
Abstract
‘It seems terribly wasteful that, during the release of hormones and neurotransmitters from a cell, the membrane of a vesicle should merge with the plasma membrane to be retrieved for recycling only seconds or minutes later.’ – Erwin Neher, Nature 1993;
Introduction
The unit of life, the cell, is encapsulated within a proteoliposome that precisely regulates the entry and exit of ions and molecules into and out of the cell, essential to maintain the delicate chemical balance to sustain life. Neurotransmitters, hormones or digestive enzymes, synthesized within the cells, are packaged and stored within membrane-bound vesicular compartments, ready for release to the outside on demand. In eukaryotic cells, numerous physiological processes rely on secretion that occurs in every organism, from the simple yeast to humans. The secretion of neurotransmitters at the nerve terminal, for example, to enable neurotransmission, allowing thought, movement and coordination. Following a meal, secretion of digestive enzymes from the exocrine pancreas help to digest food. As a consequence of digestion, elevation of blood glucose triggers insulin release from β-cells of the endocrine pancreas to regulate blood glucose levels. Similarly, exposure to allergens such as pollen or to a parasite, elicits an allergic inflammatory response, stimulating mast cells to secrete histamine and other compounds. These are some of the few examples demonstrating how critical cell secretion is to life, and the severe health consequence that impairement of the process may lead to, such as various neurological, digestive, hormonal or immune disorders, to name a few.
The examination of cells following secretion using electron microscopy (EM) demonstrates increased presence of partially empty vesicles, suggesting that during secretion, only a portion of vesicular contents are expelled from the cell. This could only be possible if vesicles were to temporarily dock and fuse at the cell plasma membrane to expel a fraction of their contents. Since empty vesicles are also generated following cell secretion, this suggests that either some vesicles fully discharge, and/or partially discharged vesicles re-engage at the cell plasma membrane either once or multiple times to fully expel their contents. Illogically however, for nearly half a century, it was believed that during secretion, membrane-bound secretory vesicles completely merge at the cell plasma membrane, resulting in diffusion of intra-vesicular contents to the cell exterior and compensatory retrieval of the excess membrane by endocytosis, failing to explain the generation of partially empty vesicles in cells following secretion. Furthermore, it made little sense to presume that mammalian cells possess an ‘all or none’ mechanism of cell secretion resulting from complete vesicle merger at the cell plasma membrane, when even single-cell organisms have developed specialized and sophisticated secretory machinery such as the secretion apparatus of Toxoplasma gondii, the contractile vacuoles in paramecium, and the different types of elaborate secretory structures found in bacteria. This conundrum was finally resolved using atomic force microscopy (AFM) examination of live cells undergoing secretion. Isolated live pancreatic acinar cells in near physiological buffer, when imaged at greater force using AFM, demonstrate at nanometer resolution, the size and shape of secretory vesicles called zymogen granules (ZGs) lying immediately below the apical plasma membrane of the cell. 1 Within 2.5 min of exposure to a physiological secretory stimulus, the majority of ZGs within the pancreatic acinar cells swell, followed by a decrease in ZG size and no loss of secretory vesicles observed throughout the experiment. 1 These results demonstrated, for the first time, the transient fusion and partial discharge of vesicular contents during cell secretion. They also demonstrated the swelling of intracellular secretory vesicles following stimulation of cell secretion and their deflation following partial discharge of their contents. 1 The other breakthrough in our understanding of cell secretion was also made using AFM, 2–11 and came with the discovery of the porosome – a new cellular structure. Porosome structure–function has been further confirmed using EM. 6–11 Since porosomes in exocrine and neuroendocrine cells measure 100–180 nm, and only a 20–45% increase in porosome diameter is demonstrated following the docking and fusion of 0.2–1.2 μm in diameter secretory vesicles, 2–5 it is concluded that secretory vesicles ‘transiently’ dock and fuse, as opposed to complete vesicle merger at the porosome base, to release intravesicular contents during secretion. If 0.2–1.2 μm in diameter secretory vesicles were to merge at the 100–180 nm porosomes, the porosome structure would be completely obliterated, which is never observed. The further determination of the composition of porosomes, 6,7,9–12 their structure 2–12 and dynamics 2–5 using high-resolution AFM and EM, and their functional reconstitution into artificial lipid membrane, 6,10 have provided, for the first time, a molecular understanding of the secretory process in cells. In agreement with these findings, 1–12 it has been demonstrated that ‘secretory granules are recaptured largely intact following stimulated exocytosis in cultured endocrine cells’; 13 ‘single synaptic vesicles fuse transiently and successively without loss of identity’; 14 and ‘ZG exocytosis is characterized by long fusion pore openings and preservation of vesicle lipid identity’. 15 In recent years, there has been a flurry of research in the field, and a number of papers from several laboratories investigating the porosome in different cell types, both in their native and disease states, such as porosomes in neurons, in sensory hair cells and in RBL-2H3 (rat basophilic leukemia cell line) cells and bone marrow-derived mast cells (BMMCs), have been reported. 16–22 The discovery of the porosome, its structure, function, composition and functional reconstitution into artificial lipid membrane, has resulted in a paradigm shift in our understanding of the secretory process in cells. In this Minireview, a brief background on the discovery of the porosome – the universal secretory portal in cells – and our current understanding of its structure and function is summarized.
Porosome: a universal cellular organelle
Porosomes were first discovered in acinar cells of the exocrine pancreas
2
more than 16 years ago, and subsequently demonstrated to be present in every cell examined.
2–6,16–22
Exocrine pancreatic acinar cells are polarized secretory cells possessing an apical and a basolateral domain. This well-characterized cell synthesizes digestive enzymes, which are packaged and stored within 0.2–1.2 μm in diameter apically located membrane-bound secretory vesicles, called ZGs. It had been well established that following a secretory stimulus, ZGs dock and fuse at the apical plasma membrane to release their contents to the outside. Compared with neurons, which are also polarized cells, and where secretion of neurotransmitters occurs within milliseconds following stimulation, the pancreatic acinar cells secrete digestive enzymes over several minutes after a meal, and therefore this slow secretory cell was ideal, and chosen to study cell secretion. In the mid-1990s, AFM studies were first conducted on isolated live pancreatic acinar cells to evaluate, at high resolution, the structure and dynamics of the apical region of the plasma membrane in both resting and stimulated cells. These studies demonstrated, at the apical plasma membrane, the presence of a group of circular ‘pits’ measuring 0.4–1.2 μm in diameter containing smaller 100–180 nm in diameter ‘depressions’ or pores
2
(Figure 1). Depressions were subsequently demonstrated to be cup-shaped lipoprotein structures where membrane-bound secretory vesicles transiently dock and fuse to release their contents. Hence, the depressions were renamed ‘porosomes’ or secretory portals. Typically three to four porosomes are present within each pit at the apical plasma membrane of pancreatic acinar cells.
