
Abstract
To evaluate the influence of low temperatures on the proliferation of neural stem cells (NSCs) and the regulation of their signaling pathways after brain trauma, we examined changes in the expression levels of specific miRNAs and their target genes. We also evaluated NSC proliferation in the hippocampus after brain trauma under low-temperature conditions. We found that the expression profile of miRNAs in the hippocampus after trauma changed at both normal and low temperatures, and the expression of miR-34a decreased significantly lower in rats exposed to low temperatures. There was significant proliferation of endogenous NSCs in the hippocampus after brain trauma at both temperatures, but NSC proliferation was slower at low temperatures. In addition, the expression of Notch1 significantly increased in the hippocampus after brain trauma at both temperatures. However, at low temperatures, the degree of up-regulation of Notch signaling molecules was significantly lower. We conclude that low-temperature environments can inhibit the proliferation of endogenous NSCs in the hippocampus, possibly by alleviating the effects of miR-34a down-regulation and Notch signaling up-regulation induced by traumatic brain injury.
Introduction
Traumatic brain injury (TBI) is a condition that severely threatens human health, causing neuronal degeneration, necrosis and neurological dysfunction. The discovery of neural stem cells (NSCs) in 1992 brought new light to the treatment of craniocerebral injuries. 1 It has been found that TBI can activate the proliferation and differentiation of dormant NSCs, which then migrate to specific areas and integrate with existing neural circuits, causing limited improvement to damaged neurological functions.2,3 Further investigation into the effects of trauma on NSC proliferation and differentiation, the mechanisms by which they are regulated after trauma, and the ways to improve the ability of NSCs to proliferate and differentiate after craniocerebral injury, is key to developing better therapies for brain trauma patients.
Studies have indicated that microRNAs (miRNAs) have an important regulatory role in the proliferation and differentiation of NSCs during the embryonic stage and in adults. In particular, miR-34a inhibits the expression of Notch1 signaling molecule, which is closely involved in NSC proliferation and differentiation.4–8 In addition, it has recently been shown that the expression levels of nearly 100 miRNAs in the hippocampus change after brain trauma. 9 This suggests that it is likely that miRNAs are involved in the regulation of NSC proliferation and differentiation after brain trauma.
During the past few years, extreme climatic events have occurred more frequently as a consequence of global warming. Under freezing conditions, the incidence of traumatic injuries increases drastically. Research has also shown that prolonged exposure to low-temperature environments may damage multiple body systems including the nervous, circulatory, digestive and respiratory systems. 10 However, it remains unclear as to how low-temperature environments influence the proliferation and differentiation of NSCs in the hippocampus after brain trauma. In this study, we developed a mouse model of closed head injury (CHI). We then used miRNA chip technology and real-time polymerase chain reaction (RT-PCR) to examine changes in the expression levels of specific miRNAs and their target genes, and to assess NSC proliferation in the hippocampus, at different time points after brain trauma under low-temperature conditions. This allowed us to evaluate the influence of low temperatures on NSC proliferation and the regulation of its signaling pathway after brain trauma.
Materials and methods
Trauma model and experimental groups
The study was conducted according to the guidelines for the humane and ethical treatment of research animals of the Institutional Animal Care and Use Committee of Nanchang University. Male BALB/c mice weighing 20–25 g, aged eight weeks, were obtained from the Experimental Animal Department of Nanchang University. Experimental CHI was performed, as previously described. 11 Briefly, the animals were anesthetized with iso-flurane; their heads were fixed on the stereotaxic frame; and their skulls were exposed by longitudinal incision of the skin. Focal trauma was induced to the left hemisphere 1–2 mm lateral to the midline in the mid-coronal plane using a weight drop device with a 40-g metal rod falling freely from a height of 25 cm. An impactor tip was placed at the left lateral to the midline in the mid-coronal plane. After trauma, the mice received supporting oxygenation with 100% O2 until fully awake. They were then returned to their cages and given food and water ad libitum. Sham-operated animals underwent anesthesia and scalp incision but were not subjected to experimental head trauma.
