
Abstract
Acute liver failure (ALF) is characterized by severe neurological complications, known as acute hepatic encephalopathy, where brain ammonia and inflammatory processes play a dominant role. In experimental models of acute liver failure SERPINB3 was found significantly increased in microglia, the intrinsic immune cells of the central nervous system. The aim of the present study was to investigate the extent of brain tissue damage and the inflammatory milieu in experimental acute liver failure using a SERPINB3-transgenic mouse model. C57BL/6J wild-type and transgenic mice were inoculated with acetaminophen or phosphate-buffered saline and sacrificed 20 h postinjection. Proliferation and apoptotic activity were analyzed in brain tissue by immunohistochemistry and terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling technique. The expression of cytokines was analysed in brain and liver tissue by real time polymerase chain reaction and in the corresponding serum samples using a Bio-Plex system. Acetaminophen induced a significantly lower body temperature and shorter survival in transgenic than in wild-type mice, despite liver function was similar in both groups. The brain of transgenic mice, expressing SERPINB3 positivity in microglia, showed increased glial cell number, associated to significant lower apoptotic death events, compared with wild-type mice. In mice injected with acetaminophen, remarkably higher values of cytokines mRNA were observed in the liver of both groups, with a trend toward higher values in transgenic animals. In brain tissue similar increase of tumor necrosis factor-αwas detected in transgenic and wild-type mice, while IL-10 mRNA increased only in the wild-type group. A remarkable increase of circulating Th1 cytokines was detected in serum of transgenic mice, while in the wild-type group they remained rather unchanged. These figures were associated with lower levels of granulocyte macropage colony-stimulating factor, despite similar increase of IL-10 values in both groups. In conclusion, in acute liver failure SERPINB3 determines an enhanced inflammatory background, mainly mediated by higher levels of Th1 proinflammatory cytokines.
Keywords
Introduction
Acute liver failure (ALF) is a condition with an unfavorable prognosis. Encephalopathy, brain edema and intracranial hypertension are serious neurological complications responsible for mortality in patients with ALF. 1 Several factors contribute to the cerebral complications of ALF and the genesis is most likely multifactorial.2,3 Brain edema, hyper-ammoniemia and systemic inflammation play a pivotal role in cerebral alterations.4–6 Different mechanisms in specific regions of the brain have been shown to contribute to the progression of cerebral alterations and to the generation of intracranial pressure. Early increase of blood– brain barrier permeability in certain areas, followed by vasogenic edema, determines initial abnormal intracranial pressure. 7 The metabolism of ammonia in astrocytes than induces an increase of glutamine and lactate that mediate cellular swelling, as a direct cytotoxic mechanism. 8 Alterations in lactate and cerebral blood flow are later events that further increase intracranial pressure. 7 Several studies suggest that also inflammation plays a significant role in the pathogenesis of acute hepatic encephalopathy. The presence of microglial activation, systemic and brain inflammation are associated with a more rapid progression of encephalopathy and brain edema in ALF patients 9 and in rats with ALF resulting from hepatic devascularization. 10 Systemic inflammation in ALF determines increased circulating levels of proinflammatory cytokines, including tumor necrosis factor alpha (TNF-α) and interleukins (IL)-1β and IL-6 11 released by the necrotic liver, but also produced by the brain. 12
The knowledge of the sequence of cerebral events involved in ALF has improved recently and in addition to inflammation, other factors have also been involved. Matrix metalloproteinase (MMP)-9, implicated in various brain injuries, has been shown to contribute to the pathogenesis of brain extravasation and edema. 13 It is of interest to note that MMP-9 can be induced by the serpin SERPINB3 14 and using reverse transcription-polymerase chain reaction (RT-PCR) differential display Thakker– Varia et al. 15 have detected a remarkable increase of SERPINB3 expression in lipopolysaccharide (LPS)-activated microglial cultures. In addition, this serpin was able to increase microglial cell number. SERPINB3 (known as SCCA1, squamous cell carcinoma antigen) belongs to the ov-serpin family, serine protease inhibitors involved in multiple biological functions and cell homeostasis control. 16 Two isoforms (SERPINB3 or SCCA1 and SERPINB4 or SCCA2) were originally purified from squamous cell carcinoma of uterine cervix. 17 Genomic cloning and sequencing revealed that they were 91% identical at the amino acid level, but showed distinct properties and substrates 18 and their physiological roles are poorly understood. While in normal conditions SERPINB3 is expressed in epithelia,19,20 in sweat glands, in endothelial cells of the veins, in arteries walls 21 and in peripheral blood mono-nuclear cells, 22 its over-expression has been described in neoplastic cells of epithelial origin 20 and in preneoplastic and neoplastic liver lesions, but it is not detectable in normal hepatocytes.21,23
The aim of the present study was to assess the SERPINB3 influence in experimental ALF induced by acetaminophen using a SERPINB3-transgenic mouse model.