2
The basolateral membrane on the contrary, where secretion does not normally occur, is devoid of such pit and porosome structures.
2
High-resolution AFM images of porosomes in live acinar cells following stimulation of secretion further reveal dialation of the porosome opening during secretion, returning to its resting size following completion of the secretory process.
2
Exposure of pancreatic acinar cells to cytochalasin B, a fungal toxin that inhibits actin polymerization and secretion, results in a decrease in porosome size and a consequent loss in secretion.
2
The immunolocalization of secretory proteins at the porosome opening following stimulation of cell secretion has further established porosomes to be the secretory portals in cells.
2–5
As previously stated, studies demonstrate porosomes to be present in all secretory cells examined, including neurons
6
(Figure 2) and astrocytes.
11
In both neurons and astrocytes, porosomes measure 10–15 nm, which is an order of magnitude smaller than porosomes in the exocrine pancreas or in neuroendocrine cells measuring 100–180 nm. Furthermore, neurons possess porosomes that have a central plug for the rapid opening and closing of the structure at the presynaptic membrane for neurotransmitter release.
23
Analogous to pancreatic acinar cells, examination of resting growth hormone secreting cells of the pituitary,
5
chromaffin cells of the adrenal medulla,
4
β-cells of the endocrine pancreas, or mast cells, all demonstrate the presence of porosomes as permanent secretory portals.
24
Porosomes have therefore been recognized as the universal secretory machinery at the plasma membrane in cells (Figures 1 and 2).
Porosomes in acinar cells of the rat exocrine pancreas. (a) Atomic force microscopy (AFM) micrographs of pancreatic porosomes within pits, at the surface of the apical plasma membrane in live pancreatic acinar cells
10
and (b) at the cytosolic compartment of isolated pancreatic plasma membrane preparations.
10
Several circular pits (arrowheads) with porosomes within are seen in the AFM micrograph. AFM micrograph of the cytosolic side of isolated pancreatic plasma membrane preparation depicting a single inside-out pit (arrowheads) containing four inverted cup-shaped porosomes within (arrowhead), associated with a zymogen granule (ZG) to the left in (b). Scale bar in (a) and (b) = 200 nm. (c)–(f) Transmission electron micrograph of a porosome associated with a docked ZG at the apical end of a pancreatic acinar cell.
10
(c) Part of the apical end of a pancreatic acinar cell demonstrating within the bordered square, the presence of a porosome-associated ZG. Scale bar in (c) = 400 nm. The area within the square is magnified to demonstrate the apically located microvilli (MV) and a section through the porosome and the ZG. Note the ZG membrane (ZGM) bilayer is attached directly to the base of the porosome cup. A higher magnification of the porosome in (e) depicts, in greater detail, the porosome bilayer and cross-section through the three protein rings, with the thicker ring present close to the opening of the porosome complex to the outside. The third and the lowest ring away from the porosome opening is seen fused with the ZGM.
10
An outline of the porosome (FP) is shown to demonstrate the continuity of the plasma membrane (PM)-associated porosome facing the cell lumen (L) with the ZGM. (g) Transmission electron micrographs of ZGs, co-isolated with porosomes. Porosomes associated with the surface of ZGs are shown (bar=120 nm; top panel only).
10
At higher magnification, details of the porosome complex demonstrating the presence of separate PM and the ZGM are seen. Note the apically arranged ring complex of the porosome, similar to what is observed in electron micrographs of the structure in intact cells. (h) Schematic diagram depicting pits and porosomes within the apical PM in acinar cells of the exocrine pancreas with docked ZGs, some transiently fused and expelling intravesicular contents to the outside in the process of secretion
2
Porosomes at the presynaptic membrane in synaptosomes from rat brain neurons. (a) Electron micrograph of a single synaptosome full of 40–50 nm synaptic vesicles.
6
Scale bar =100 nm. (b) Atomic force microscopy (AFM) micrograph of an isolated synaptosome from rat brain. Scale bar=100 nm. (c) AFM micrograph of a 10 nm porosome complex at the presynaptic membrane of an isolated synaptosome. Note the central plug and the eight-protein densities.
8
(d) Electron microscopy (EM) micrograph of a 10 nm cup-shaped neuronal porosome with a central plug (arrowhead) and a 50 nm docked synaptic vesicle (SV).
6
(e) AFM micrograph of an inside-out presynaptic membrane prepared from rat brain synaptosomes. Note the larger 40–50 nm SVs docked to the inverted cup-shaped porosome.
6
(f) Schematic drawing depicting a SV docked to the base of a cup-shaped neuronal porosome complex at the presynaptic membrane (Pre-SM) for neurotransmitter release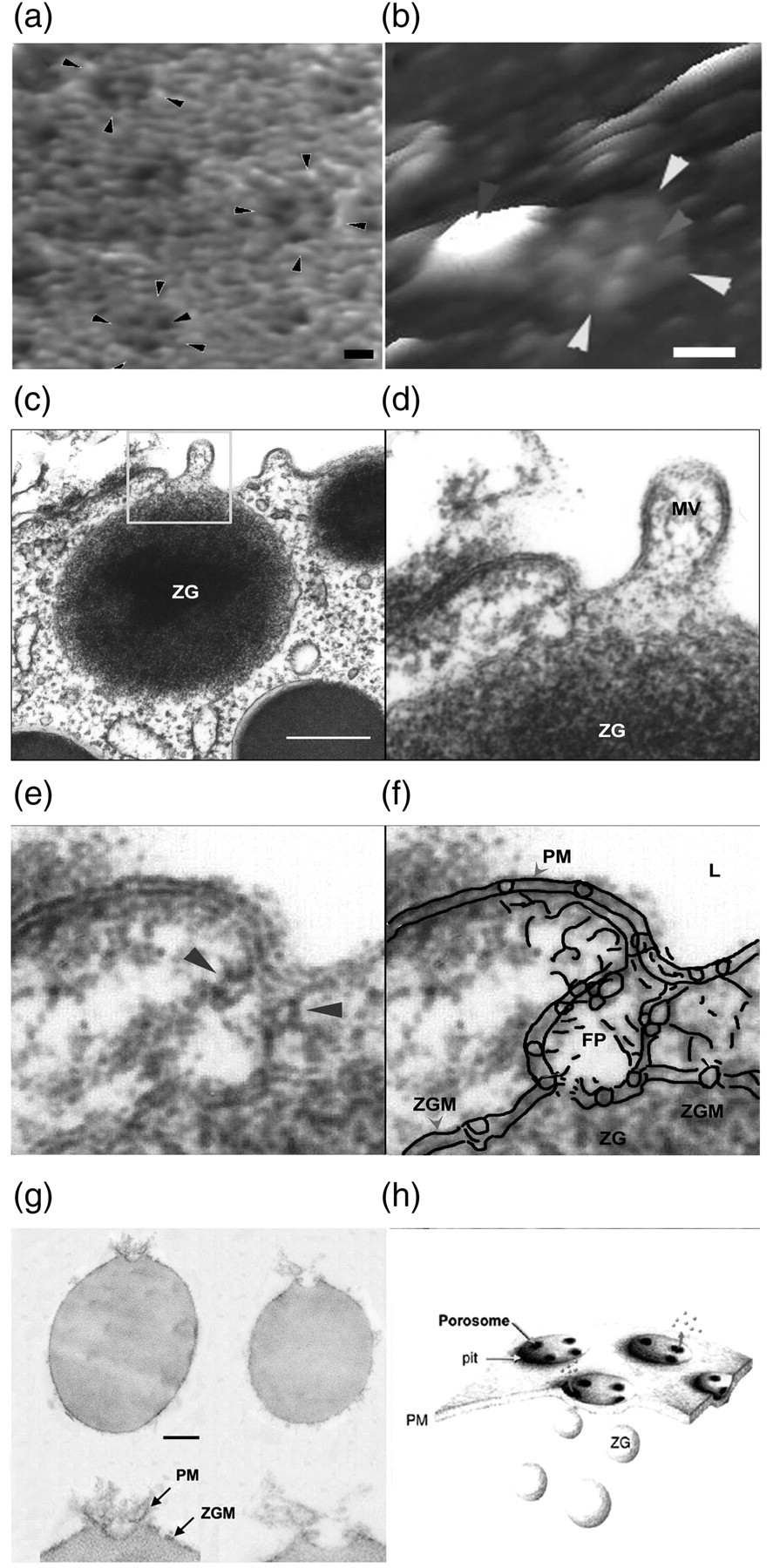

