
Abstract
Systemic administration of Salmonella to tumor-bearing mice leads to the preferential accumulation within tumor sites and retardation of tumor growth. The cancer stem-like cell (CSC) hypothesis suggests that CSCs are the root of cancer and induce metastasis and recurrence. The objective of this study was to examine if Salmonella could inhibit the growth of CSCs derived from mouse breast cancer. Systemically injected Salmonella preferentially accumulated within tumors for at least three weeks and the bacteria accumulated preferentially not only in subcutaneous but also in orthotopic tumors over livers and spleens at ratios ranging from 1000:1 to 10,000:1. Salmonella were capable of delaying tumor growth and enhancing survival in both subcutaneous and orthotopic tumor models. More strikingly, Salmonella acted to retard tumor growth and extensively prolong the survival time of the mice bearing CSC-induced tumors. Our results also found that Salmonella predominantly, although not exclusively, resided in the CSC regions of the tumor. These data suggest that Salmonella can inhibit the growth of breast cancer by targeting the CSC niche. In conclusion, Salmonella can be used for the management of breast cancer.
Introduction
Increasing evidence reveals that tumors contain heterogeneous populations including cancer stem-like cells (CSC) and surrounding differentiated cells. CSCs in the tumor and their specific phenotypes may be responsible for their escape from conventional therapies. Recurrence of the tumor may occur because CSCs survive, suggesting that the removal of CSCs is crucial for cancer therapy. CSCs have been identified in numerous tumors, including those of the breast, 1 brain, 2 colon 3 and pancreas. 4 CSCs have been identified in tumor cell lines, based on the expression of surface markers such as CD44, CD133 and stem cell antigen-1 (Sca-1), or by their ability to form spheres. 1 Sca-1 has been recognized as the CSC marker in the 4T1 mouse model of breast cancer. 5 Herein, we identified a CSC population characterized by the Sca-1 phenotype.
Aggressive tumors often have an insufficient blood supply, partly because tumor cells grow faster than the endothelial cells that make up the blood vessels, and partly because the newly formed vascular supply is disorganized. 6 This results in acidity and nutrient deprivation, as well as regions with reduced oxygen concentrations. The hypoxic condition is necessary to maintain the ability of CSCs. 7 It is proposed that CSCs prosper within the hypoxic region of the tumor. Hence, hypoxia is considered a major hindrance to therapy. As hypoxia is a common characteristic of human tumors, which adversely affects the prognosis of patients with cancer, targeting hypoxia may be an effective means of improving cancer treatment. Some anaerobic and facultative anaerobic bacteria have been used experimentally as anticancer agents because of their selective growth in the hypoxic/necrotic regions of solid tumors after systemic administration. Bacteria can actively move away from the vasculature and penetrate into the hypoxic regions of tumors. We see bacterial therapy as the most innovative and flexible method to target the CSC niche of tumors.
Materials and methods
Bacteria, cells and mice
A vaccine strain of Salmonella enterica serovar choleraesuis (S. Choleraesuis) (ATCC 15480) was obtained from Bioresources Collection and Research Center (Hsinchu, Taiwan). This rough variant of S. Choleraesuis (S.C.), designated vaccine 51, was obtained by spreading an 18-h broth culture of the virulent strain 188 of S. Choleraesuis strain Dublin over the surface of a dried nutrient agar plate and placing a drop of a suspension of Salmonella anti-O phage no. 1, and selecting for a phage-resistant colony after incubation at 37°C for 24 h.