Materials and methods
Experimental acute liver failure model
The study was carried out in C57BL/6J mice transgenic for human SERPINB3 (kindly provided by Professor G. Cassani, Technogen S.c.p.A, Piana di Monte Verna, CE, Italy), and in C57BL/6J wild-type mice (Charles River Italia S.p.A, Calco, Lecco, Italy), aged 12–14 weeks. The transgenic mice have been shown previously to express SERPINB3 mRNA in the brain, lung and liver 10–20 times higher than in kidney and muscle, where the expression was trivial (<0.6 pg/μg RNA). 24 The mouse-homologous Serpinb3 mRNA was not detectable in the same tissues in transgenic and wild-type mice. 24 By immunohistochemistry SERPINB3 protein was documented in transgenic animals in the hepatocytes in the liver (Please see Supplementary Figure S1 available online at http://ebm.rsmjournals.com/lookup/suppl/doi:10.1258/ebm.2012.012135/-/DC1), while in the lung it was mainly detected in the cytoplasm of inflammatory cells (mainly macrophages). 25
Mice were maintained under a 12 h light/dark cycle at 21°C±1°C and bred at the Animal Care Facility of the Experimental Surgery Division of the University of Padua. In preliminary experiments, to assess the optimal dose to induce ALF, a group of 10 mice (5 transgenic and 5 wild-type) were inoculated intraperitoneally with increasing concentrations of acetaminophen and a concentration of 0.8 mg/g body weight was chosen for further experiments. An additional group of 10 mice (5 transgenic and 5 wild-type) was inoculated with the optimal dose of acetaminophen and mice were observed until death. High doses of acetaminophen are potentially fatal, 26 because acetaminophen, converted in the liver to a reactive quinone form (N-acetyl-p-benzoquinone imine) 27 by cytochrome P-450 enzymes, covalently binds to cellular macromolecules producing reactive oxygen species. 28
To carry on the study, 24 mice (12 transgenic and 12 wild-type) were inoculated with acetaminophen and an identical control group was inoculated with phosphate-buffered saline (PBS) solution. All mice were sacrificed 20 h postinjection. Body temperatures were controlled throughout the experiment.
After sacrifice, at least two tissue samples were taken from the brain and the liver of each animal: the first one was formalin fixed and paraffin embedded, while the other was immediately frozen in liquid nitrogen and stored at -80°C until use.
The experimental protocol was approved by the Animal Investigation Committee of the Italian Ministry of Health and all measures were taken to minimize any pain or discomfort of the animals.
Immunohistochemistry
SERPINB3 expression
All brain specimens were immunostained with a polyclonal rabbit antiSERPINB3 antibody (Hepa-Ab, Xeptagen, Marghera-VE, Italy) as previously described.25,29 Samples were processed using a sensitive avidin-streptavidin-peroxidase technique and stained with a mixture of 3,3-diamino-benzidine tetra hydrochloride and hydrogen peroxide. To detect any non-specific staining, control slides were prepared lacking primary antibody, lacking primary and secondary antibodies or stained with normal sera.
Proliferation and apoptosis evaluation
In treated mice 20 h after injection, proliferation and apoptotic activity were analyzed in the glial cell compartment of brain tissue identified by immunohistochemistry performed using the primary monoclonal antibody anti-CD68 (AbD Serotec, Oxford, UK).
Four-μm-thick sections were cut and processed for immunohistochemical analysis of Ki-67 to detect proliferating cells. Briefly, after dewaxing and hydration, sections were incubated in citrate buffer 5 mmol/L at pH 6.0 in a microwave oven for 30 min for antigen retrieval. Afterwards, sections were treated with normal serum (Immunotech, Marseille, France) and incubated for 60 min with the primary monoclonal antibody anti-Ki67 (MIB-1, Genetex, Irvine, CA, USA) at a concentration of 1:100. Sections were subsequently incubated with rabbit horseradish peroxidase (HRP) polymer (Dako, Glostrup, Denmark) for 30 min. Immunoreactivity was visualized with 3-3’-diaminobenzidine (Dako). Negative controls for non-specific binding were processed omitting the primary antibody and revealed no signal. Ki-67 immunostaining was randomly evaluated counting at least 100 nuclei of glial cells. The percentage of positive nuclei was expressed as the proliferative index (PI).