Porosome isolation: using immunoprecipitation and chromatography
The fusion of membrane-bound secretory vesicles at the porosome base is mediated by calcium and a specialized set of three soluble N-ethylmaleimide-sensitive factor (NSF)-attachment protein receptors called SNAREs.
25–29
In neurons, for example, target membrane proteins SNAP-25 and syntaxin called t-SNAREs, present at the base of neuronal porosomes at the presynaptic membrane, and a synaptic vesicle-associated membrane protein (VAMP) or v-SNARE, are part of the conserved protein complex involved in membrane fusion and neurotransmission. It is demonstrated that the t-SNARE protein SNAP-23 is present in acinar cells of the exocrine pancreas and is involved in cell secretion.
30
This suggested that if v-SNARE-associated ZGs were to dock and fuse at the cup-shaped porosomes in pancreatic acinar cells, they would require the t-SNAREs to be present at the porosome base facing the cytosolic compartment of the cell. To test this hypothesis, isolated plasma membrane preparations in near physiological buffered solution have been imaged at ultrahigh resolution using AFM. These studies reveal scattered circular disks measuring 0.5–1 μm in diameter, with inverted cup-shaped structures within
10
(Figure 1b). On several occasion, ZGs ranging in size from 0.4 to 1 μm in diameter have been observed in association with one or more of the inverted cups, suggesting the circular disks to represent pits, and inverted cups the porosomes, in inside-out pancreatic plasma membrane preparations.
10
To further confirm that the cup-shaped structures are indeed where secretory vesicles dock and fuse, immuno-AFM studies have also been carried out using a polyclonal monospecific SNAP-23 antibody that recognizes a single 23 kDa protein in Western blots of pancreatic plasma membrane preparations. These studies demonstrate the selective localization of the SNAP-23 antibody to the base of cup-shaped structures.
10
These results confirm that the inverted cup-shaped structures in inside-out pancreatic plasma membrane preparations are porosomes, where secretory vesicles transiently dock and fuse to release their contents during cell secretion. To establish that porosomes are indeed the secretory portals at the cell plasma membrane, immunogold-labeled secretory proteins have been demonstrated to accumulate at the porosome following stimulation of secretion (Figure 3). Based on these results, porosomes from the acinar cells of the exocrine pancreas have been immunoisolated from detergent-solubilized plasma membrane preparations using SNAP-23 specific antibody.
10
The immunoisolated porosomes have also been used both for reconstitution in lipid membrane for structure–function studies
9,10
(Figures 4 and 5) and for the determination of their composition using immunoblot assays (Figure 6).
Atomic force microscopy (AFM) and immuno-AFM micrographs of the porosome, demonstrating the release of secretory products via the porosome opening to the outside.
9
(a) A pit with four porosomes within, present at the apical in a live pancreatic acinar cell.
9
(b) Following stimulation of secretion, amylase-specific immunogold localize at the porosome opening, demonstrating them to be secretory release sites
9
Isolated and lipid reconstituted porosome complexes from exocrine pancreas and neurons.
6,10
(a) Negatively-stained electron micrograph of an immunoisolated pancreatic porosome complex measuring approximately 150 nm.
10
(b) Atomic force microscopy (AFM) micrograph of an immunoisolated pancreatic porosome complex in buffer.
10
Note the similarity in size and morphology of the structure. Also note the spokes and the three rings, with an approximately 25–30 nm central ring which forms the base of the porosome complex facing the cytosolic compartment, and possibly composed of t-SNAREs and associated proteins. (c) Electron micrographs of immunoisolated pancreatic porosome complex reconstituted into liposomes.
10
Note the cup-shaped porosome structures measuring approximately 150 nm. Scale bar = 100 nm. (d) Negatively-stained electron micrograph of isolated neuronal porosome protein complex measuring approximately 15 nm. Note the complex exhibiting a circular profile and having a central plug. Approximately 8–10 interconnected protein densities are observed at the rim of the structure, which are connected to a central element via spoke-like structures. At the centre of the structure, which corresponds to the porosome base, there is a ring formed most likely of three SNARE pairs.
8,32
(e) Electron density maps of negatively-stained electron micrograph of an isolated neuronal porosome complex. Note the complex exhibiting a circular profile and having a central plug, with eight interconnected protein densities at the rim of the complex, and a central element.