Neurobehavioral evaluation
The neurological impairment of mice was assessed according to a modified neurological severity score (NSS) one hour post-CHI, referred to as NSS1. Assessors were not informed what treatment had been given to the mice. The NSS1 reflects the initial severity of injury and is inversely correlated with clinical outcome.12,13 Only mice with an NSS of 7–8, representing severe injury, were chosen for experiments. The modified NSS system consists of 10 individual clinical tasks involving motor function, alertness and physiological behavior (Table 1). One point is awarded for failing to perform a particular task.
NSS for head-injured mice

For each failed task, the mouse receives 1 point (maximum = 10 [failure in all tasks], minimum = 0 [success in all tasks]). Some of the animals were killed four hours after skin incision, and their brains were removed for immunohistochemical analysis. The remaining animals were divided randomly into a normal-temperature TBI group (NTBI group) and a low-temperature TBI group (LTBI group). Each group was divided into four subgroups: sham, 4-hour, 1-day and 3-day. The NTBI group was kept at ≍25°C and the LTBI group was kept in climate boxes at 4°C for four hours, then at ≍25°C . To determine the microRNA expression profile in the hippocampus after TBI, both sham animals and injured animals that had experienced normal-and low-temperature exposure were killed four hours post-injury and subjected to miRNA microarray analysis. To determine the expression levels of miR-34a and Notch1 mRNA in the hippocampus, animals from both the normal- and low-temperature groups were killed four hours, one day and three days after injury (n = 6 for each time point). They were subjected to RT-PCR analysis. For analysis of Notch1 and nestin protein expression levels and of the proliferation response of NSCs in the hippocampus after TBI, injured animals from normal- and low-temperature groups were killed after three days (n = 6 for each group).
Tissue preparation
Animals were killed with an overdose of chloral hydrate. Animals were perfused through the heart with 50 mL normal saline followed by 50 mL phosphate-buffered saline (PBS) containing 4% paraformaldehyde. The brains were removed and fixed in 4% paraformaldehyde at 4°C for 24 h and then cryoprotected in 20% and 30% sucrose solution. Coronal sections of the brain (1-4 mm behind the bregma) were obtained using freezing microtome. Each slice was 8 /μm thick. Every 10th slice was processed for hematoxylin and eosin (H&E) staining or immunofluorescence. Figure 1 shows H&E stains of the cerebral cortex and hippocampus after TBI. For RNA extraction, animals were perfused through the heart with normal saline, and then the ipsilateral hippocampus was dissected.

Hematoxylin and eosin staining of cerebral cortex and hippocampus after closed head injury, confirming that serious injury had been delivered to the mice used in this study. (a) sham group; (b) CHI group. Bar = 20 μm. (A color version of this figure is available in the online journal)
RNA extraction
Total RNA, including miRNA, was extracted from the ipsilateral hippocampus (approximately 30 mg) with the mirVana total RNA extraction kit according to the instructions from the manufacturer (Applied Biosystems/ Ambion). The quality and quantity of the RNA was evaluated by 260/280 ratio and Agilent 2100 Bioanalyzer (Agilent Technologies, Clara, CA, USA).
Array hybridization and data analyses
Microarrays from Agilent Technologies were used to assess the level and composition of miRNA. In brief, ≍100ng of total RNA was dephosphorylated and ligated with pCp-Cy3. Labeled RNA was purified and hybridized to Agilent miRNA arrays (v12.0). Images were scanned with the Agilent microarray scanner (Agilent), gridded and analyzed using Agilent feature extraction software version 9.5.3. Microarray analysis involved multiple sample analyses, including normalization, data adjustment, t-test/analysis of variance (ANOVA), and clustering. The t-test was performed between the control and test-sample groups. T values were calculated for each miRNA, and probability values were computed from the theoretical t distribution. ANOVA was performed across the three groups. P values were calculated for each miRNA. Raw data were analyzed with GeneSpring GX software (Agilent Technologies, Inc., Clara, CA, USA).