8–10
Murine 4T1 breast tumor cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 50 μg/mL gentamicin, 2 mmol/L
Animal studies
The mice were inoculated subcutaneously or orthotopically with 106 tumor cells. When the tumors had grown to 50 mm3 to 100 mm3, the mice were injected intravenously with Salmonella (2 × 106 colony-forming units [cfu]). At various time points post infection, groups of mice were sacrificed, and the numbers of Salmonella in the tumors, livers and spleens were determined on Luria–Bertani (LB) agar plates and expressed as cfu per gram of tissues. The mice were inoculated orthotopically with 103 tumor cells harvested from Sca-1+ sorted 4T1 cells. When the tumors had grown to 50 to 100 mm3, the mice were injected intravenously with Salmonella (2 × 106 cfu). The numbers of Salmonella in the tumors, livers and spleens were determined as previously described. In a separate experiment, palpable tumors were measured every three days in two perpendicular axes with a tissue caliper and the tumor volume was calculated as: (length of tumor) × (width of tumor)2 × 0.45, and the survival of the mice in the treated and control groups was monitored daily. 10
Fluorescence-activated cell sorting analysis of Sca-1+ cells within 4T1 cell line and 4T1-derived tumors
Allophycocyanin-conjugated anti-Sca-1 antibody and Sca-1− or Sca-1+ cells were sorted by a fluorescence-activated cell sorting (FACS) cell sorter. The percentage of Sca-1+ cells in parental 4T1 cells and the purity of sorted Sca-1− or Sca-1+ 4T1 cells were analyzed by WinMDI software (The Scripps Research Institute, La, Jolla, CA, USA). Tumor cells (4T1; 1 × 105) were inoculated subcutaneously in a group of three mice for two weeks. The tumors were harvested and digested with Collagenase/Hyaluronidase solution (StemCell Technologies, Vancouver, Canada) at 37°C for one hour. The dissociated tumor cells were obtained by a 40 μm cell strainer (BD Biosciences, Franklin Lakes, NJ, USA) and stained with antilineage marker antibodies (anti-CD4, anti-CD8a, anti-CD19, anti-NK1.1, anti-F4/80 and anti-Ly6G/C, all conjugated with R-phycoerythrin fluorochrome and purchased from BD Biosciences) in combination with anti-Sca-1-FITC antibody (BioLegend, San Diego, CA, USA) on ice for 30 min. The percentage of Sca-1+ cells was analyzed with Epics XL flow cytometry (Beckman Coulter, Inc., Indianapolis, IN, USA) calculated from lineage tumor cells with WinMDI software.
Tumor spheroid assay
Cells were harvested from Sca-1− or Sca-1+ sorted 4T1 cells and prepared as density of 1 × 104 cells/mL in DMEM/F12 medium containing 0.5% methylcellulose (Sigma-Aldrich, St Louis, MO, USA), 0.4% bovine serum albumin (Invitrogen, Carlsbad, CA, USA), 10 ng/mL epidermal growth factor (Peprotech, Rocky Hill, NJ, USA), 10 ng/mL basic fibroblast growth factor (Peprotech), 5 μg/mL insulin (Sigma-Aldrich), 1 μmol/L hydrocortisone (Sigma-Aldrich) and 4 μg/mL heparin (Sigma-Aldrich). Two milliliters of cell suspension was seeded into wells of an ultralow attachment six-well-plate (Corning, Lowell, MA, USA) and incubated for seven days. Images were captured with inverted microscopy.
