
Abstract
A reliable animal model of facial nerve (FN) injury forms the basis for effective scientific studies of injury and repair of the FN. Currently, rodents are the most widely used animal model for such studies, most of which adopt a postauricular incision approach. However, it is difficult to carry out the procedure on perinatal rodents owing to characteristics of anatomy and the direction of incision. Based on anatomical studies on the extracranial segment of the FN in mice, we established a FN axotomy model by using a subauricular incision (SI) approach. The effect and the outcome of the FN transection were evaluated by electrophysiology, behavioral assessment and histological observation in 20 healthy four-week-old BALB/c mice. Favorable results were obtained in all of the mice and none died during the operation or subsequent observation period. Compared with the classic postauricular incision approach, the SI approach possesses multiple advantages including easier access, more satisfactory exposure of the FN trunk, minimal invasion and providing a larger operation field, which would be critical for those studies on the rodent in early development or delicate surgical manipulations.
Introduction
In humans, the facial nerve (FN) exits from the stylomastoid foramen (SF) of the petrous part of the temporal bone and then courses superficially. The FN is vulnerable to facial trauma or operation damages in adjacent regions, resulting in motor function disorders of the muscles of facial expression and a reduced quality of life in patients. 1 Therefore, the repair of FN injury has been extensively studied. These investigations mainly focused on the best way to prevent FN injury, 2 to promote functional recovery of the FN and the innervated muscles involved in facial expression,3,4 or further to repair and improve facial appearance. 5
To date, many studies have been performed to investigate FN injury, and various physical and chemical stimulus have been applied to observe the changes induced in FN axons and nuclei, as well as the related microenvironment.6–8 According to the different degrees of mechanical injury, the methods used to induce animal models of FN injury can be categorized as compression, 9 crush, 10 avulsion, 11 transection12,13 and resection. 14 Establishment of a reliable FN injury animal model is necessary for the development of scientific research related to functional recovery.
Because of the similarities between rodents and humans with regard to anatomical structures and pathophysiological reactions, the rodent has become the most widely used animal model for FN-related studies.11,15 The surgical technique adopting a postauricular incision approach to establish the facial resection model in the rodent, known as the ‘Kreutzberg model’, has been extensively reported.16,17 However, since the difficulties of performing this procedure on perinatal rodents owing to their anatomy characteristics and the direction of incision (see Discussion), most previous studies using the ‘Kreutzberg model’ involved adult rodents (≥8 weeks old).3,13,15
In this study, we investigated the anatomy of the extracranial segment of the FN in mice, and devised a mouse FN axotomy model using a subauricular incision (SI) approach. The animal model was systematically evaluated by electrophysiology studies, behavioral assessment studies and histomorphometric studies. Our purpose is to improve the FN injury model, so as to provide a promising addition to the traditional model for future studies involving the FN injury model, especially in young mice.
Materials and methods
Experimental animals and grouping
Twenty healthy four-week-old BALB/c mice (weighing 12–15 g, either sex, from Experimental Animal Center, the Second Military Medical University, Shanghai, China) were used, and were kept in four cages (five mice in each cage) with a 12-hour light/dark cycle having access to food and water ad libitum. The 20 mice were divided into a nerve resection group (NRG, N = 10) and a sham operation group (SOG, N = 10) using a table of random numbers. In both groups, the left-side FN was used for the operation, and the right side served as the control.
Surgical procedure
The animal was anesthetized by intraperitoneal injection of chloral hydrate (40 mg/kg). The operation region of each mouse was depilated by 8% Na2S solution and disinfected by iodine tincture. A 3-mm horizontal incision was made at the site 5 mm subauricular (Figure 1a). The subcutaneous fascia was dissected along the designated incision and the FN was exposed through cautious dissection along the anterior border of the cartilage of the external acoustic meatus (CEAM) (Figure 1b). In the NRG, the FN trunk was transected from the posterior border of the CEAM and a 3-mm length of the distal FN trunk was resected. Subsequently, the fascia tissue and skin were closed layer by layer with 3–0 silk sutures in the SOG and NRG. The animal care and the surgical protocols in this study were in strict accordance with the Rules for Animal Care and Use for Research and Education of the Second Military Medical University.