RT-PCR analysis of mRNA and miRNA
According to the method described by Varkonyi, 13 RT-PCR analysis of mature microRNA expression was performed as follows: cDNA was synthesized with miRNA stem-loop primer and 100 ng of total RNA. TransStart Green qPCR SuperMix (Beijing TransGen Biotech Co., Ltd, Beijing, China) was used to conduct RT-PCR using a 7500 real-time PCR System (Applied Biosystems) according to the following protocol: 95°C for 10 min; 40 cycles of 95°C for 15 s and 60°C for 30 s, and then a melt curve analysis. The relative expression of miRNA and mRNA expression data were normalized to U6 RNA and glyceraldehydes-3-phosphate dehydrogenase by 2 2 ΔΔCT, respectively. Primer sequences are shown inTable 2.
Primer sequences of miRNAs used for realtime polymerase chain reaction

Immunofluorescence staining
The frozen sections were rehydrated with PBS and then incubated in the blocking solution (2% goat serum, 0.35% Triton X-100 and 1% bovine serum albumin in PBS) for one hour at room temperature. Primary antibodies diluted in blocking buffer (mouse monoclonal anti-Notch1, Chemicon, Billerica, MA, USA; 1:100 and rabbit polyclonal anti-nestin, Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA; 1:100) were added to individual sections and incubated overnight at 4°C. The sections were then washed with PBS and incubated for one hour at room temperature with corresponding secondary antibody (anti-mouse or anti-rabbit antibody, Santa Cruz, both 1:200). The stained sections were dehydrated, sealed with cover slips, examined and photographed under a laser scanning confocal microscope (Olympus, Tokyo, Japan). The number of positive cells in the ipsilateral hippocampal dentate gyrus (DG) were counted blindly in five coronal sections per animal. For each section, 10 randomly selected non-overlapping fields of vision were evaluated with a ×40 objective using a microscopy, and the average value was taken.
Western blot analysis
The ipsilateral hippocampus was homogenized in RIPA protein lysis buffer (Cell Signaling, Danvers, MA, USA) and 50 μg of protein was analyzed by immunoblotting. The protein was separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes using a trans-blot apparatus (TRANS-BLOTRSD, Bio-Rad, Philadelphia, PA, USA). After blocking in Tween 20/Tris-buffered saline (TTBS) containing 5% (w/v) non-fat milk at room temperature for one hour, the membranes were incubated with a 1:1000 dilution of primary antibody (mouse monoclonal anti-Notch1 or rabbit polyclonal anti-β actin) overnight at 4°C. Blots were washed three times with TTBS and incubated with a 1:2000 dilution of peroxidase-conjugated rabbit anti-mouse IgG secondary antibody or horseradish peroxidase-conjugated goat anti-rabbit IgG secondary antibody (Santa Cruz Biotechnology Inc), respectively, for one hour at room temperature. Blots were again washed three times with TTBS and then visualized by enhanced chemilumines-cence (Pierce, USA), followed by autoradiography at several exposures to ensure appropriate grayscale density.
Statistical analysis
Data are shown as mean ± SEM and analyzed by SPSS17.0 (IBM Corporation, Somers, NY, USA). Statistical comparisons of data were performed by one way ANOVA or two-sample t-test. P < 0.05 was regarded as statistically significant.
Results
Experimental CHI model mice with NSS scores of 7–8 were chosen for experimentation
Histopathological analysis revealed significant cerebral contusions, cellular edema, neuronal cell death, after TBI, showing that the CHI mouse model had been established successfully (Figure 1).
Body temperature
Rectal temperature was measured with a digital thermometer and taken as the body temperature. The rectal temperature of each mouse was measured with a digital thermometer (TD-300, Shibaura Electric Co Ltd, Tokyo, Japan). The ambient temperature in the animal quarters was 25 or 4°C, as needed, and the body temperature of each animal in the LTBI group was found to change with the ambient temperature (Table 3).