Cell migration assay
Cell migration assay was conducted using the Oris Universal Cell Migration Assembly kit (Platypus Technologies, LCC, Madison, WI, USA) following the manufacturer's protocol. Briefly, 5 × 104 cells/100 μL/well were loaded into stopper-loaded wells and incubated overnight to permit cell attachment. To start cell migration, the stoppers was removed, the wells were washed gently with phosphate-buffered saline (PBS), complete cell culture medium was added and incubated for 16h. Pictures of wells were captured with inverted microscopy after fixation and staining with 0.5% crystal violet/50% EtOH. Cell migration area was analyzed with ImageJ software (
Immunofluorescence staining
BALB/c mice were inoculated orthotopically with 106 4T1 cells at day 0. The mice were injected intravenously with 2 × 106 cfu of Salmonella at day 10. Tumor-bearing mice were injected with the specific hypoxic cell marker Hypoxyprobe-1 (60 mg/kg; Chemicon, Temecula, CA, USA) and sacrificed 30 min later. We examined the locations of Salmonella, CSCs and hypoxic regions within 4T1 tumors at day 14. Tumors were excised and snap-frozen. Cryostat sections were prepared, fixed in 10% formalin, permeabilized with cold acetone, incubated with rabbit anti-Salmonella serum (1:100), mouse anti-Ly-6A/E (Sca-1) antibody (1:50; Clone D7; BioLegend), and anti-Hypoxyprobe-1 antibody (1:50; Clone 4.3.11.3; Chemicon) at room temperature for 40 min, and subsequently incubated with Texas Red-conjugated anti-mouse IgG (1:100; KPL, Guildford, UK, USA) and fluorescein-conjugated goat anti-rabbit IgG (1:100; KPL) at room temperature for one hour. Nuclei were stained with 50 μg/mL of 4′,6-diamidino-2-phenylindole (DAPI). The stained tissues were examined under a fluorescence microscope at a magnification of ×400. Terminal deoxynucleotidyltransferase-mediated deoxyuridine triphosphate nick end-labeling (TUNEL) assay was used to detect cell apoptosis within tumors and was performed according to the manufacturer's instructions (Promega, Madison, WI, USA). Nuclei were stained with 50 μg/mL of DAPI. TUNEL-positive cells were counted under the microscope. We counted three high-power (×200) fields that showed the highest density of positive-stained cells per field to determine the average percentage of apoptotic (TUNEL-positive) cells in each section. 9
Statistical analysis
The unpaired, two-tailed Student's t-test was used to determine differences between groups. The survival analysis was performed using the Kaplan–Meier survival curve and log-rank test. Any P value less than 0.05 was regarded statistically significant.
Results
Tumor-targeting potential of Salmonella
We monitored kinetics of the bacterial distribution after injection with 2 × 106 cfu of Salmonella into mice bearing syngeneic 4T1 tumors (Figure 1a) implanted subcutaneously. The bacterial amount was much higher in tumors than in livers and spleens of the mice at all the time points examined. Their amounts in the tumors continuously remained at a peak level for at least three weeks after bacterial inoculation, which were approximately three to five orders of magnitude higher than those found in the livers or spleens. Notably, as tumors became larger at later time points examined, total bacterial number within the tumor actually increased with time, indicating that the bacteria not only preferentially accumulated but also amplified within the tumors. By contrast, as early as day 7, Salmonella were undetectable in the blood from all the mice tested (data not shown). To ensure that the tumor-targeting potential of Salmonella was not confined to the subcutaneous site, but rather a general phenomenon, we investigated its tumor-targeting potential in more clinically relevant cancer models. Mice bearing orthotopic breast tumors were inoculated intravenously with Salmonella, and the amounts of Salmonella in the tumors, livers and spleens were determined after bacterial inoculation. A notable amount of Salmonella was found in the orthotopic breast tumor, whereas Salmonella were detected to a much lesser extent in the spleens or healthy livers (Figure 1b). Taken together, Salmonella accumulated in not only subcutaneous but also orthotopic tumors after systemic administration.
Preferential accumulation of Salmonella (S.C.) in the tumors. Mice-bearing (a) subcutaneous or (b) orthotopic tumors were injected intravenously with 2 × 106 cfu of Salmonella at day 0. The amounts of accumulated Salmonella in the tumors, livers and spleens were determined at days 1, 7, 14 and 21. Each value represents mean ± SD from four mice. Data are representative of three independent experiments
Inhibition of tumor growth by Salmonella
Antitumor effects of Salmonella were evaluated in terms of tumor growth and survival in mice bearing subcutaneous or orthotopic 4T1 tumors. Figure 2a shows that tumor growth was significantly retarded in mice treated with Salmonella compared with that in PBS-treated control mice. The mean tumor volume in the Salmonella-treated group was lowered by 40.7% compared with that in the PBS-treated groups. Figure 2b shows that survival of the mice injected with Salmonella was significantly prolonged compared with that injected with PBS. The results obtained from the orthotopic tumor model are given in Figures 2c and d. The mean tumor volume in mice treated with Salmonella was lowered by 64% compared with that treated with PBS (Figure 2c). Moreover, survival of Salmonella-treated mice was significantly prolonged compared with that of PBS-treated mice (Figure 2d). Taken together, Salmonella exerted antitumor effects in different tumor models.