8
(f) An isolated neuronal porosome complex reconstituted in lipid membrane. Note the central plug as well as the eight protein densities at the periphery or lip of the 12 nm structure
6,8
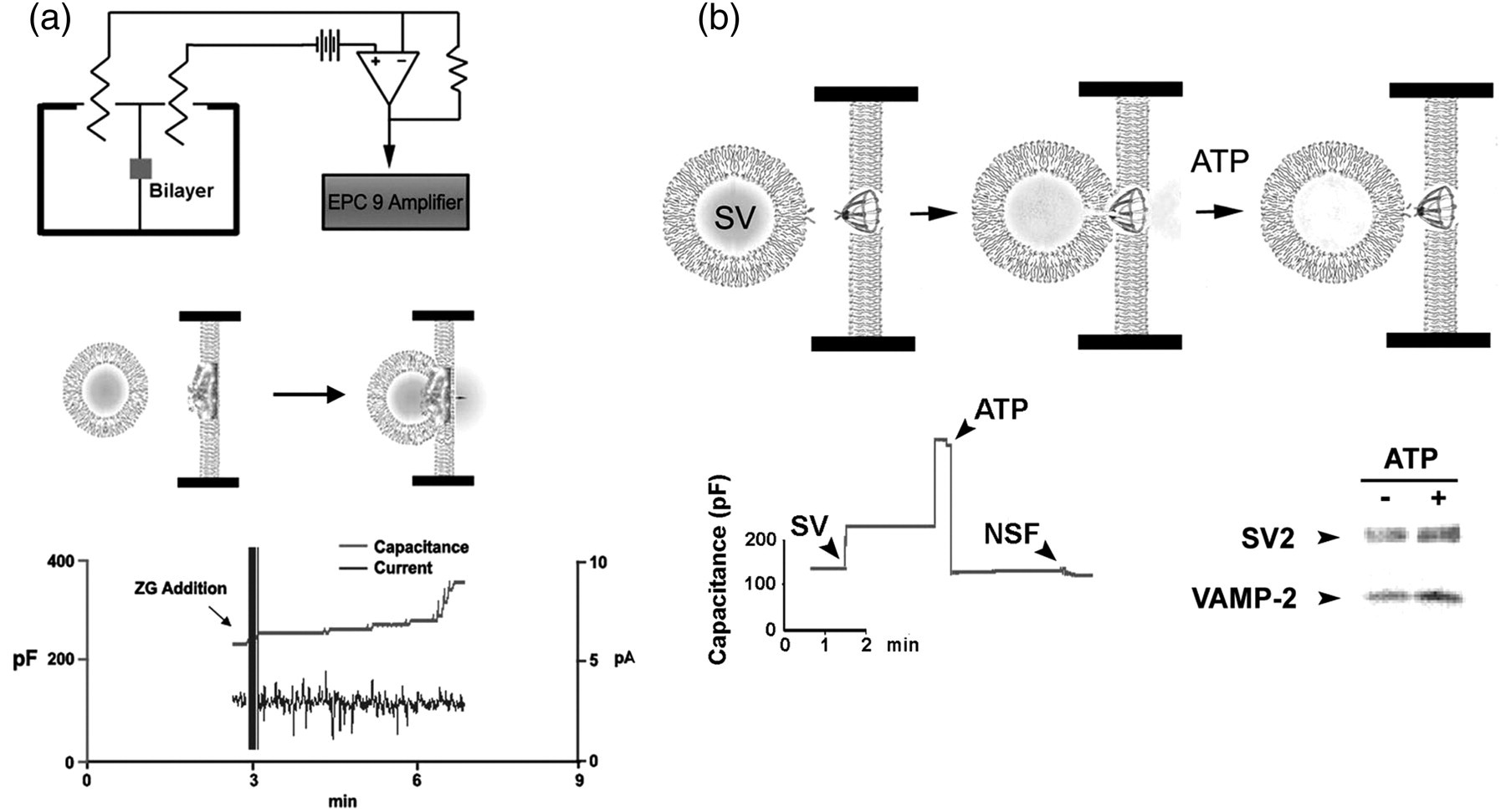
Functionally reconstituted porosome complexes. (a) Functionally reconstituted porosome complex from the rat exocrine pancreas. The top panel is a schematic drawing of the EPC9 (electric computer-controlled patch-clamp amplifier) bilayer set-up for electrophysiological measurements of membrane fusion. Middle panel shows both a schematic drawing of what transpires at the porosome-reconstituted bilayer separating the cis and the trans compartments, and the actual electrophysiological recordings during isolated zymogen granule (ZGs) fusion at the porosome. Addition of isolated ZGs to the cis compartment of the bilayer chamber results in ZG docking and fusion at the reconstituted porosomes, as demonstrated by an increase in capacitance and current activities, and a concomitant time-dependent release of amylase, which is one of the major digestive enzymes present within the ZGs, to the trans compartment of the set-up. The movement of amylase from the cis to the trans side of the chamber is determined using immunoblot analysis of the contents in the cis and the trans compartment, over time.
10
(b) The top left panel shows the schematic representation of a porosome at the presynaptic membrane with a docked synaptic vesicle (SV) at its base.
6
The top right of the figure shows a schematic drawing of an EPC9 electrophysiological bilayers apparatus, to continually monitor changes in the capacitance of the neuronal porosome-reconstituted membrane, when SVs are introduced into the cis compartment of the bilayers chamber.
6
The central panel is a representation of SV docking at the base of a porosome, fusing to release its contents, and disengaging in the presence of ATP. The left bottom panel shows capacitance measurements of porosome-reconstituted bilayers. Exposure of the reconstituted bilayers to SVs results in a dramatic increase in membrane capacitance, which drops to baseline following exposure to 50 μM ATP. Addition of full-length recombinant N-ethylmaleimide-sensitive factor (NSF) has no further effect, suggesting that the porosome preparation has native NSF associated with it. The bottom right panel is immunoblot analysis, essentially confirming what is observed in the reconstituted system, by the fact that exposure of isolated synaptosomal membrane preparations to 50 μM ATP results in the release of SVs from the membrane into the incubation medium, as demonstrated by a gain in SV-specific proteins SV2 and vesicle-associated membrane protein 2 (VAMP-2) in the incubation medium
6
Determination of the composition of porosomes using immunoblot analysis. (a) One-dimensionally resolved proteins immunoisolated from solubilized pancreatic plasma membrane preparations using an SNAP-23 specific antibody. The resolved proteins were silver-stained and transferred to nitrocellulose membranes for immunoblot analysis. Note the identification in the immunoisolates of eight protein bands in a silver-stained sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) 1D-resolved gel (far-left). Immunoblot analysis of the resolved immunoisolated pancreatic porosome proteins demonstrates the presence of actin, vimentin, syntaxin-2, calcium channel, NSF and chloride channels CLC-2 and CLC-3.
10
(b) Neuronal porosome immunoisolated from detergent-solubilized synaptosomal membrane preparation using a SNAP-25 specific antibody.