Photographs of the position of the incision (a), subauricular exposure of the facial nerve (b) and facial nerve function after nerve resection (c). (a) The horizontal incision (blue line) was about 3 mm long and 5 mm below the external acoustic meatus. (b) The exposed facial nerve trunk (black arrow) adjacent to the cartilage of the meatus after freeing the subcutaneous fascia from the primary incision. (c) View of a mouse from the nerve resection group immediately post-operatively: the tip of the nose is deviated to the right side, the left vibrissae are posteriorly positioned and immobile while the right ones are blurred because they are moving. (A color version of this figure is available in the online journal)
Electrophysiology
A multichannel physiological signal acquisition system (RM6240BD; Chengdu Instruments Co., Chengdu, China) was used in the electrophysiology study. Mice were evaluated immediately once finding the FN trunk in the operation region, after the operation (full exposure of the FN trunk in the SOG and nerve axotomy in the NRG), and before tissue harvest on postoperative day 21. The stimulating electrode was placed on the FN trunk, the recording electrode was embedded into the ipsilateral vibrissa-pad muscle, and the ground electrode was placed on the tail. Pulse stimuli (positive voltage, single stimulus, intensity 0.3 V and wavelength 0.5 ms) were applied three times to evoke compound muscle action potentials (CMAPs).
Behavioral observations
Experimental animals were observed preoperatively and at postoperative day 1, 3, 7, 14 and 21. The following parameters were evaluated: orientation of the tip of the nose, open and closed amplitude of bilateral rima oculi, blink reflex and vibrissa swing (Table 1).
Facial nerve function in mice

The blink reflex in the mouse was detected by air current stimulation of the cornea. Each mouse was placed in a fixed position, and a 10-mL syringe connected with a 26-gauge needle was used to rapidly push a volume of 5-mL air onto the cornea at the location of 2 cm before the eyes. A digital camera (Caplio G × 100; Ricoh, Tokyo, Japan) was used to obtain videos of blink reflex evaluation.
The animals were placed in a box (18 cm × 16.5 cm × 10 cm) with black-painted inner walls in a quiet environment at room temperature. The same digital camera was used to videorecord two minutes of the animals’ activity at about 8:00 on each of the abovementioned time points from the top of the box. The static position of the vibrissa as well as the amplitude and frequency of its swing were observed.
According to the scoring system developed by de Faria et al. 18 for the rat model of FN axotomy, combined with the methods reported by Farrag et al. 19 and Bodjanac et al., 20 the normal side was set as a control, and the blink reflex and vibrissa swing of the operated side were each classified into five grades (ranging from 0 to 4, see Table 1). The FN functions of mice in the two groups were evaluated and analyzed by an evaluator who was double-blinded to this study. All scores for each of the two parameters in each group were summed for assessment of the FN function.
Tissue harvesting and processing
On the 21st day following surgery, the animals were anesthetized again by intraperitoneal injection of chloral hydrate (40 mg/kg) and were transcardially perfused with 0.9% NaCl (40 mL) followed by 4% paraformaldehyde in 0.1 M phosphate-buffered saline (pH 7.4, 30 mL). The brains were taken out, and fixed for four hours in the same fixative, then cryoprotected overnight in 30% sucrose (4°C). In accordance with The Mouse Brain in Stereotaxic Coordinates (2nd edn), 21 the FN nuclei were serially cryosectioned at a thickness of 20 μm (CM1900; Leica Geosystems, Heerbrugg, Switzerland).
Every fifth section was incubated with 3% normal goat serum in phosphate-buffered saline for 30 min to block nonspecific binding sites. Subsequently, sections were incubated overnight at 4°C with a mouse antineuronal nuclei (NeuN) monoclonal antibody (1:200; Chemicon, Millipore, Billerica, MA, USA) followed by corresponding secondary antibody (CY3-labeled goat anti-mouse IgG [A0521; Beyotime, Shanghai, China]) and Hoechst 33258 (861405; Sigma, St Louis, MO, USA) incubation. The sections were embedded in glycerol supplemented with antifade reagent (N-propyl-galat). Histomorphometric studies of brain tissue cryosections were carried out with a Nikon microscopy (Eclipse E600, DXM 1200, × 100; Nikon, Tokyo, Japan) and pictures taken by Nikon ACT-1 software (version 2.63). According to the criteria reported by Park et al., 22 the cells we identified as the facial motoneurons (FMNs) were those localized in the facial nucleus with strong fluorescence of NeuN expression, and a distinct cell nucleus.
