2c).
Recovery of COX-2, mPGES-1 and PGE2 expression in ESCs after cyclic stretch stimuli. After being subjected to cyclic stretch with 5% elongation at 4 cycles/min for four hours, ESCs were immediately harvested or continued to be cultured for indicated hours. Gene expression of COX-2 and mPGES-1 were determined relative to GAPDH and normalized to the unstimulated control by realtime quantitative PCR (a, b). The production of PGE2 was assessed by ELISA (c). Values are the mean ± SEM of samples from four subjects per group. *Statistically significant difference versus unstimulated baseline control. COX-2, cyclooxygenase-2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; mPGES-1, microsomal postaglandin E2 synthase-1; PGE2, prostaglandin E2; ESC, human endometrial stromal cell; PCR, polymerase chain reaction; ELISA, enzyme-linked immunosorbent assay
Up-regulation of COX-2, mPGES-1 and PGE2 by the uniaxial cyclic stretch were frequency-dependent in ESCs
Second, we investigated whetherthe response tocyclic stretch was frequency-dependent in ESCs. Cyclic stretches of 5% elongation with a gradient frequency from 2 cycles/min to 8 cycles/min were performed on ESCs for four hours. DynamicCOX-2, mPGES-1andPGE2expression as a function of frequency are shown in Figure 3. Only at the frequency higher than 4 cycles/min were significant COX-2, mPGES-1 and PGE2 inductions presented (Figure 3). Furthermore, there was a significant trend that up-regulation of COX-2, mPGES-1 and PGE2 expression was frequency-dependent (Spearman correlation, P < 0.05).
Effect of frequency on COX-2, mPGES-1 and PGE2 expression in ESCs. ESCs were mechanically unstimulated or stimulated with cyclic stretch of 5% elongation at gratitude frequency (2, 4, 6 and 8 cycles/min) for four hours. Protein production of COX-2 and mPGES-1 were determined relative to GAPDH and normalized to the unstimulated control by Western blot analysis, and subsequently quantified by densitometry using Quantity One software (a). The production of PGE2 was assessed by ELISA (b). Values are the mean ± SEM of samples from six subjects per group. ‘Statistically significant difference versus unstimulated control. COX-2, cyclooxygenase-2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; mPGES-1, microsomal postaglandin E2 synthase-1; PGE2, prostaglandin E2; ESC, human endometrial stromal cell; ELISA, enzyme-linked immunosorbent assay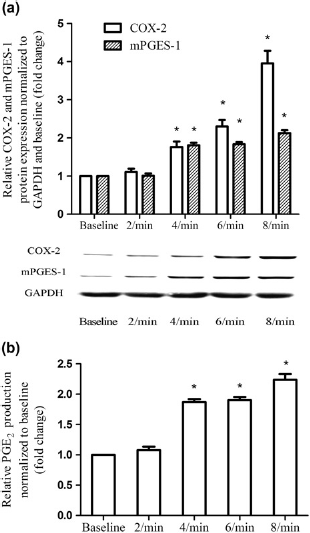
Up-regulation of COX-2, mPGES-1 and PGE2 by the uniaxial cyclic stretch were magnitude-dependent in ESCs
Besides the observed frequency-dependence to the cyclic stretch, we also examined the effect of stretching magnitude in ESCs (Figure 4). Significant increased COX-2, mPGES-1 and PGE2 production were noted with increased stretch magnitude from 3% to 10% elongation at the frequency of 4 cycles/min for four hours in ESCs (Spearman correlation, P < 0.05). More than eight-fold of COX-2 protein (Figure 4a) expression was noted with 10% elongation stretch.