The body temperature of the animal in each group (°C, χ- ± s)
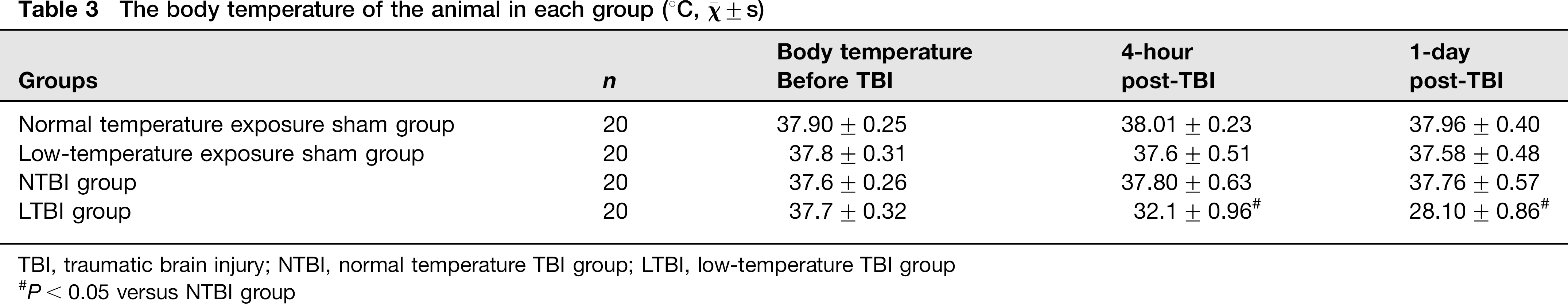
TBI, traumatic brain injury; NTBI, normal temperature TBI group; LTBI, low-temperature TBI group
P < 0.05 versus NTBI group
NSC proliferation in the hippocampus after TBI
Nestin is a specific marker for NSCs. In the present study, immuno-fluorescence staining of nestin was employed to determine the level of NSC proliferation in the ipsilateral hippocampal DG after TBI. The results showed that clustered nestin-positive cells were predominately located in DG. Three days after the injury, both the NTBI and LTBI groups showed more positive cells than the sham group. There were fewer nestin-positive cells in the LTBI 3-day group than in NTBI 3-day group (Figures 2 and 5a). These data indicate that TBI can induce NSC proliferation in the hippocampus, and low-temperature exposure revealed a decrease in trauma-induced proliferation.

Immunofluorescence staining of nestin in the ipsilateral DG at three-day postinjury. (a) Sham group with normal temperature exposure; (b) 3-day postinjury group with normal-temperature exposure; (c) sham group with low-temperature exposure; and (d) 3-day postinjury group with low-temperature exposure. Higher numbers of nestin-positive (red) cells were observed in the ipsilateral DG in three-day mice that had been exposed to either normal or low temperatures than in the sham group. Six animals per group. Bar=20 μm. DG, dentate gyrus. (A color version of this figure is available in the online journal)
MiRNA expression profile
Of the 627 miRNAs spotted on the microarrays (Sanger miRBase database Release version12.0, Sanger Institute, Cambridgeshire, UK), 161 miRNAs (26%) were detected in the hippocampus of the sham group. At four hours post-TBI, the number of miRNAs detected increased to 252 (40%) in the NTBI group and 209 (33%) in the LTBI group. Thirty-eight miRNAs showed more than two-fold altered expression in the NTBI and LTBI groups relative to the sham group. Of these miRNAs, 31 showed more than two-fold altered expression (24 up-regulated and 7 down-regulated) in the NTBI group and 23 miRNAs showed more than two-fold altered expression (20 up-regulated and 3 down-regulated) in the LTBI group (Table 4). Among the altered miRNAs, most of them (30 of 38) showed less expression in the LTBI group than in the NTBI group. miR-200b was up-regulated more than two-fold and miR-34a was down-regulated more than two-fold in the NTBI group. These results were further validated by RT–PCR (Figure 3b). After ANOVA, miRNAs with probability values below a critical value (typically 0.05) in all three groups were selected for hierarchical cluster analysis (Figure 3a). In hierarchical clustering, the samples showed two distinct clusters: (a) a cluster containing the sham and LTBI groups and (b) another containing the NTBI group. This revealed that the miRNA expression profile in the LTBI group was more similar to that of the sham group, suggesting that low-temperature exposure affects miRNA expression in the hippocampus after TBI. All these results indicate that TBI can induce changes in miRNA levels in the hippocampus and that low-temperature environments can affect miRNA expression after brain injury.