The neurons localized in the FN nucleus of each section were then counted by two observers who were blinded to this study. The FMN count of each side was multiplied by five for an estimation of the total FMN count.
Results
All the experimental animals survived the operation. Surgical incision healing was uneventful, without infection or hematoma. No bite or scratching was observed during the postoperative follow-up.
Topographic anatomy of the FN in mice
Shortly after its emergence from the SF, the FN trunk travels anteriorly under the clavotrapezius and sternomastoid muscle (SM).
18
Consequently, this results in difficultly in exposing the FN trunk without transecting the muscle tendons at that site in perinatal mice. When the FN trunk reaches the posterior border of the CEAM, it travels over the cartilage surface, enters into the parotid tissue from the deep surface of the parotid gland (PG) and branches under it. The FN could easily be found near the SI, free of any other transverse nerve. Both the anterior auricular vein (AAV) and superficial temporal vein (STV) run vertically and superficially across the FN trunk near the anterior border of the CEAM and should be protected during the mobilization
23
(Figure 2).
Drawing of the topographic anatomy of the facial nerve and the surgical approaches for facial nerve axotomy in mouse. FN, facial nerve; SI, sub-auricular incision; PI, postauricular incision; AAV, anterior auricular vein; STV, superficial temporal vein; SM, sternomastoid muscle; CM, clavotrapezius muscle; SF, stylomastoid foramen; CEAM, cartilage of the external acoustic meatus; PG, parotid gland. (A color version of this figure is available in the online journal)
Electrophysiology
Before the FN axotomy, an evident M-shape CMAP could be invoked by electrical stimuli in the vibrissa-pad muscles of mice in the NRG. However, CMAP could not be evoked immediately after FN axotomy or three weeks later (Figures3a–c). Incontrast, the CMAP waves evoked immediately after FN exposure and three weeks later were similar to those observed before the operation (Figures 3d–f).
Evoked CMAPs of vibrissa-pad muscle in the facial nerve resection group (NRG, a–c) and the sham operation group (SOG, d–f). (a, d) Immediate response when finding the facial nerve trunk, (b) immediately after nerve axotomy and (e) immediately after full exposure of the facial nerve trunk. (c, f) Response before tissue harvest on postoperative day 21 in the NRG and SOG, respectively. CMAP, compound muscle action potential; NRG, nerve resection group; SOG, sham operation group. (A color version of this figure is available in the online journal)
Behavioral observations
In mice from the SOG, there was no postoperative deviation in the orientation of the tip of the nose. The bilateral blink reflex and swing of the vibrissa were normal except for the occurrence of a mild, transient reduction in swing amplitude in three mice during the first week postoperatively. However, in mice from the NRG, the tip of the nose deviated to the right and the blink reflex of the left eye completely vanished at postoperatively day 1. The left vibrissa was in a static state, without any swings or movement, in contrast to the situation observed for the right vibrissa (Figure 1c). On postoperative day 14, the left vibrissa of two mice in the NRG showed slight tremor. This number increased to three on postoperative day 21. FN function scoring results indicated that the operation procedure of FN exposure had no or mild effects on FN function, whereas the complete set of facial palsy signs appeared in the NRG after the operation. The vibrissa swing recovered slightly in a minor fraction of mice during the 14–21 days after the operation, with the amplitude degree scored as 1 according to the FN function scale (Table 2).