Antitumor effects of Salmonella (S.C.) on mice-bearing tumors. Groups of eight BALB/c mice that had been inoculated (a, b) subcutaneously or (c, d) orthotopically with 4T1 cells (106) at day 0 were treated intravenously with Salmonella (2 × 106 cfu) at day 10. Vehicle control mice received PBS. Tumor volumes (mean ± SEM, n = 8). (b, d) Kaplan–Meier survival curves are shown (n = 8). *P < 0.05; **P < 0.01; ***P < 0.001. Data are representative of two independent experiments. PBS, phosphate-buffered saline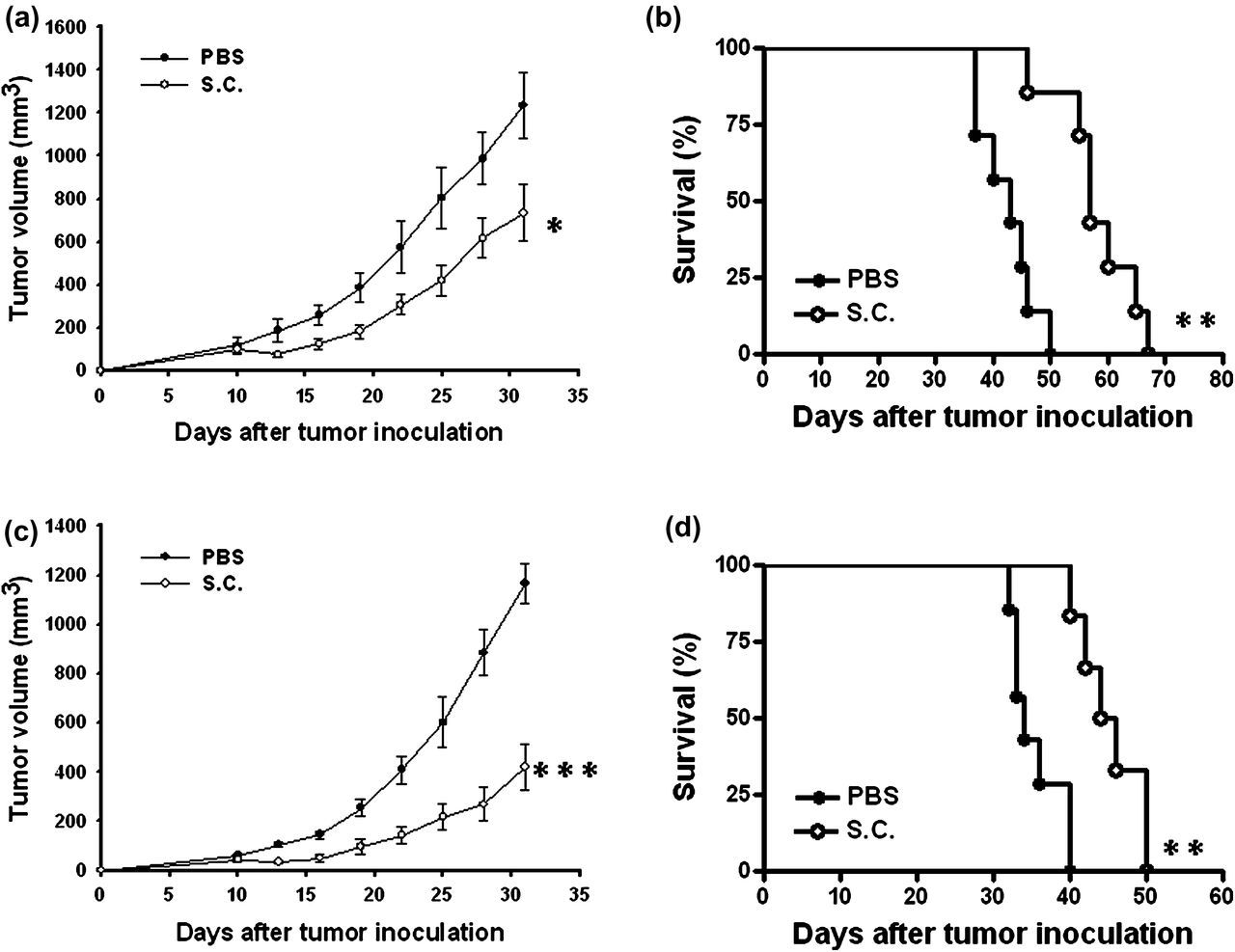
Isolation and characterization of Sca-1+ 4T1 cell population
To investigate if an increase in the Sca-1+ cell population correlates with the malignancy and metastatic ability of 4T1 cells, we examined Sca-1 expression in vitro and in vivo (Figure 3a and b). There were 6.7 ± 0.3% Sca-1+ cells in 4T1 cell lines. Interesting, there were 15.03 ± 4.6% Sca-1+ cells in engrafted tumors. The host may provide some growth factors to increase the Sca-1+ cell population. Furthermore, to examine if 4T1 Sca-1+ cell populations have sphere-forming ability, Sca-1− or Sca-1+ cells were sorted by a FACS cell sorter and examined in a sphere-forming test. We found that the group of Sca-1+ cells had much larger and greater numbers of spheres than the group of Sca-1− cells (Figure 3c). Meanwhile, the Sca-1+ cell population showed significantly the migration ability compared with the Sca-1− cell population (Figure 3d). At the same tumor inoculation dose (1000 cells), the mean tumor volume in the Sca-1− group was lowered by 63.0% compared with that in the Sca-1+ groups at day 60 (Figure 3e). To investigate the tumorigenic ability of the Sca-1+ cells, we sorted 4T1 cells into Sca-1+, and Sca-1− cells and injected them into the mammary glands of mice. As shown in Table 1, the Sca-1+ cells formed tumors in five out of five mice but Sca-1− cells only formed tumors in two out of five mice at day 30. Our results are consistent with previous studies that showed that the Sca-1+ cell population has a more aggressive phenotype than the Sca-1− cell population.
5