To investigate the presence/extent of apoptosis, the terminal deoxynucleotidyltransferase-mediated dUTP-biotin nick end-labeling (TUNEL) method was used. Sections were processed in accordance with Gavrieli's method. 30 Briefly, after deparaffinization and re-hydration, sections were digested with proteinase K (Boehringer, Mannheim, Germany) at a concentration of 20 μg/mL for 15 min. The slides were then incubated with TdT/biotinylated dUTP diluted in buffer (Boehringer) and developed by using 3-3’ -diaminobenzidine and 30 mL hydrogen peroxide. For negative controls some slides were incubated in buffer without TdT or biotinylated UTP. For positive controls some slides were incubated with 1 μg/mL DNAse (Sigma-Aldrich, Milan, Italy). The TUNEL positivity was randomly evaluated counting at least 100 nuclei of glial cells. The percentage of positive nuclei was expressed as the apoptotic index (AI). All the images were acquired by a microscope (Axioplan 2, Zeiss, Germany) equipped with the Image Pro-plus V.5 (Media Cybernetics, Silver Spring, MD, USA) morphometrical analysis system.
Quantitative realtime PCR (SYBR green assay) for cytokine expression
Cytokine expression (TNF-α, IL-6 and IL-10) was assessed in brain and in liver tissue by realtime PCR. Total RNA was extracted from frozen tissue using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Two micrograms of extracted RNA for each tissue sample were reverse transcribed into cDNA first-strand by First Strand cDNA Synthesis kit, using 200 U of SuperScript II Reverse Transcriptase and 0.5 μg of oligo dT (Invitrogen). The SYBR Green realtime PCR was performed with a Light Cycler Apparatus (Roche, Monza, Italy), using a FastStart DNA MasterPLUS SYBR Green KitTM (Roche, Monza, Italy) in glass capillaries. Primers were designed from the published sequences of mouse TNF-α mRNA (Accession number GenBankTM NM_013693.2), mouse IL-6 mRNA (Accession number GenBankTM NM_031168.1), mouse IL-10 mRNA (Accession number GenBankTM NM_010548.1) and mouse β-Actin mRNA (NM_007393.2). The nucleotide sequences were as follows:
TNF-αsense: 5’ AGCCCCCAGTCTGTATCCTT 3’; TNF-α antisense: 5’ CTCCCTTTGCAG AACTCAGG 3’; IL-6 sense: 5’ AGTTGCCTTCTTGGGACTGA 3’; IL-6 antisense: 5’ CAGA-ATTGCCATTGCACAAC 3’; IL-10 sense: 5’ CCAAGCCT-TATCGGAAATGA 3’; IL-10 antisense: 5’ TTTTCACAGGGGA-GAAATCG 3’; β-actin sense: 5’ AGCCATGTACGTAGCCA TCC 3’; β-actin antisense: 5’ CTCTCAGCTGTGGTGGT-GAA 3’.
Amplification of specific transcripts was confirmed by melting curve profiles at the end of each PCR cycle, using the specific routine built-up in the Light Cycler instrument. A final extension step was performed at 37°C for one min. The relative quantification of the transcripts of each gene of interest was normalized by the housekeeping gene (β-actin).
Serological assays
Serum alanine transaminase (ALT) concentration was achieved by laboratory routine assay. Ammonia (NH3) was measured in serum by the Ammonia Checker IITM instrument (Kyoto Daiichi Kagaku Co Ltd, Kyoto, Japan) according to the manufacturer's instructions.
A panel of mouse cytokines (IL-1β, TNF-α, IFN-γ, IL-2, IL-4, IL-5, IL-10 and granulocyte macropage colony-stimulating factor [GM-CSF]) was analyzed in plasma samples of both transgenic and wild-type mice obtained before and after acetaminophen injection, using the Bio-Plex protein assay system (Bio-Rad Laboratories, Hercules, CA, USA) following manufacturer's instructions and the results were analyzed by X-Map technology (Luminex, Mirai Bio, Alameda, CA, USA).