6
Immunoisolates, when resolved by SDS-PAGE, and the resolved proteins in the gel stained using Sypro dye, reveals 12 specific bands, suggesting the presence of at least 12 proteins in the complex, not including the heavy chain (*) of the SNAP-25 antibody. When the resolved proteins were electrotransferred to nitrocellulose membrane and probed with various antibodies, calcium channel P/Q, actin, Gao, syntaxin-1 (Syn1), synaptotagmin-1 (Syt1), N-ethylmaleimide-sensitive factor (NSF), vimentin and the chloride channel CLC-3 were identified
6

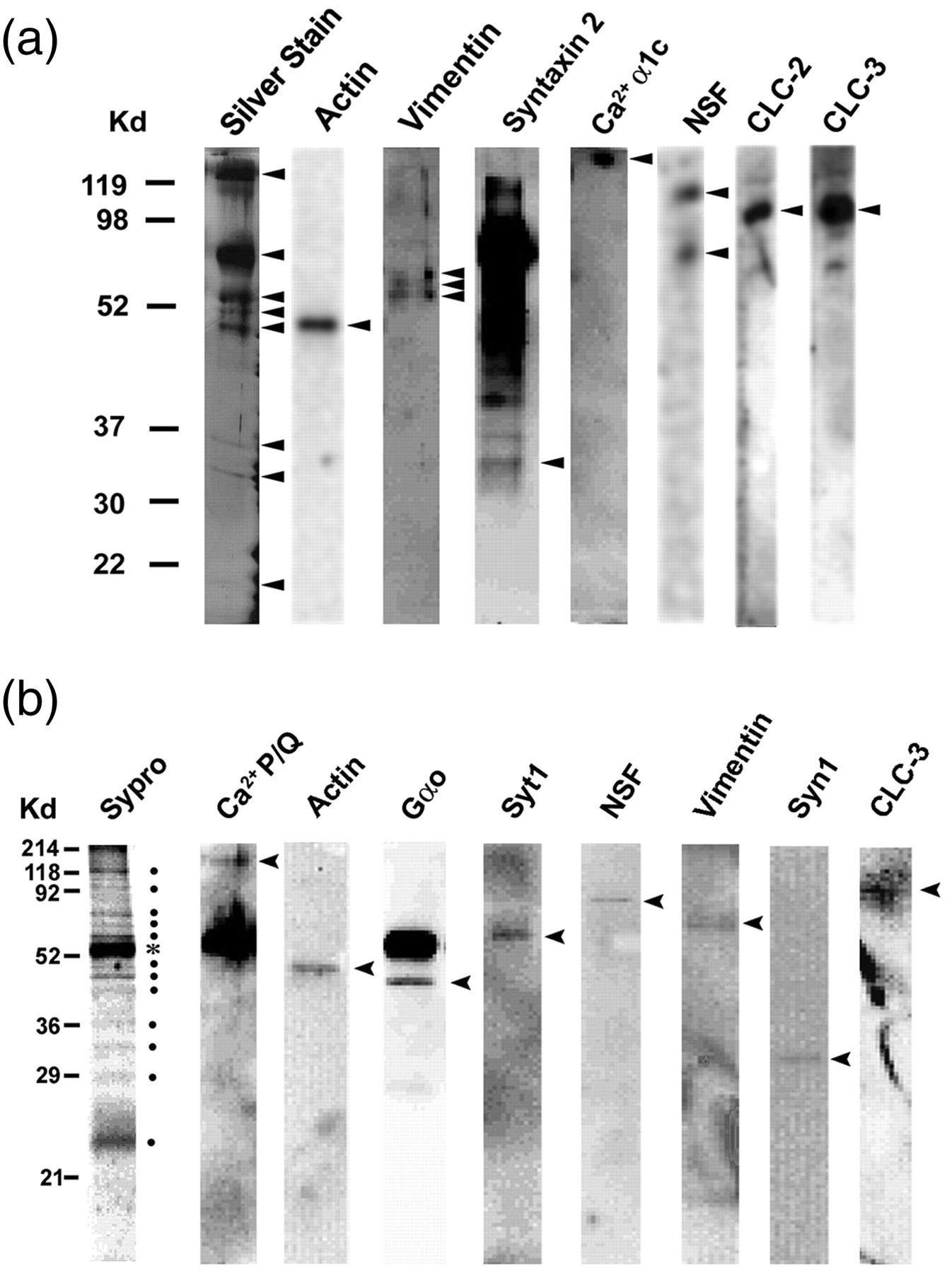
Similarly, porosomes from neurons have also been immunoisolated using SNAP-25 specific antibody using solubilized total rat brains or isolated synaptosomal membrane preparations 6–8 (Figure 5). Neuronal porosomes have also been isolated using gel filtration chromatography. 7 Freshly isolated and solubilized synaptosomal membrane preparations from rat brain have been used in porosome purification using a calibrated G-200 Sephadex gel filtration columns (Pharmacia, Piscataway, NJ, USA). 7 Each fraction is then assayed using immunoblot and mass spectrometry, for the presence of various proteins that may co-elute with t-SNAREs. Besides helping in the determination of porosome composition, these studies have further helped in elucidating the porosome structure, its molecular weight, and its functional reconstitution into artificial lipid membrane. 6–8,10 Results from these studies demonstrate that the neuronal porosome complex measuring 10–15 nm has a molecular mass of >650 kDa. 7 Isolated porosome structure and size have also been determined using AFM, EM and photon correlation spectroscopy. 6–11
Porosome: structure–function
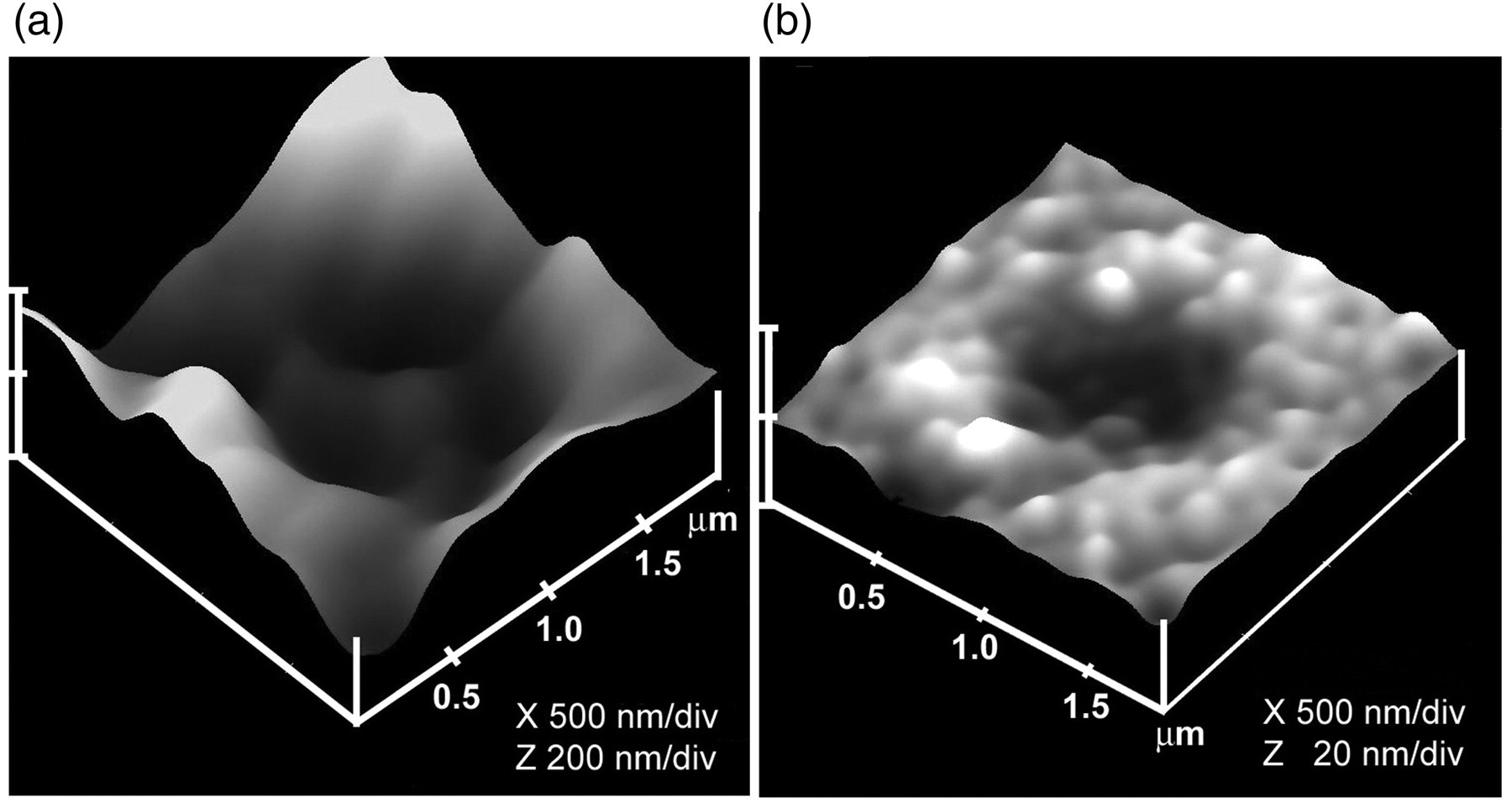
Since porosomes are relatively small structures measuring 100–180 nm in diameter in pancreatic acinar cells or in neuroendocrine cells, and merely 10–15 nm in neurons or astrocytes, it would be extremely rare to be able to take a cross-section through the porosome complex along with any associated secretory vesicles. Nonetheless, porosomes are observed in EM micrographs of isolated cells or tissue preparations (Figures 1 and 2). EM studies confirm porosomes to possess a cup-shaped morphology, with similar dimensions determined using AFM. 2–5 Additionally, EM studies on isolated pancreatic, as well as neuronal porosomes, reveal that the complex possesses both lateral and a number of vertically arranged ridges, possibly protein complexes. A ring-like structure is observed at the base of the porosome (Figure 4), and may represent SNARE proteins arranged in a ring conformation. In the exocrine pancreatic porosome complex, three lateral rings are observed. The two anterior rings may compose cytoskeletal proteins for regulating the opening and closing of the porosome to the outside. 