Effect of magnitude on COX-2, mPGES-1 and PGE2 expression in ESCs. ESCs were mechanically unstimulated or stimulated with cyclic stretch of gratitude magnitude (3% elongation, 5% elongation, 8% elongation and 10% elongation) at 4 cycles/min for four hours. Protein production of COX-2 and mPGES-1 were determined relative to GAPDH and normalized to the unstimulated control by Western blot analysis, and subsequently quantified by densitometry using Quantity One software (a). The production of PGE2 was assessed by ELISA (b). Values are the mean ± SEM of samples from six subjects per group. *Statistically significant difference versus unstimulated control. COX-2, cyclooxygenase-2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; mPGES-1, microsomal postaglandin E2 synthase-1; PGE2, prostaglandin E2; ESC, human endometrial stromal cell; ELISA, enzyme-linked immunosorbent assay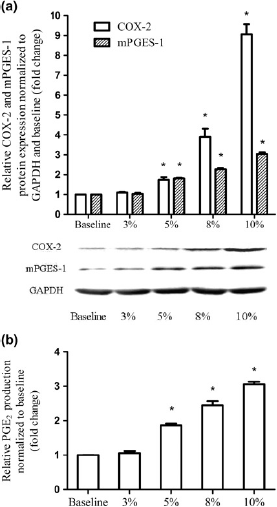
Discussion
In the present study, we demonstrated that increased stretch stimuli mimicking the hyperperistalsis of endometriosis induced COX-2, mPGES-1 and PGE2 overexpression in ESCs in vitro. In addition, the stretch-induced COX-2 and PGE2 production were magnitude- and frequency-dependent with persistent effects. By contrast, the stretch stimuli mimicking the peristalsis of healthy women did not stimulate significant COX-2, mPGES-1 and PGE2 overexpression. These findings implied that hyperperistalsis in endometriosis is capable of causing dysfunction in the endometrium by mechanotransduction and may be involved in the pathogenesis of endometriosis.
Because it is quite difficult to study the effect of the stretch-mediated cell regulation in vivo, we investigated the mechanical stretch response of ESCs using an in vitro model. Compared with the previous studies using a Flexcell system (Flexcell International Co., Hillsborough, NC, USA) to provide the equiaxial stretch,30,31 we used the uniaxial system with mild modification according to the machine reported by Kurpinski et al., 28 which provided uniaxial or anisotropic strain. It provided mechanical stimuli more similar to in vivo conditions of subendometrial myo-metrial peristalsis, and was more sensitive to uncover the magnitude- and frequency-dependent nature of the stretch-mediated cell response.28,35
COX-2 and PGE2 play important roles in increased proliferation, migration and invasion of the ESCs in endo-metriosis, and promote favorable implantation of endometrial cells outside the uterus by perpetuating a positive feedback loop that leads to local inflammation and hyperestrogen.19–21 COX-2 was widely reported to be over-expressed in the eutopic endometrium of endometriosis patients.20,24,25 As a PGE2 synthase, mPEGS-1 is preferentially coupled with the inducible COX-2 to promote PGE2 generation, and also has been reported to be involved in the pathogenesis of endometriosis. 22,23 In the present study, quick and persistent overexpression of COX-2, mPGES-1 and PGE2 were induced by endometriosis-like stretch with a frequency- and magnitude-dependent manner in ESCs, but not by physiological stretch. It indicated that hyper-peristalsis of endometriosis may cause the aberrant augmentation of COX-2 and PGE2 in the endometrium by mechanotransduction. Because COX-2, mPGES-1 and PGE2 have been reported to be mechanosensitive genes in bladder smooth cells, vascular endothelia, fetal lung epithelia, fibroblasts, cartilage explants and others, the findings also corroborated and broadened the previous studies.
Interestingly, after the stretch withdrawal, the gene expression of COX-2 and mPGES-1 did not fall immediately, but continued to be overexpressed in a few hours. Persistent PGE2 was also noted. It suggested that the cyclic mechanical stretch had a persistent effect on the regulation of COX-2 and PGE2. With the unusual inherent background and hyperestrogen status in the endometrium of endometriosis in vivo,19,41 once the ESCs suffered mechanical stimuli which can induce COX-2 or PGE2 overexpression, a cascade of steroids and inflammatory factors will be subsequently activated, and dysfunction of the endometrium may advance stepwise.