Scores of facial nerve function for mice in two groups at each time point
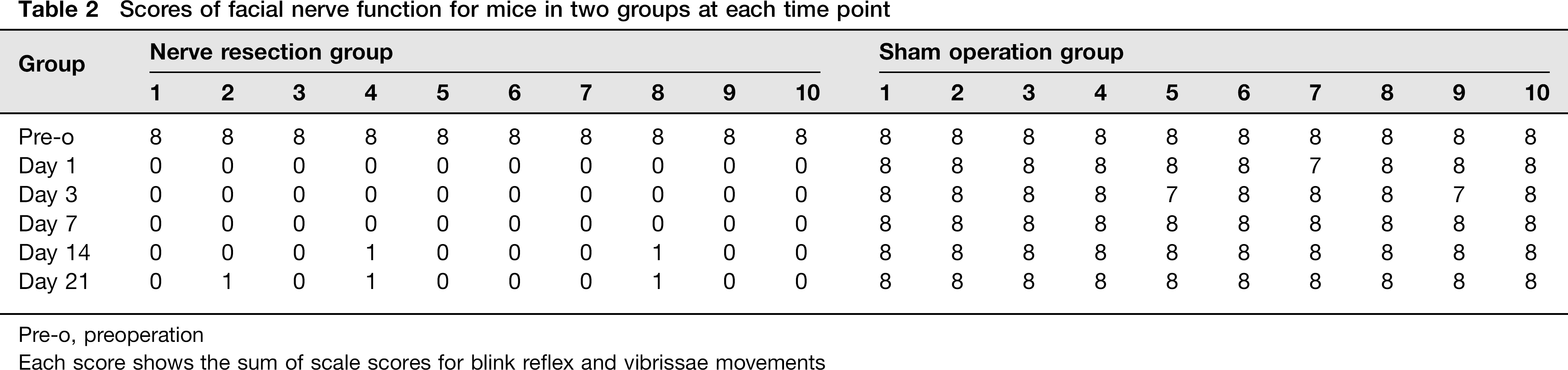
Pre-o, preoperation
Each score shows the sum of scale scores for blink reflex and vibrissae movements
Counts of FMNs
We compared the effects of two different treatments on FMN death in this study. Three weeks after the operation, the number of surviving FMNs was quantitatively analyzed via immunofluorescence. In the NRG, the left side exhibited a ~20% loss of FMNs, compared with the opposite side (t = 11.36, P < 0.05). In contrast, both sides of the SOG displayed no or minimal loss of FMNs (t = 1.198, P = 0.262) (Figure 4).
Differential facial motoneuron (FMN) loss three weeks postoperatively. (A) Photomicrographs of differential FMN remnants in the nerve resection group (a, b; NRG, n = 10) and sham operation group (c, d; SOG, n = 10) (NeuN immunofluorescence). In the NRG, the FMNs (red) on the left side were more sparse than the right side. Such difference was not seen in the SOG. L, left side; R, right side. The motoneurons were labeled by red and the nuclei were labeled by blue. Dotted lines indicate the boundary of the facial motor nucleus. Bar = 200 μm. (B) Statistics and histograms of counts of the number of surviving FMNs three weeks postoperatively in the NRG (n = 10) and SOG (n = 10). The counts of the number of surviving FMNs on the left (L, operation) and right (R, control) side of the two groups are included. In the NRG, compared with the opposite side, the left side exhibited a ~20% loss of FMNs (t = 11.36, P < 0.05). In contrast, both sides of the SOG displayed no or minimal loss of FMNs (t = 1.198, P = 0.262). Data are mean ± SEM. *P < 0.05 in Student's t-test comparisons. (A color version of this figure is available in the online journal)
Discussion
Since the FN axotomy model in rats was successfully established by Georg Kreutzberg, this animal model has been extensively used in related studies of various injuries and repair mechanisms of the FN.16,17 This model came to be known as the ‘Kreutzberg model’ and was recognized as a classic FN injury model. 24 The ‘Kreutzberg model’ adopts a postauricular incision approach and the FN is exposed forward from the location of the SF 18 (Figure 2). The position of the FN exiting from the SF is relatively fixed, which allows easy location of the nerve. However, the post-auricular incision is not consistent with the running-course orientation of the FN and near to the clavotrapezius and SM tendon attachments. 18 So it is difficult for postauricular incision approach to get a satisfactory exposure of the nerve in those smaller rodents, such as mice in the perinatal period, or to obtain enough space for delicate surgical manipulation in the operative field (e.g. nerve resection or anastomosis) through the postauricular incision approach. Consequently, the operator might always have to extend the incision, perform another auxiliary incision or cut-off the adjacent tendon, each of which will undoubtedly increase the invasiveness of the operation and may therefore lead to a higher operative mortality, especially in perinatal mice because of its vulnerability to trauma.