9,10 The composition of the ring at the opening or mouth of the structure is predicted to be actin, since exposure of acinar cells to cytochalasin B, a fungal toxin that inhibits actin polymerization, results in a collapse of the porosome opening and a consequent loss in secretion. 2 Furthermore, immunoanalysis of isolated pancreatic porosomes demonstrate the presence of actin, vimentin and α-fodrin, 9 suggesting the ring to be composed of cytoskeletal proteins, which regulate the opening and closing of the structure to the outside. In contrast, porosomes in neurons and astrocytes possess a central plug with 8–12 vertical spokes lining the porosome cup 6,11 that allow rapid closing and opening of the porosome. The spokes are attached to the central plug at the base of the porosome cup and to a ring at the mouth of the porosome. 6 The central plug in the neuronal porosome complex has been implicated in opening during neurotransmitter release, and in the closing of the complex in its resting state. 23 If the central plug was to function as a stopper, then it would necessitate its vertical movement capability. In agreement, ultrahigh resolution AFM imaging of the presynaptic membrane of isolated synaptosome preparations demonstrate the presence of the neuronal porosome plug at various conformational states, that is, in their open, partially open and close conformation. 23 These results suggest that the central plug, when fully distended outwards, is in the close conformation, when half-way retracted, is in the semi-open state, and when completely retracted into the porosome cup, is in its fully open status, supporting the hypothesis that the central plug operates as an exit door for the neuronal porosome. 23 The presence of such a mechanism in the neuronal porosome complex makes sense, since it would allow the rapid opening and closure of the structure to the outside, during fast neurotransmitter release at the nerve terminal. 23 This is opposed to a slow secretory cell such as the exocrine pancreas or neuroendocrine cell, where no such plug is present at the porosome opening; instead, the opening dilates during secretion, and returns to its resting size following completion of the process. 2,23 Since porosomes are stable structures enabling their isolation, it was hypothesized that if membrane-bound ZGs were to fuse at the base of the porosome, it should be possible to isolate ZG-associated porosomes. To test this hypothesis, ZGs have been isolated from stimulated pancreatic tissue, and the preparation processed for EM. Results from these studies confirm the hypothesis and reveal the structure of porosome-associated vesicles (Figure 1g). The EM micrographs of such porosome-associated vesicles further reveal vertical structures originating from within the porosome complex. The presence of such vertically arranged ridges lining the porosome have also been reported 31 in NG108–15 nerve cells.