Sca-1 as CSC marker for 4T1 murine cells. 4T1 cells were stained with APC-conjugated anti-Sca-1 antibody and Sca-1− or Sca-1+ cells were sorted by FACS cell sorter. (a) The percentage of Sca-1+ cells in parental 4T1 cells and the purity of sorted Sca-1- or Sca-1+ 4T1 cells were analyzed by WinMDI software. (b) BALB/c mice bearing 4T1 tumors ranging from 100 to 200 mm3 were sacrificed, and the Sca-1+ cells in the tumors were determined at day 14. The percentage of Sca-1+ cells within lineage tumor cells was determined by FACS. (c) The cells (2 × 104) were cultured under mammosphere culture condition for seven days. (d) Cell migration capability of Sca-1− or Sca-1+ 4T1 cells was determined (mean ± SD, n = 4). (e) The growth curves of orthotopic Sca-1− or Sca-1+ 4T1 cells in mice at day 70 are shown (mean ± SEM, n = 6). *P < 0.05; ***P < 0.001. Data are representative of two independent experiments. CSC, cancer stem-like cell; APC, allophycocyanin; FACS, fluorescence-activated cell sorting. (A color version of this figure is available in the online journal) Tumor-initiating ability of Sca-1+ cells in mouse model
Bacterial colonization in the CSC niche
To examine if Salmonella targeted the CSC niche in the hypoxic tumor regions, we injected Salmonella into 4T1 tumor-bearing mice, and observed the bacterial distribution, hypoxic regions and CSC niche within tumors. As shown in Figure 4a, Salmonella predominantly resided, not absolutely but very closely, in the CSC niche of the tumor. Meanwhile, the Sca-1+ cells were observed in the hypoxic regions of tumors (Figure 4a). The characteristic of stem cells is their ability to repopulate cell populations in vivo. We sorted 4T1 cells with Sca-1 antibody; and then orthotopically injected 103 cells into the mammary fat pad of mice. All mice injected with Sca-1+ cells developed tumors. We next examined if Salmonella could target breast CSCs in a mouse model. We monitored kinetics of the bacterial distribution after injection with Salmonella into mice bearing CSC-induced tumors (Figure 4b). Similarly, a notable amount of Salmonella was found in the orthotopic breast CSC tumor, whereas Salmonella were detected to a much lesser extent in the spleens or healthy livers. Taken together, Salmonella accumulated in not only differentiated tumors but also in the CSC population after systemic administration.