Compared with the classic postauricular incision, the SI might solve these abovementioned problems. We designed the SI because from our previous study, it was found that the subauricular segment of the FN traveled horizontally from the postauricular side to the rostral side, overstrode the CEAM and then entered the PG (Figure 2). In the sub-auricular area, there are no bulging muscles or large vascular and nerve bundles covering the FN trunk. A satisfactory exposure of the FN could easily be acquired through superficially incising the skin and subcutaneous fascia, and pushing a small posterior part of the PG towards the rostral side, which cover the anterior part of the subauricular FN trunk. The complicated intramuscular dissection or muscle tenotomy is avoided. Furthermore, since the incision we designed is parallel with the running orientation of the FN trunk (Figure 2), the length of the exposed FN can simply be adjusted by changing the length of incision. In brief, the relatively fixed running course of the subauricular FN, the incision with superficial dissection, rare adjacent tissue and the flexibility in the adjustment of the exposed FN length made it feasible for dissecting a longer FN trunk with an easier manipulation through the SI approach. This meant that the operative trauma and the surgical duration, which are important determinants of operative mortality in experiment animals, would be reduced effectively.
In this study, we also found another distinct advantage associated with a special anatomical structure in the sub-auricular area – the CEAM, which played an important role throughout the operation. First, the CEAM is strongly suggested as an anatomical marker during the exploration and dissection of the FN. When entering the surgical zone through the SI approach, the cartilage is commonly the first tissue explored, the reasons of which are as follows: (1) the cartilage can be easily located according to the position of the external ear; (2) the cartilage can be easily distinguished from the adjacent tissues according to its histological characteristics (large, white and tough); (3) the cartilage can be easily dissected because its connection with the surrounding tissues is loose. Since the FN is the only nerve transversely travelled on the cartilage, after pushing the PG aside, it can be sufficiently exposed on the cartilage. During the dissection of the FN trunk in the rostral direction, it is important to protect both the AAV and STV in the region close to the anterior border of the cartilage. The blood vessels in this position crossed perpendicularly and superficially with the FN trunk. It was important to be very cautious and patient when dissecting in this region to avoid the occurence of vessel rupture and hemorrhage, which not only can affect visualization of the operation field, but also induce the death of mice by blood loss. The other important role of the cartilage is providing a rigid platform for operation. For the subauricular segment of the FN trunk mainly traveled over the CEAM, the whole course of nerve dissection can be performed on the cartilage. The relatively large and tough characteristics of the cartilage provide enough space and rigid basement for the dissection. All these factors are also beneficial for further delicate surgical manipulation, in which the ‘Kreutzberg model’ is limited.
We performed operative intervention on 20 four-week-old mice using this operative incision approach, established the FN axotomy model in perinatal mice and acquired favorable results. According to the assessment results from the electro-physiology, it was verified that the paralysis of the vibrissa-pad muscle caused by nerve axotomy could last at least three weeks until being reinnervated. From the results of behavioral observation in the postoperative period, the efficacy of the FN axotomy and the minimal influence of the FN-exposed manipulations were reconfirmed. Moreover, the reductions of FMNs in the FN nucleus sections were found, which further proved the availability of our model. The results in the SOG also showed that the dissection of the FN caused minimal pathological lesion of the FN and had little or no FN functional damages. Therefore, the FN axotomy model via SI approach is reliable. Furthermore, clinical data showed that the segment of FN near to the PG is one of the regions most easily injured in surgical treat-ments.25,26 For this reason, it might be an ideal pathological model for this kind of injury.
In the later period of the experiment, three mice in the NRG exhibited slight swings of the vibrissa, but did not evoke a muscle action potential during the subsequent electrophysiology evaluation. Therefore, after precluding the possibility of reinnervation of the vibrissa-pad muscle, we considered that this phenomenon may be concerned with synkinetic facial movement.27,28
Conclusion
We provided a new method for establishing the FN axotomy model in perinatal mice, and this novel approach involves less tissue injury because the FN is easier to explore and can also provide a larger space for surgical manipulation. This operation approach could be adopted for those studies on the rodent in early development with poor tolerance of operation (e.g. perinatal mice) or with delicate surgical manipulations (e.g. microneurorrhaphy, nerve transplantation).
Footnotes
Acknowledgements
This work was supported by the Special Project of the ‘Eleventh Five-year Plan’ for Medical Science Development of PLA (grant no. 08G069) and the Key Basic Research Foundation of Shanghai China (grant no. 08JC1407100).