Although many advancements have been made in our understanding of the porosome structure–function, the distribution of specific proteins within the porosome complex, remain to be determined. Ideally, atomic coordinates of the complex using either X-ray crystallography or solution nuclear magnetic resonance spectroscopy (NMR) would provide structural details at the atomic level. 8 However, the size and complexity of the membrane-associated porosome, has precluded determination of its atomic structure, which ultimately would provide a molecular understanding of its function. Solution NMR has not been possible primarily due to the large molecular size of the porosome complex, and even the neuronal porosome measuring just 10–15 nm is beyond the operating limits of current NMR. Similarly, X-ray crystallography has not been possible, due in part to the solubility problems of this membrane-associated complex. In recent studies however, these limitations have been partially overcome in furthering our understanding of the fine structure and arrangement of proteins within the native porosome complex in neurons, using high-resolution EM 3D contour mapping 8 (Figure 4). Negatively-staining EM performed using a low electron dose in a Tacnai 20 electron microscope (Philips Electron Optics/FEI, Eindhoven, The Netherlands) operating at 200 kV, demonstrated that proteins at the central plug of the neuronal porosome complex interact with proteins at the periphery of the structure. Similar to what is observed in AFM micrographs, approximately 8–10 interconnected protein densities were observed at the lip of the porosome complex in EM micrographs. The 8–10 interconnected protein densities were also connected to the central plug, via spoke-like elements. Electron density and contour mapping, and the resultant 3D topology profile of the porosome complex, provide further details of the arrangement of proteins in a circular symmetry, and their connection to the central plug via distinct spokes (Figure 4). Furthermore, the center of the porosome complex representing the porosome base where synaptic vesicles dock and fuse, was first demonstrated in 2009 to be composed of three SNARE pairs 32 (Figure 4d); in subsequent studies, 33 in 2010, it was further confirmed that three SNARE pairs are minimally required for fast vesicle fusion and neurotransmitter release in live neuronal cells. Results from electron density, contour mapping and 3D topology studies of isolated porosome complexes, provided for the first time, at the nanometer scale, the arrangement of proteins (Figures 4d and e) within the neuronal porosome complex. 8,32
In recent years, isolated porosomes from both exocrine pancreas, 10 neurons, 6 as well as astrocytes, 11 have been reconstituted in artificial liposomes to determine their reconstituted structure (Figure 4f) and their reconstituted function (Figure 5). Immunoisolated supramolecular porosome complex from the exocrine pancreas has been reconstituted into artificial liposomes, and the liposome-reconstituted complex examined using EM (Figure 4c). EM micrographs reveal a 150–200 nm cup-shaped basket-like structure similar to what is observed in their native states at the cell plasma membrane, and when porosomes are co-isolated with ZG preparations (Figure 1g). To determine if the isolated porosomes are intact and therefore functional, the complexes have been reconstituted into lipid bilayer membrane in an electrophysiological membrane bilayer set-up that is capable of continually monitoring changes in the reconstituted membrane under various experimental conditions (Figure 5). When isolated pancreatic porosomes are reconstituted into a lipid bilayer membrane in such a electrophysiological membrane bilayer set-up, with the lipid membrane separating a cis and a trans compartment, isolated ZGs were found to dock and fuse at the porosome-reconstituted bilayer. ZG fusion at the bilayer is demonstrated by an increase in capacitance and conductance, and a time-dependent expulsion of intravesicular contents from within the ZGs in the cis compartment to the trans compartment of the EPC9 (electric computer-controlled patch-clamp amplifier) electrophysiological bilayer set-up 10 (Figure 5a). It has also been demonstrated that chloride channel activity is associated with the porosome complex, and the chloride channel blocker 4,4'-diisothiocyano-2,2'-stilbenedisulfonic acid inhibits current activity in the porosome-reconstituted bilayer, 10 demonstrating its requirement in the proper functioning of the pancreatic porosome complex.
Similar to the reconstitution of isolated porosomes from the exocrine pancreas in artificial lipid membrane, 10 neuronal porosomes have been both structurally and functionally reconstituted. 6 To test whether complete and functional neuronal porosomes are immunoisolated, the immunoisolated preparations have been reconstituted to lipid membrane prepared using brain dioleoylphosphatidylcholine (DOPC) and dioleoylphosphatidylserine (DOPS) in a ratio of 7:3, respectively. 6 These studies demonstrate the presence of porosomes in patches similar to what is observed at the presynaptic membrane in intact synaptosomes. Furthermore, imaging of the reconstituted immunoisolate at ultrahigh resolutions using AFM, demonstrates the presence of 10–15 nm porosomes having a 2 nm in diameter central plug as observed in their native state 6 (Figure 4f). These studies confirm the complete isolation and structural reconstitution of the neuronal porosome complexes into artificial lipid bilayers. 6 To assess the functionality of the reconstituted neuronal porosome preparations, electrophysiological fusion assays using isolated synaptic vesicle preparations and membrane-reconstituted porosomes have been used, 6 and membrane capacitance and conductance is continually monitored (Figure 5b). Following reconstitution of the bilayer membrane with neuronal porosomes, isolated synaptic vesicles are added to the cis compartment of the bilayer chamber in the EPC9 electrophysiological set-up. Results from such experiments demonstrate that a large number of synaptic vesicles fuse at the porosome-reconstituted bilayer, reflected by the stepwise increase in membrane capacitance 6 (Figure 5b). The addition of 50 μmol/L ATP to the cis compartment of the bilayer chamber results in t-/v-SNARE disassembly and the release of docked vesicles, followed by the return of the bilayers membrane capacitance to resting levels, demonstrating transient fusion of synaptic vesicles at the bilayer-reconstituted porosome complex. 6 To further assess if such a mechanism of the release of docked synaptic vesicles following ATP exposure occurs in synaptosomes, synaptosomal membrane preparations were exposed to 50 μmol/L ATP, and the supernatant fraction was assessed for synaptic vesicle association by monitoring levels of the synaptic vesicle proteins SV2 and VAMP-2. Results from the study demonstrate that both SV2 and VAMP-2 proteins are enriched in supernatant fractions following exposure of isolated synaptosomal membrane to ATP, demonstrating release of synaptic vesicles. Thus, AFM, electrophysiological measurements and immunoanalysis, confirm the dissociation of porosome-docked synaptic vesicles in isolated synaptosomal membrane preparations following ATP exposure. 6 As previously suggested, 2 such a mechanism may allow for multiple rounds of transient docking–fusion and release events that synaptic vesicles may undergo during neurotransmitter release, without compromising vesicle identity and integrity. 13–15 Isolated inside-out synaptosomal membrane preparation, when placed on mica and imaged by AFM in near physiological buffer, demonstrates the presence of 40–50 nm synaptic vesicles docked to neuronal porosomes at the presynaptic membrane. Similar to the ZGs, exposure of synaptic vesicles to GTP, resulted in an increase in synaptic vesicle swelling, and further exposure to Ca2+ resulted in the transient fusion of synaptic vesicles at the presynaptic membrane, expulsion of intravesicular contents and the consequent decrease in synaptic vesicle size. Therefore, synaptic vesicles do not completely merge at the presynaptic membrane during neurotransmitter release. 1
Porosome composition: immunoblot and mass spectrometry
Studies demonstrate the porosome to include among other proteins, SNAP, syntaxin, the cytoskeletal proteins actin, α-fodrin and vimentin, calcium channels β3 and α1c, together with the SNARE regulatory protein NSF 9,10 (Figure 6). Chloride ion channels CLC-2 and CLC-3 have also been identified as part of the porosome complex, and as previously stated, are critical to porosome function. Isoforms of the various proteins identified within the porosome complex have been also been demonstrated using two-dimensional benzyldimethyl-n-hexadecylammonium chloride (2D-BAC) gel electrophoresis. As an example, three isoforms each of the calcium ion channel and vimentin are found in porosomes. Similarly, using yeast two-hybrid analysis 12 and immunoisolation, studies have confirmed the presence and direct interaction between some of these proteins with t-SNAREs within the porosome complex. For example, direct interaction between SNAP-23 and L-type calcium channel has been reported in the pancreas. 12 To further determine the composition of the neuronal porosome complex and associated proteins, the complex has been both immunoisolated from solubilized rat brain synaptosome preparations using SNAP-25 specific antibody and using gel filtration chromatography, followed by matrix-assisted laser desorption ionization-time of flight mass spectrometry. Results from these studies demonstrate the association of additional proteins previously undetected using immunoblot analysis of the neuronal porosome complex. Currently, more than 30 proteins have been identified to compose the neuronal porosome (unpublished observation). In addition to these proteins, studies report that the neuronal porosome assembly requires membrane cholesterol. 7 Results from recent studies 7 demonstrate a significant inhibition in the interaction between porosome-associated t-SNAREs and calcium channels following depletion of membrane cholesterol. Since calcium is critical to SNARE-induced membrane fusion, the loss of interaction between SNAP-25, syntaxin-1, and calcium channels at the neuronal porosome complex, would seriously compromise or even abrogate neurotransmitter release at the nerve terminal.