The expression levels of certain miRNAs were altered more than two-fold after TBI with exposure to normal or low temperatures
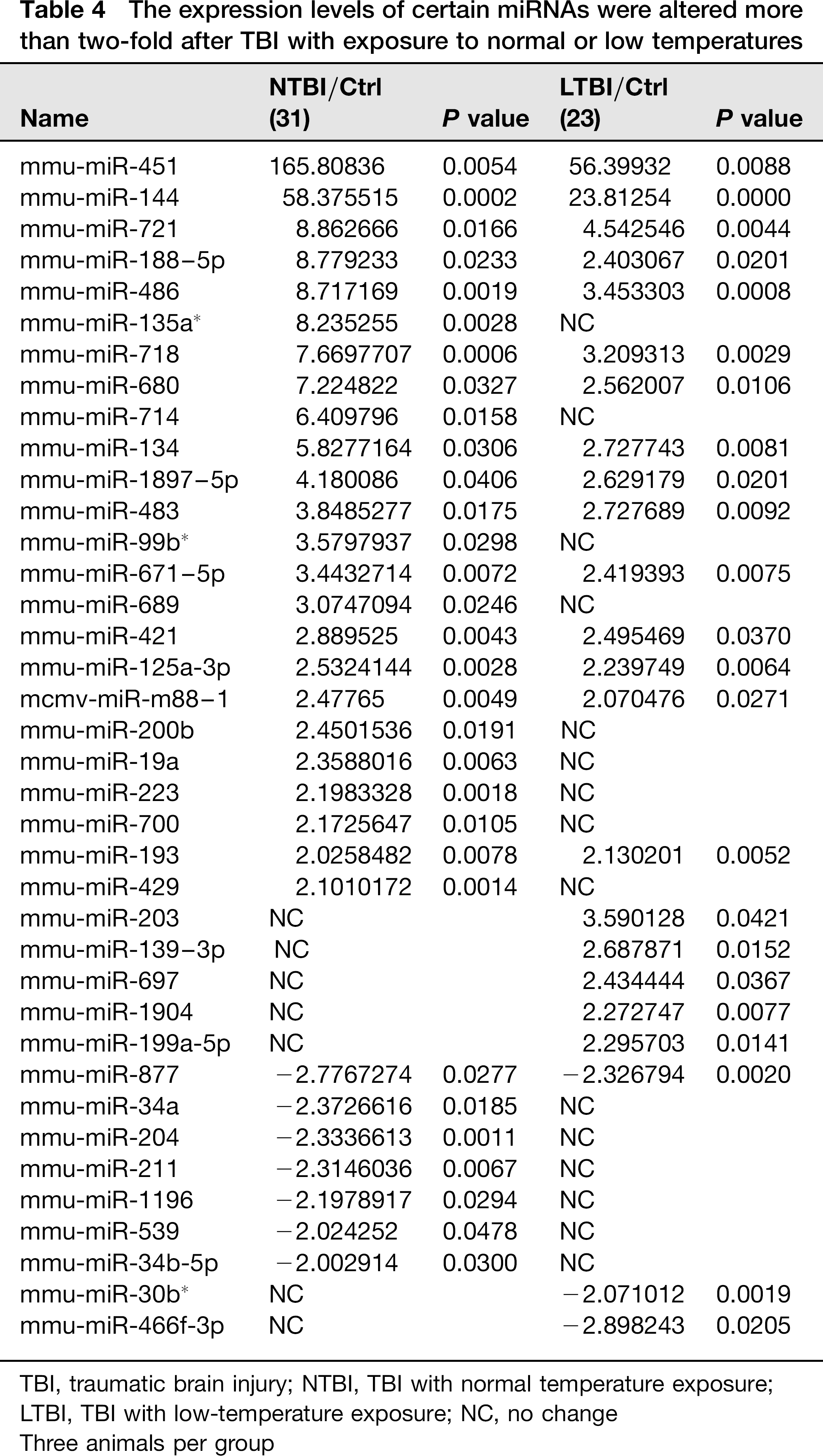
TBI, traumatic brain injury; NTBI, TBI with normal temperature exposure; LTBI, TBI with low-temperature exposure; NC, no change
Three animals per group
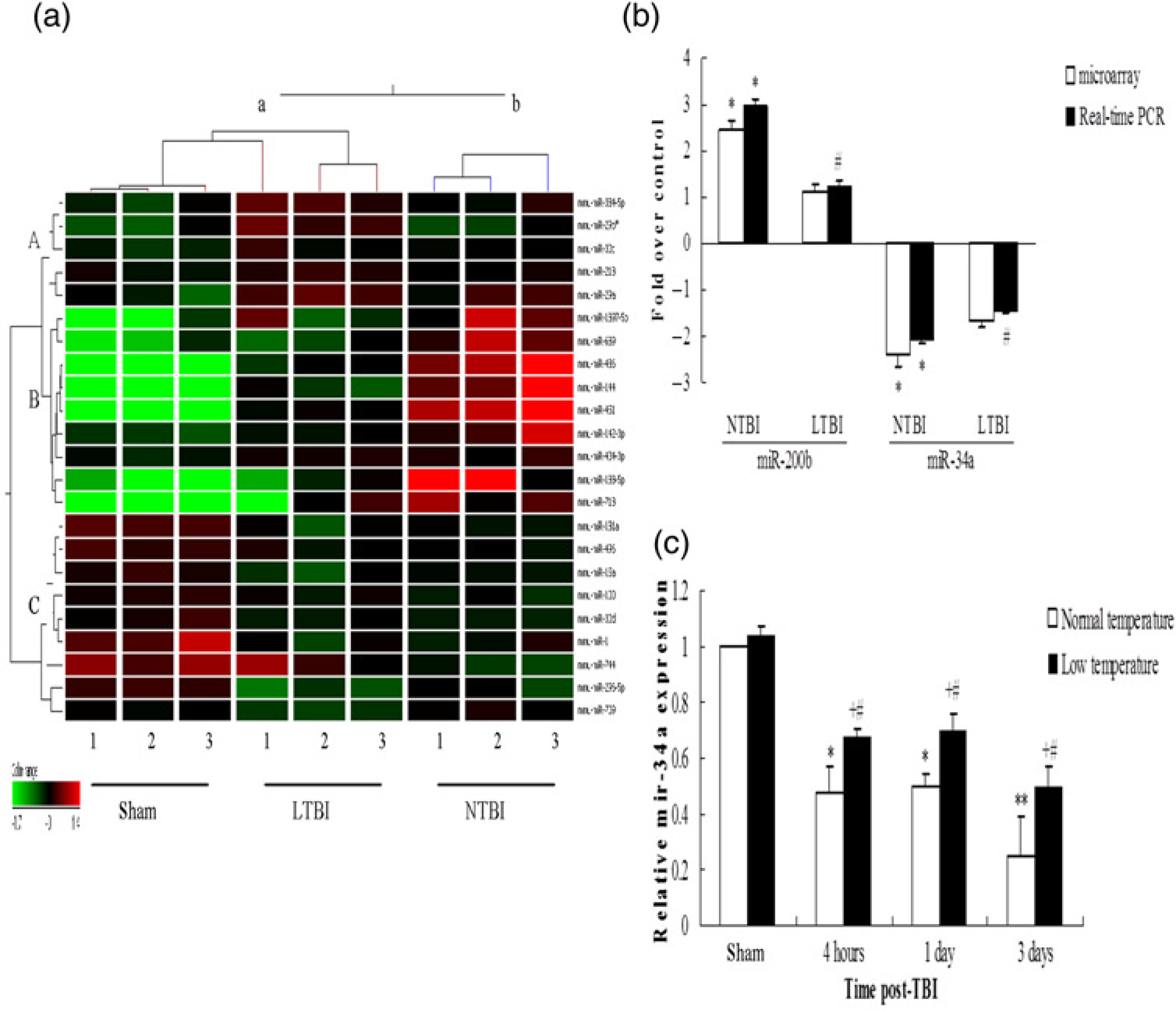
miRNAs expression analysis. (a) Hierarchical clustering analysis of miRNAs expressed differently in the sham group, NTBI and LTBI groups (n = 3/ group). miRNAs whose levels were altered significantly (P
MiR34a/Notch1 signaling
We next determined whether miR-34a had a pivotal role in the regulation of NSC proliferation in the hippocampus after TBI by targeting the Notch1 pathway in vivo. The miR-34a level was determined in the hippocampus four hours, one day and three days after TBI. Our results revealed that (1) miR-34a levels in the NTBI group were down-regulated significantly four hours, one day and three days postinjury relative to those of the sham group (P < 0.05 or P < 0.01); (2) miR-34a levels in LTBI group were also down-regulated significantly four hours, one day and three days postinjury relative to those of the sham group (P < 0.05); and (3) miR-34a levels four hours, one day and three days after injury were lower in the NTBI group than in the LTBI group (P< 0.05), suggesting that TBI can induce down-regulation of miR-34a and that low-temperature environments can