Statistical analysis
Statistical analysis was carried out using the Student's t-test, the non-parametric Mann-Whitney U test, Wilcoxon matched pairs test and unpaired t-test with Welch's correction, when appropriate. All tests were two-tailed. The significance was set as P < 0.05. All analyses were performed using GraphPad InStat software (San Diego, CA, USA).
Results
Mouse behavior after acetaminophen injection
Twenty hours after acetaminophen injection all 10 initially treated mice displayed signs of hepatic encephalopathy, such as trembling, reduced locomotive activity, ataxia and lethargy. Transgenic mice showed significantly shorter survival time than wild-type mice (mean ±SD: 29.2 ± 2.6 h versus 50.4 ± 5.9 h, P = 0.0159) (Figure 1). In the additional group of 24 acetaminophen-injected mice, 20 h after treatment, body temperature showed a more marked decrease in the transgenic group than in controls (mean + SD: 27.4 ± 1.4°C versus 30 ± 1.6°C, P = 0.0152). At biochemical level normal values of transaminase and ammonia were detected in mice injected with PBS (ALT median ± SEM: transgenic = 4.38 ±1.4 U/dL, wild-type 4.38 ± 0.7 U/dL, P = ns; ammonia median ± SEM: 73.00+9.5 μg/dL versus 96.00+33.7 μg/dL, P = ns). In acetaminophen-treated mice increased values of both parameters were detected, although a trend toward lower ALT values was observed in the transgenic mice group, compared with wild-type mice (median ± SEM: 621.25 ± 688.5 U/dL versus 1129.25 ± 1493.5 U/dL, P = ns). Values of ammonia were similar in both groups (median ± SEM: 222.00 ± 25 μg/dL versus 236.50 ± 50.6 μg/dL, P = ns), as shown in Figure 2 (left panels). At histology both transgenic and wild-type mice showed widespread hepatocellular necrosis, mainly localized in centrolobular region. In periportal areas swollen hepatocytes and mild-moderate inflammatory infiltrate was observed (Figure 2, right panels).
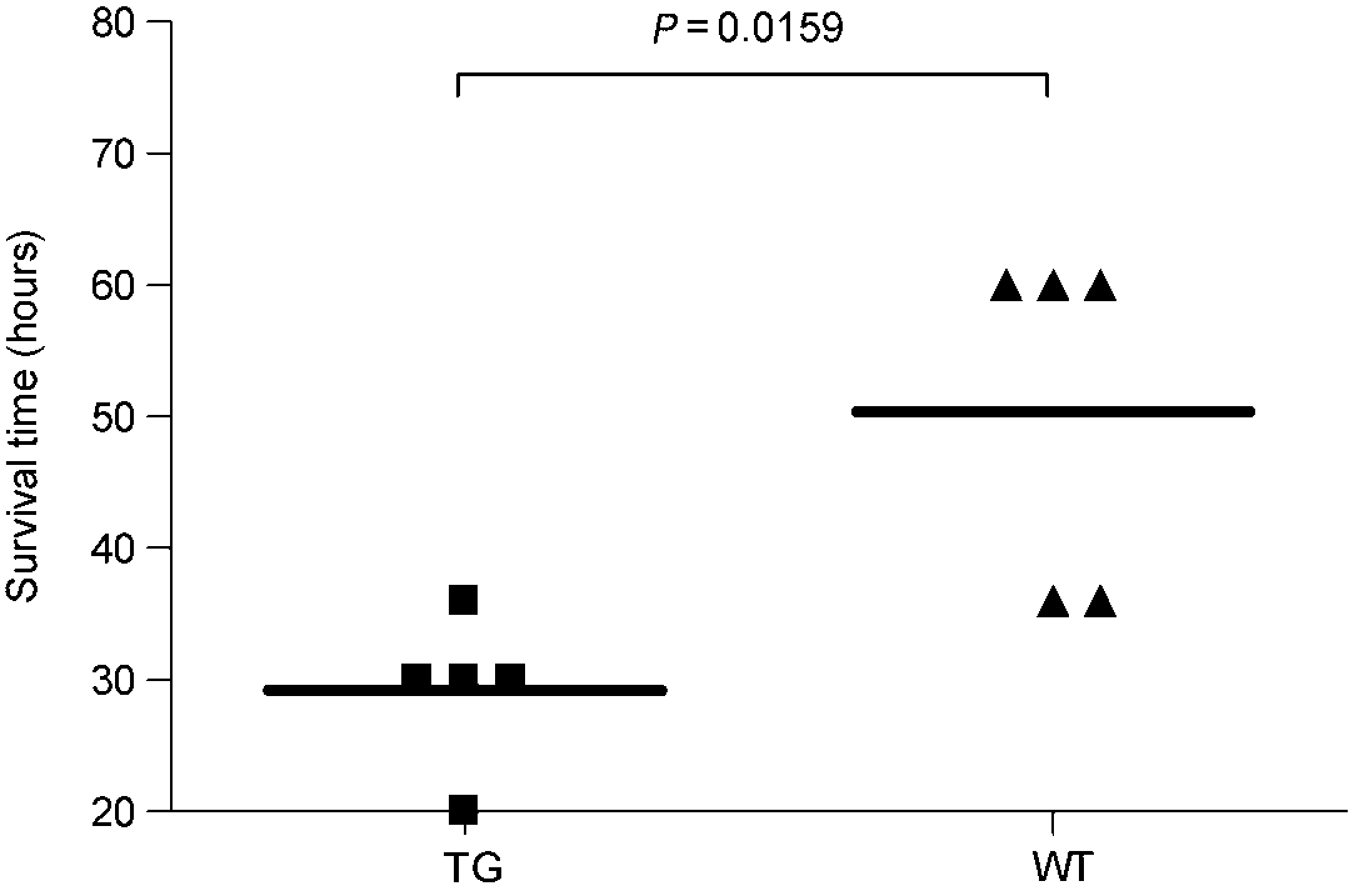
Survival time in transgenic and wild-type mice after acetaminophen injection. Transgenic mice (TG) showed a significantly shorter life time than the matched group of wild-type (WT) mice. Horizontal bar represent median values
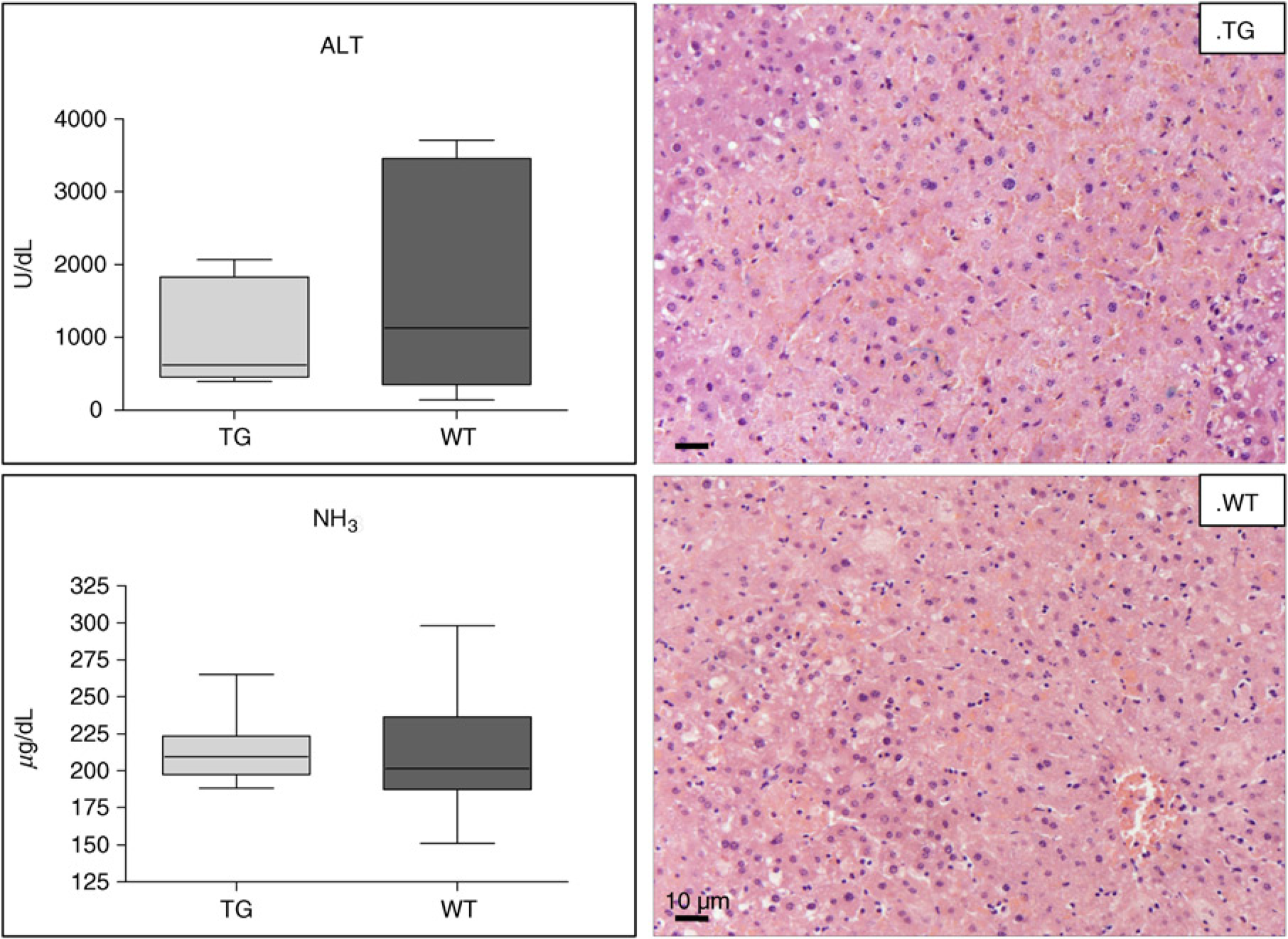
Biochemical parameters in treated mice. Box plot of serum values of alanine transaminase and ammonia (NH3) in transgenic (TG) and wild type (WT) mice after acetaminophen injection. The box indicates the lower and the upper quartile and the middle line indicates the median. Bars indicate the range of values distribution. Swollen hepatocytes, congestion and mild granulocyte infiltration are well seen in both TG and WT mice (hematoxylin-eosin staining in liver, original magnification: × 200). (A color version of this figure is available in the online journal)
Liver tissue analysis
Cytokine mRNA expression in liver tissue of mice injected with PBS showed a trend toward higher values of TNF-α, IL-6 and IL-10 in the transgenic group compared with wild-type mice (TNF-α: 2-ΔΔCT median ± SEM = 1.75 ± 1.1 versus 0.71 ± 0.60, P = ns; IL-6: 2-ΔΔCT median ± SEM = 1.27 ± 0.53 versus 0.83 ± 0.58, P = ns; IL-10: 2-ΔΔCT median ± SEM = 1.80 ± 1.05 versus 0.80 ± 1.25, P = ns). In mice injected with acetaminophen remarkably higher values of cytokines were observed in both groups, with the persistence of a trend toward higher values in transgenic animals (Figure 3b).