The colonization of Salmonella (S.C.) in the CSC niche of tumors. (a) Detection of Salmonella in the CSC niche of tumors. 4T1 tumor-bearing mice were injected intravenously with Salmonella (2 × 106 cfu), and at 14 days tumors were removed for immunofluorescence double-staining with anti-Sca-1 antibody by Texas Red to visualize the CSC niche and with anti-Salmonella serum by fluorescein to visualize the bacteria. Nuclei were counterstained with DAPI. The merged column represents the superposition of the tumor sections stained with anti-Sca-1, anti-Salmonella and DAPI to visualize colocalization. Tumors were removed for immunofluorescence double-staining with anti-Sca-1 antibody by Texas Red to visualize the CSC niche and with anti-Hypoxyprobe-1 antibody by fluorescein to visualize the hypoxic regions. Nuclei were counterstained with DAPI. The merged column represents the superposition of the tumor sections stained with anti-Sca-1, anti-Hypoxyprobe-1 and DAPI to visualize colocalization (×400). (b) Preferential accumulation of Salmonella (S.C.) in CSC-induced tumors. Mice-bearing the CSC-induced tumors were injected intravenously with 2 × 106 cfu of Salmonella at day 0. The amounts of accumulated Salmonella in the tumors, livers and spleens were determined at days 1 and 7. Each value represents mean ± SD from four mice. Data are representative of two independent experiments. CSC, cancer stem-like cell; DAPI, 4′,6-diamidino-2-phenylindole. (A color version of this figure is available in the online journal)
Increased apoptotic cells in the CSC-induced tumors following the treatment of Salmonella
The tumorigecic ability of the Sca-1+ cell population was significantly higher than the Sca-1− cell population. We orthotopically injected the Sca-1+ cell population as CSCs to induce tumor growth. To ensure that the antitumor potential by which Salmonella inhibited the stem cell characteristics of breast CSCs derived from 4T1 Sca-1+ cells, we investigated its tumor-targeting potential in CSC-induced tumor models. Treatment of mice bearing CSC-induced tumors with Salmonella significantly retarded tumors growth and prolonged the survival of mice compared with the control group (Figures 5a and b). The tumors from CSC-induced mice treated with Salmonella were analyzed for apoptotic cells by the TUNEL assay. Representative results for the TUNEL assay are shown in Figure 5c. The TUNEL assay showed an increase in the amount of cells undergoing apoptosis in the Salmonella-treated CSC-induced tumors compared with PBS-treated CSC-induced tumors (Figures 5c and d). Taken together, these results indicate that Salmonella resulted in retarding CSC–induced tumor growth, enhancing apoptosis in the CSC-induced tumors.