alleviate the effects (Figure 3c). We then investigated the expression pattern of its target, Notch1, in the hippocampus after TBI. Immunofluorescence staining of Notch1 displayed clustered Notch1-positive cells in the ipsilateral hippocampal DG (Figure 4b). Notch1-positive cell quantification analysis showed increased numbers of Notch1-positive cells in the ipsilateral hippocampal DG three days postinjury in both the NTBI and LTBI groups relative to the sham group (P < 0.01, Figure 5b). Three days after injury, there were fewer Notch1-positive cells in the ipsilateral hippocampal DGs of the LTBI group than in those of the NTBI group (P < 0.01, Figures 4d and 5b). Western blot analysis confirmed these results (Figures 5c and d). This suggested that TBI can induce up-regulation of Notch1 protein and that low temperatures can weaken the induction. These data collectively indicate that TBI can decrease miR-34a expression significantly and increase the expression of its target, Notch1. In this way, it can induce NSC proliferation in the hippocampus. This suggests that the miR-34a/Notch1 pathway might have a role in trauma-induced NSC proliferation in vivo. In addition, exposure to low temperatures might affect NSC proliferation by weakening activation of the miR-34a/Notch1 pathway in the hippocampus after injury.

Immunofluorescence staining of Notch1 at three days post-TBI. (a) Sham group exposed to normal temperatures; (b) 3-days postinjury group exposed to normal temperatures; (c) sham group exposed to low temperatures; and (d) 3-days post-injury group exposed to low temperatures. Increased notch1-labeling (green) was observed in both 3-days post-injury groups relative to their respective sham groups. Six animals per group. Bar = 20 μm. TBI, traumatic brain injury. (A color version of this figure is available in the online journal)

Quantification analysis of the number of nestin or Notch1-positive cells in the ipsilateral DG and immunobloting of Notch1 at three days post-TBI. (a) Quantification analysis of the number of nestin-positive cells in the ipsilateral DG. #P
Discussion
Studies have shown that TBI can stimulate endogenous NSCs to proliferate and differentiate into mature neurons that integrate into existing neural networks, partially restoring lost neurological function.14,15 In our experiments, we observed that, at normal temperatures, the number of nestin-positive cells in the hippocampus DG area on the lesioned side in the brain trauma group was significantly higher than in the sham-operated group. This suggests that brain trauma can indeed induce proliferation of NSCs in the hippocampus DG area. Promoting the recovery of neurological function by maximizing the proliferation and differentiation of endogenous NSCs may become a new therapeutic strategy for the treatment of lesions of the central nervous system, such as those caused by brain trauma.