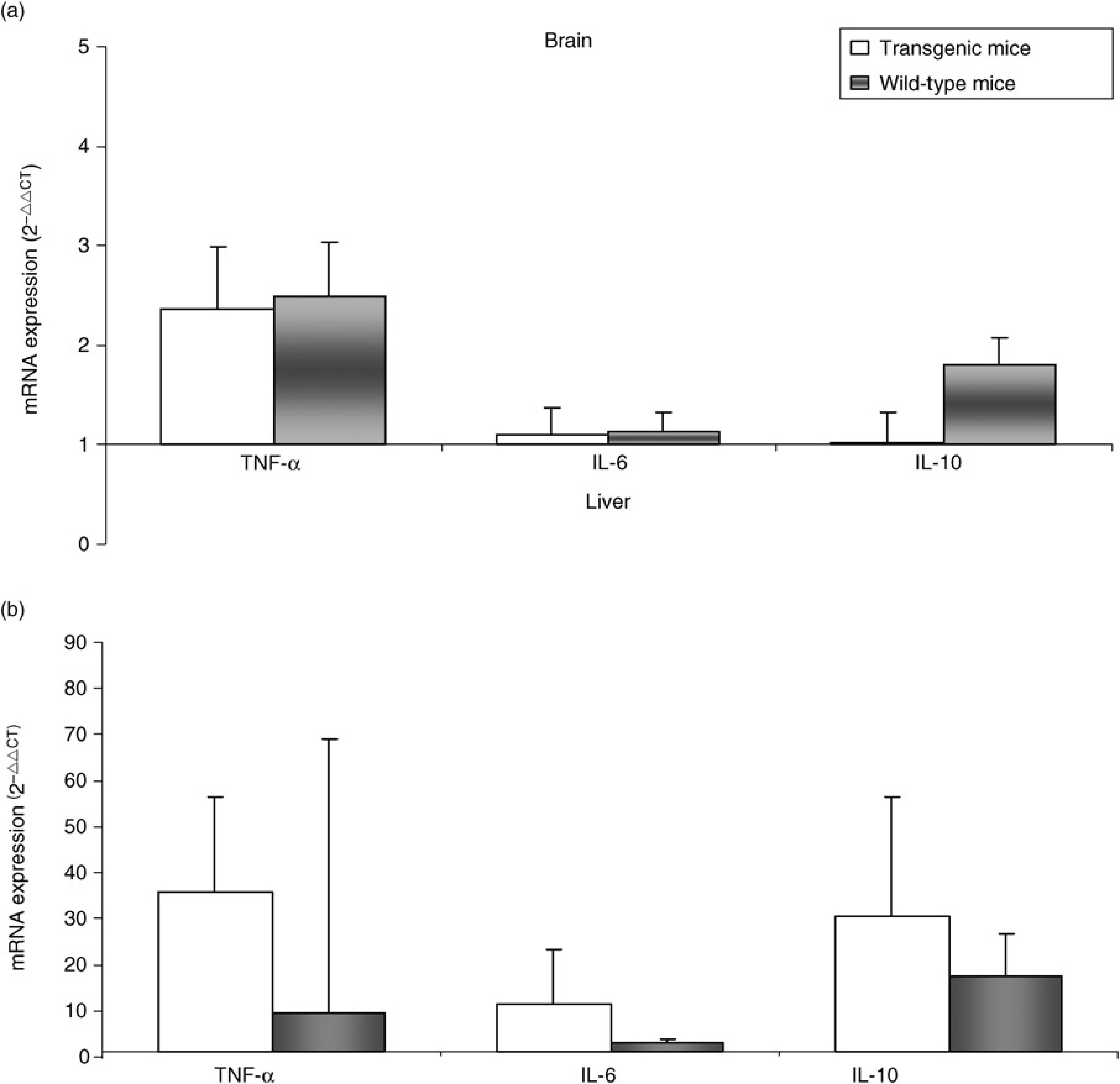
Cytokine mRNA expression in brain and liver tissue. Median expression of cytokines in brain (a) and liver (b) tissue of acetaminophen treated transgenic and wild-type mice, detected by realtime polymerase chain reaction. Changes in mRNA gene expression were reported as fold increase relative to controls (animals treated with saline solution) using the 2-ΔΔCTmethod. The results were normalized to the β-actin housekeeping gene. Vertical bars represent standard error median
Brain tissue analysis
Immunohistochemistry documented the presence of SERPINB3 in vascular endothelial cells in brain tissue of both transgenic and wild-type mice. In addition, transgenic mice showed SERPINB3 reactivity also in microglial cells and these features were detected in mice with and without ALF (Figure 4). An increased number of glial cells was also detected in the brain of transgenic animals, independently of acetaminophen treatment. These results were supported by the finding of a reduced glial cell AI, compared with the control mice group (mean ± SD, 2.0 ± 1.6% versus 7.0 ± 6.7%, P = 0.0276) (Figure 5), despite a similar PI (mean ± SD: 33.1 ± 13.5% versus 22.7 ± 17.3%, P = ns).

Immunohistochemical staining for SERPINB3 in brain tissue in mice with or without acetaminophen treatment. SERPINB3 expression was clearly detectable in vascular endothelial cells (thick arrow) and microglial cells (thin arrow) in transgenic