Antitumor effects of Salmonella (S.C.) on mice bearing CSC-induced tumors. Groups of BALB/c mice that had been inoculated (a) orthotopically with Sca-1+ 4T1 cells (103) at day 0 were treated intravenously with Salmonella (2 × 106 cfu) at day 30. Vehicle control mice received PBS. Tumor volumes (mean ± SEM, n = 7–8). (b) Kaplan-Meier survival curves at day 125 are shown (n = 7–8). (c) Increase in CSC-induced tumor cells undergoing apoptosis in CSC-induced tumor-bearing mice treated with Salmonella (S.C.). Groups of four BALB/c mice that had been inoculated orthotopically with Sca-1+ 4T1 cells (103) at day 0 were treated intravenously with Salmonella (2 × 106 cfu) at day 30. Vehicle control mice received PBS. Tumors were excised at day 40 and (c) TUNEL assay was used to detect apoptotic cells (×200). (d) TUNEL-positive cells were counted from three fields of the highest density of positive-stained cells in each section to determine the percentage of apoptotic cells (mean ± SEM, n = 4). **P < 0.01; ***P < 0.001. Data are representative of three independent experiments. CSC, cancer stem-like cells; PBS, phosphate-buffered saline; TUNEL, terminal deoxynucleotidyltransferase-mediated deoxyuridine triphosphate nick end-labeling; DAPI, 4′,6-diamidino-2-phenylindole. (A color version of this figure is available in the online journal)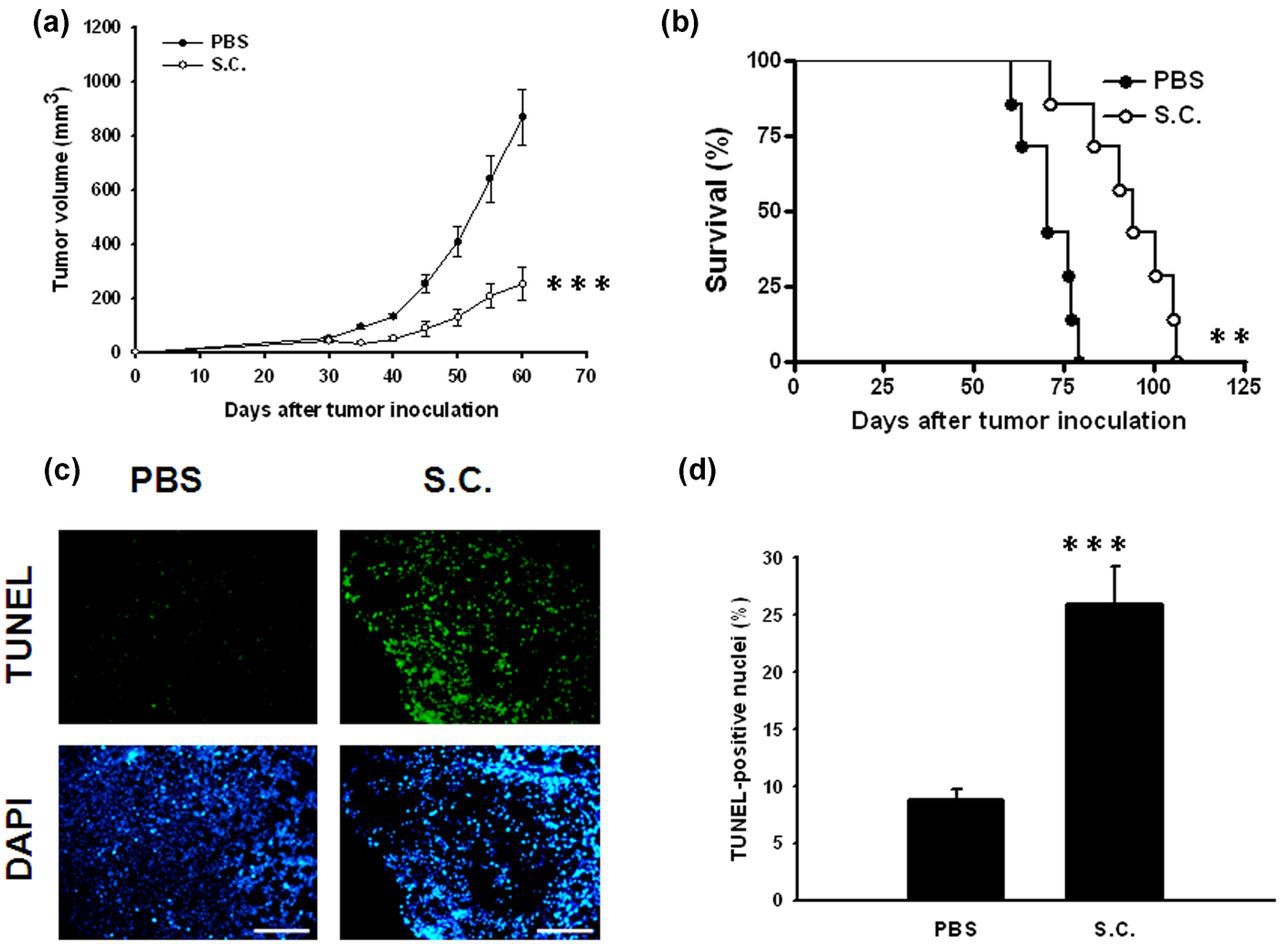
Discussion
Systemic administration of Salmonella to tumor-bearing mice leads to the preferential accumulation within tumor sites and retardation of tumor growth. Salmonella can effectively eradicate primary and metastatic tumors, including bone, prostate, breast pancreas and sarcoma. 11–14 Many studies suggest that Salmonella have the clinical potential for treatment of metastatic tumors. 15–18 It has been shown that the local environment in different organs may affect the properties of tumors growing at that sites. 9 In addition, the activity of some immune cells may be altered by different tumor microenvironments. Orthotopic, rather than subcutaneous tumor transplantation is the method of choice for assaying the defining property of CSCs, tumor propagation. 19 The uniqueness of our study is the use of a murine orthotopic CSC tumor model to investigate the tumoricidal effect of Salmonella. Herein, the mean tumor volume in the subcutaneous tumor model treated with Salmonella was lowered by 40.7% compared with that treated with PBS, but that was lowered by 64% in the orthotopic tumor model. It is conceivable that an orthotopic tumor model is a clinicially more relevant setting than an ectopic subcutaneous tumor model.