In summary, these studies demonstrate porosomes to be permanent, supramolecular, lipoprotein structures at the cell plasma membrane, where membrane-bound secretory vesicles transiently dock and fuse to release intravesicular contents to the outside (Figure 7). Porosomes may therefore be considered the universal secretory portals in cells.
34–49
In recent years, there has been a flurry of research in the field, and a number of studies from several laboratories investigating the porosome in different cell types, both in their native and disease states, such as porosomes in neurons, in sensory hair cells and in RBL-2H3 and BMMC cells, have been reported.
16–22
A detailed study of porosomes in the exocrine pancreas
50
and the neuronal porosome proteome
51
has recently been elucidated. The discovery of the porosome, its structure, function, composition and its functional reconstitution into artificial lipid membrane, has resulted in a paradigm shift in our understanding of the secretory process in cells. The AFM has enabled the determination of live cellular structure–function at sub-nanometer to angstrom resolution, in realtime, contributing to the birth of the new field in Cell Biology, NanoCellBiology. Future directions will involve an understanding of the protein distribution and their arrangement at the atomic level within the porosome complex. This objective is currently being pursued using electron crystallography.
Schematic diagram depicting the transient docking and fusion of secretory vesicles at the porosome base in the cell plasma membrane, increase in turgor pressure of vesicles via the entry of water through water channels called aquaporins (AQP), expulsion of vesicular contents, and the dissociation of vesicles from the cell plasma membrane.
51
NSF, N-ethylmaleimide-sensitive factor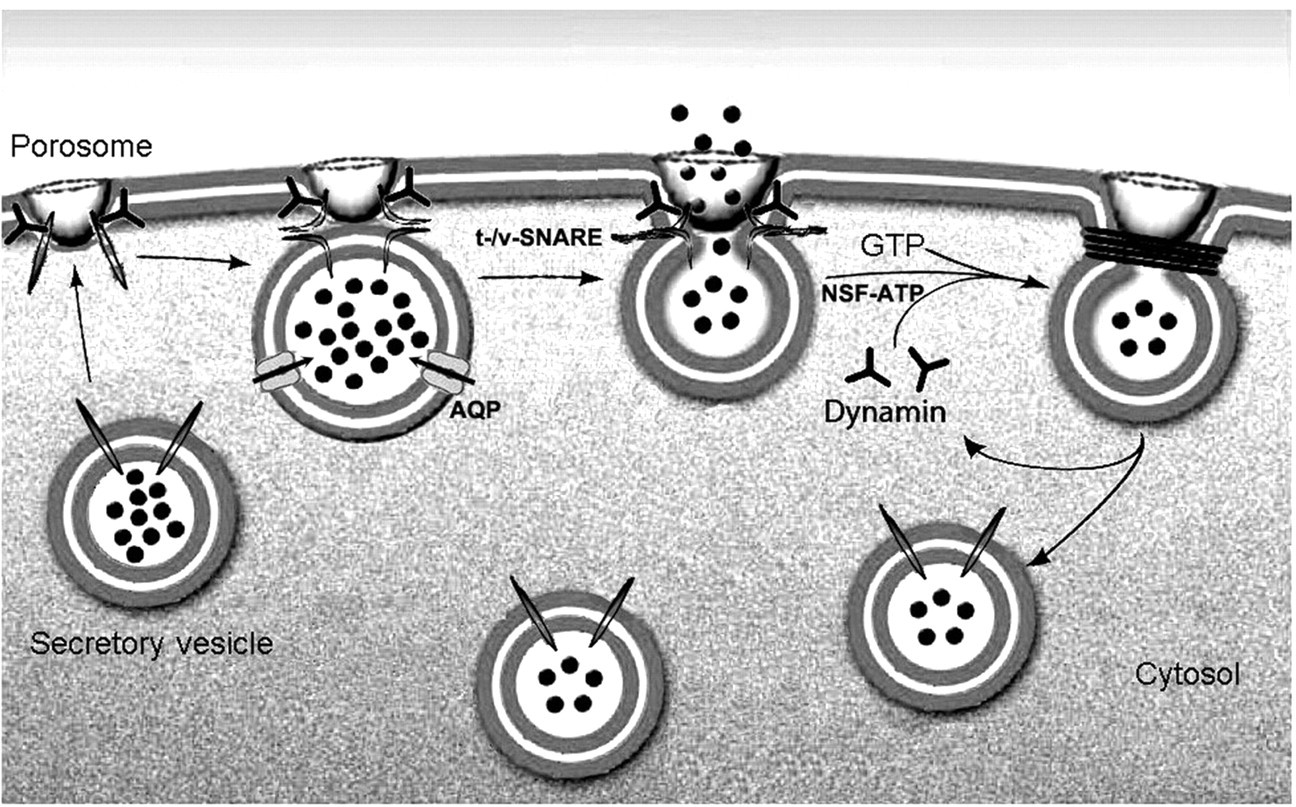
Footnotes
Acknowledgements
The author would like to thank Jin-Sook Lee and Won Jin Cho for their help in preparation of the figures. This work was supported by a grant from the National Science Foundation.