As to the molecular mechanisms by which cells sense and response to the stretch stimuli, we are still largely in the dark, though several pathways or models have been hypothesized.42–46 Zhao et al., 37 Mohan et al. 47 and Amma et al. 39 reported the stretch-induced COX-2/PGE2 up-regulation mediated by activation of the nuclear factor-κB pathway in vascular endothelial cells, amnion cells and fibroblasts, respectively. In addition, the Wnt pathway and mitogen-activated protein kinase pathway were also mentioned to produce stretch-induced COX-2/PGE2 in osteoblastic cells, lung epithelial cells and podocytes.48–50 Interestingly, all these pathways were also confirmed to be involved in the dysfunctional eutopic endometrium of endometriosis.51–53 This overlapping plausibly implied the possible roles of stretch stimuli in the pathogenesis of the abnormality in eutopic endometrium and endometriosis. As a first step, we did not explore the mechanotransduction pathways involved in the present study. Future studies with delicate design are needed to explore these complicate interactions.
Subendometrial myometrial peristalsis is one of the origins of the uterine movement in non-pregnant women, 54 which takes part in the process of reproduction, such as the transport of sperm and excretion of shed endo-metrium. 55 Some researchers have suggested that hyperperistalsis and dysperistalsis may be involved in the pathogenesis of uterus-related diseases, such as infertility and endometriosis.12,30 In previous publications, hyperperistalsis was reported to be involved in the pathogenesis of endometriosis by physical processes, such as facilitating retrograde of the fragments of basal endometrium into the peritoneal cavity56,57 and interfering with sperm transpor-tation. 58 To our knowledge, Harada et al. 30 reported the phenomena of mechanotransduction in ESCs for the first time in 2005. However, based on the previous studies, the present study revealed the magnitude- and frequency-dependent manner of mechanotransduction in ESCs, which gave an explanation that the hyperperistalsis may induce pathological biochemical signals in the endometrial cells and participate in the pathogenesis of endometriosis subsequently. In addition, from the view of embryonic origin, the subendometrial myometrium is different from the outer myometrium, but the same as the endome-trium.13–15 It constitutes the archimetra with the endome-trium together, which is functionally conservative during development. 17 Our findings also confirmed the link between the subendometrial myometrium and the endome-trium by mechanotransduction. It suggested that endometriosis might be an archimetra disease including both the endometrium and the subendometrial myometrium.
Moreover, the hypothesis that the subendometrial myo-metrium participated in the pathogenesis of endometriosis might provide a potential clinical guide. Previous theory that the abnormal eutopic endometrium plays a decisive role in the pathogenesis of endometriosis promotes studies of endometrial markers for microinvasive diagnosis and direct treatment.59–62 Similarly, showing the important role of dysfunctional peristalsis of the subendometrial myo-metrium in endometriosis might promote non-invasive diagnosis and evaluation in future. Besides, the movements of the subendometrial myometrium can be non-invasively recorded by vaginal sonography and magnetic resonance imaging.17,26,29,63 It could be useful for the diagnosis and follow-up of endometriosis in future, for example using vaginal sonography to evaluate the direction of the suben-dometrial peristalsis has been reported to be effective in predicting the outcome of in vitro fertilization. 58 More attention should be paid to evaluating the frequency and magnitude of the subendometrial myometrial peristalsis for a better diagnosis, evaluation and follow-up of endometriosis.
Conclusions
Though limited by the in vitro nature of the loading culture system, the present study showed that hyperperistalsis was capable of causing the dysfunction of endometrium in a magnitude- and frequency-dependent manner, particular in view of the pathological inducible COX-2 and PGE2 production. It supported the hypothesis that the subendo-metrial myometrium took part in the pathogenesis of endo-metriosis, which broadened the understanding of endometriosis and may provide new methods for diagnosis and evaluation of the disease.
Footnotes
Acknowledgements
The authors thank Meili Liu, PhD, Wei Song, PhD, Xiaoling Jia, PhD, Lisha Zheng, PhD, and Yan Huang, PhD, from the Key Laboratory for Biomechanics and Mechanobiology of Ministry of Education in China, for their positive suggestions during the experiment. We thank Honghui Shi, MD, and Hua Yang, PhD, from the Department of Obstetrics and Gynecology, Peking Union Medical College Hospital, for their help in patient enrollment and tissue collection. We also thank Dr Edward C Mignot, Shandong University, for linguistic advice. The authors are grateful for the support of the National Natural Science Foundation of China (no. 30950020).