mice, while it was detectable only in vascular endothelial cells in wild-type mice. Moreover, an increased number of glial cells is well seen in transgenic mice, independently of acetaminophen treatment. Original magnification: ×20. (A color version of this figure is available in the online journal)

Apoptosis in glial cell compartment. (A) Box plot of glial cell apoptotic index (%) in brain tissue of transgenic (TG) and wild-type (WT) animals after saline (control) or acetaminophen injection. The box indicates the lower and the upper quartile and the middle line indicates the median; bars indicate the range of values distribution. A higher number of apoptotic nuclei are seen in WT (C) than in TG (B) mice (terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling technique, original magnification: ×400). (A color version of this figure is available in the online journal)
Transcription analysis of cytokines in brain tissue documented a similar increase (about 5 times) of TNF-α in injured mice (Figure 3a), compared with the values of PBS-treated mice (2-ΔΔCT median ± SEM: transgenic = 0.58 ± 0.01, wild-type = 0.56 ± 0.01, P = ns). IL-6 levels were very low in mice without ALF (2-ΔΔCT median ± SEM: transgenic = 0.47 ± 0.01, wild-type = 0.46 ± 0.02, P = ns) and this finding remained unchanged in acetaminophen-treated mice. IL-10 expression was very low in mice without ALF (2-ΔΔCTmedian ± SEM: transgenic = 0.48 ± 0.02, wild-type = 0.46 ± 0.01, P = ns) and among the animals injected with acetaminophen, it increased about four times in wild-type mice, but not in the transgenic group (Figure 3a).