The tumor was induced by injection with 106 complete tumor cells in mice. The CSC-induced tumor was initiated by injection with 103 Sca-1+ cells. The complete tumor cell population showed 60–70-fold enrichment compared with the Sca-1+ cell group. The two tumors had different Sca-1+ cell populations, and the survival of mice injected with Sca-1+ cells was longer than that of the mice injected with the complete tumor cells (Figures 2 and 5).
Tumor-targeting bacteria have the ability to induce apoptosis in tumor sites by competing nutrients, stimulating immune cells, antiangiogenesis or inducing apoptotic signal pathways. Herein, we point out that Salmonella could inhibit tumor growth by targeting to the CSC niche. Then, the accumulation of Salmonella in the CSC niche induces the apoptosis of CSCs. Salmonella penetrate to tumors by detecting and chemotaxing towards small molecule gradients of serine, aspartate and ribose. 20 The CSC niche is the environment which is responsible for the maintenance of stem cell properties with growth factors and nutrients. Salmonella require nutrients which may derive from the CSC niche. Furthermore, the hypoxic microenvironment was identified as niche where CSCs are concentrated (Figure 4a). 21 Some studies also demonstrated that the CSC phenotype is promoted by hypoxia. 22,23 Salmonella had been demonstrated to grow in the relatively hypoxic foci within tumors. 9,24 However, further work is warranted to elucidate the underlying mechanism of targeting the CSC niche by Salmonella. Our study provides strong evidence demonstrating that the growth of orthotopic CSC-induced tumors could be significantly inhibited by systemic administration of Salmonella, resulting in prolonged survival in mice. The results have significant clinical implications, suggesting that the tumor-targeted and oncolytic effects of Salmonella appear to hold promise for the treatment of breast cancer. We also provide a basis for future elucidation of the underlying mechanisms of antitumor effects by Salmonella.
Footnotes
ACKNOWLEDGEMENTS
This work was supported by grants from National Science Council (NSC 100-2320-B-039-024) and China Medical University (CMU-99-N2-08 and CMU-100-N2-01).