Among the many signaling pathways involved in regulating the proliferation and differentiation of NSCs, the Notch signaling pathway is the most important. It is closely related to NSC proliferation and differentiation processes: activation of Notch signaling can help maintain the undifferentiated state of NSCs and promote their proliferation, whereas inhibition of Notch signaling can promote the differentiation of NSCs into neurons. 6 It has been shown that components of Notch signaling are also expressed in the hippocampus and SVZs of adult mammalian brains, suggesting that Notch signaling is involved in the regulation of NSC proliferation and differentiation after trauma.7,8 Here, we observed that, at normal temperatures, the expression level of Notch1 mRNA and the number of Notch1-positive cells in the hippocampus DG on the lesioned side of the brain were both significantly higher in the brain trauma group than those in the sham-operated group. This, as well as the changes in NSC proliferation in different experimental groups, has led us to conclude that brain trauma can induce activation of the Notch signaling pathway in the hippocampus, promoting NSC proliferation and differentiation.
In recent years, a growing number of articles have been published describing a link between the expression of microRNAs and NSC proliferation and differentiation after brain trauma. Studies have shown that, after brain trauma, the expression levels of many specific miRNAs involved in the proliferation and differentiation of NSCs change significantly.9,16,17 We found here that, at normal temperatures, the expression levels of 24 miRNAs were up-regulated to at least twice those of the control. The expression levels of 7 miRNAs were down-regulated to less than half that of the control. The expression levels of miR-34a decreased significantly at normal temperatures, suggesting that brain trauma can induce abnormal expression of miR-34a.
In studies related to the proliferation and differentiation of tumor stem cells, it has been found that miR-34a can negatively regulate Notch1 signaling and inhibit the proliferation and differentiation of tumor stem cells.2,3 Recently, Fineberg et al. 18 reported that miR-34a represses Numbl in murine neural progenitor cells and antagonizes neuronal differentiation. Our results also showed that brain trauma can lead to a decrease in miR-34a expression and an increase in the expression levels of Notch signaling molecules in the hippocampus. These results have led us to conclude that, in the proliferation of NSCs that occurs in the hippocampus after brain trauma, miR-34a has a regulatory role. This role involves influencing the expression of the components of the Notch signaling pathway.
Low temperatures can exert many adverse effects on the body and cause impairment of many systems including the central nervous system. 10 Kanagawa et al. 19 found the severe hypothermic environment induced a decrease of neurogenesis in the neonatal rat. Clinical observation has shown that exposure to low temperatures can lead to further damage in trauma patients and increase the mortality rate.20,21 However, the impact of low-temperature environments on TBI patients requires further in-depth investigation.
The temperature and time settings for low-temperature exposure have varied across previous experiments. Here, we chose a four-hour exposure to 4°C after trauma. We found that the body temperatures of normal mice showed no significant change after four hours of exposure to 4°C. However, TBI mice showed a progressive decrease in body temperature as exposure continued. The decrease in body temperature was accompanied by changes in the behavior of their NSCs. This suggested that a four-hour exposure to 4oC after injury may be sufficient to cause pathological and physiological changes in TBI mice.
In this study, the expression of miR-34a decreased significantly at both normal and low temperatures. However, the degree of down-regulation of miR-34a at low temperatures was significantly lower than at normal temperatures. There was significant proliferation of endogenous NSCs in the hippocampus after brain trauma at both temperatures, but NSC proliferation was slower at low temperatures. In addition, the expression level of Notch1 protein significantly increased in the hippocampus after brain trauma at both temperatures. However, at low temperatures, the degree of up-regulation of Notch signaling molecules was significantly lower.
Conclusion
In summary, we conclude from our results that low-temperature environments can inhibit the proliferation of the endogenous NSCs in the hippocampus, possibly by alleviating the effects of miR-34a down-regulation induced by TBI, which inhibits the reactive proliferation of endogenous hippocampal NSCs after trauma via the Notch signaling pathway. We believe that low-temperature environments may decrease the self-repairing capability of TBI patients and adversely affect the prognosis of TBI.
Footnotes
Acknqwledgements
This work was supported by the National Natural Science Foundation of China (81160154) and the National Key Technology R&D Program (2008BAI68B06).