Cytokine profiles in serum
The behavior of circulating cytokines was different in trans-genic and wild-type mice, as shown in Figure 6. A remarkable increase of circulating Th1 cytokines, especially of IL-1β, was detected in serum of transgenic mice treated with acetaminophen, compared with controls, while in the wild-type group these cytokines remained rather unchanged, independently of acetaminophen treatment. GM-CSF values tended to be more elevated in the trans-genic group, compared with wild-type animals, where a significant drop was detected in mice with ALF. A significant increase of IL-10 values was observed in both groups treated with acetaminophen.

Cytokine levels in serum of mice treated with acetaminophen or with phosphate-buffered saline solution (Control). Panel (a) refers to transgenic mice and Panel (b) refers to wild-type mice. Columns represent median values of different cytokines. *P < 0.05; **P < 0.01
Discussion
The present study was addressed to assess the influence of SERPINB3 in experimental ALF induced by acetaminophen using a transgenic mouse model, as previous findings have indicated that SERPINB3 was over-expressed in LPS-activated microglial cells 15 and brain and systemic inflammation are major determinants of acute liver failure.
To date, the mechanisms underlying the initiation and control of immune responses in central nervous system inflammation are poorly understood. Microgliosis is a recognized sign of brain inflammation 31 and microglia components have been involved as antigen-presenting cells in the stimulation or reactivation of brain-targeted T-cells. 32 Experimental models of brain inflammation have documented that, in addition to inflammatory macrophage-like brain cells, activated myeloid dendritic cells accumulate at inflammatory sites within brain tissue and functionally mature as disease progresses. 33
The results of our study have shown that injured trans-genic mice demonstrated a shorter survival, associated with a more pronounced Th1 cytokine profile in serum compared with control mice. This immune condition was likely the main factor determining extremely low-body temperature, leading to premature animal death. Even if this mechanism is usually considered a regulatory response to systemic inflammation,34,35 we could assume that an over-expression/persistence of SERPINB3 might amplify this process, thus becoming dangerous with lethal implications. 36
At brain level transgenic animals showed increased glial cell number, compared with wild-type animals, and these features were detected both in mice with and without acetaminophen treatment. This finding was associated with significantly lower apoptotic death events, compared with controls, while proliferation activity was not affected, leading to a final positive cell balance. Immunohistochemistry revealed the presence of SERPINB3 in vascular endothelial cells of wild-type and transgenic mice, a finding described previously in normal tissues, 21 while this serpin was detected in microglial cells only in trans-genic animals. These results are in agreement with previous data, reporting the presence of SERPINB3 in inflammatory cells/macrophages of transgenic mice. 25 The biological role of this serpin is not completely known. It has been shown that SERPINB3 renders cells more resistant to apoptotic cell death, 37 inhibits migration of NK cells 38 and it is capable to induce epithelial–mesenchymal transition, with increased invasiveness cell potential. 14 Induction of TGF-β,39,40 inhibition of inflammation and increased epithelial proliferation in the lung 25 have been also reported. It is worth to note that SERPINB3 has been found over-expressed in activated peripheral blood mononuclear cells,22,41 although their immunophenotype has not been characterized yet. We can speculate that the increased levels of SERPINB3 in inflammatory cells and/or the increased GM-CSF levels could contribute to the activation of this cell compartment, leading to the enhanced Th1 inflammatory response observed in transgenic animals after acetaminophen injury.
In conclusion, the findings of the present study provided evidence that in acute liver failure the expression of SERPINB3 determines an enhanced inflammatory background, mainly mediated by higher levels of Th1 proinflammatory cytokines.
SERPINB3-transgenic mice might become a useful model for future studies to evaluate systemic reactions to infections and sepsis.
Footnotes
Acknowledgements
The authors thank Felipe V and Caolì O for their suggestions concerning acetaminophen inoculation, Butterworth RF for useful discussion and Plebani M for ALT testing.
National Ministry of Education, University and Research [FIRB Project Prot. RBLA03S4SP_005], Grant from the Associazione Italiana per la Ricerca sul Cancro (AIRC) [AIRC Project No 10235], Grant from the University of Padova [Project No CPDA110795].