
Abstract
In a previous study, we demonstrated the ability of papaya epicarp extract (PEE) to protect against oxidative stress-induced insult in human SH-SY5Y neuronal cells in a mechanism that appeared to be by means of PEE potent antioxidant properties. To further understand this relationship, we examined the effect of PEE intervention on aluminum (Al)-induced cytotoxicity in SH-SY5Y cells. The results indicated that PEE was effective in protecting against Al-induced cell death in a dose-dependent manner. PEE restored the Al-induced inhibition of folate-dependent methionine synthase activity and the antioxidant enzymes (catalase, glutathione peroxidases and superoxide dismutase). PEE ameliorated the Al-induced impairment of intracellular glutathione and total antioxidant capacity. Together, these findings indicate that PEE supplementation can play a neuroprotective role in ameliorating the changes in redox status of SH-SY5Y cells exposed to Al, a well-known environmental toxin that is involved in the pathogenesis of neurodegenerative diseases and neurodevelopmental disorders.
Introduction
Aluminum (Al) is a well-known environmental toxin that has been involved in the pathogenesis of neurodegenerative diseases. 1–3 The ingestion of foods, in particular the foods from animal origin, once they have been cooked or stored in Al containers, is the major source of toxicity exposure to this contaminant for the human population. 4–6 Several studies have shown that Al exposure can cause oxidative stress in both animal models (rat brain) and human-cultured neuronal cells in a mechanism that involves the generation of reactive oxygen species. 7–10 Al has also been shown to inhibit the activity of folate-dependent methionine synthase (MS) enzyme in a dose-dependent manner. 11 MS requires folate, as a methyl donor, and vitamin B12, as a co-factor, for mediating the methylation reaction of homocysteine (HCY) into methionine. 12 Inhibition of MS can result in accumulation of its precursor (HCY) and it was observed that in human aortic endothelial cells, HCY increased O2 •− concentrations (1.6-fold) in concomitant manner with a 20% reduction in cell viability and a 1.5-fold increase in apoptotic death. 13 Inhibition of MS caused a depletion in glutathione (GSH) in different human cultured cell lines. 14,15 The in vitro free radical scavenging property of papaya fruit against free radicals and environmental insults is well documented. 16,17 Furthermore, a recent study from our research group has shown that papaya epicarp extract (PEE) exhibited antioxidant activity and protected against hydrogen peroxide-induced oxidative stress and GSH depletion in human SH-SY5Y neuronal cells; however, the mechanism has not yet been elucidated. 18 The present study was conducted to evaluate the protective effects of PEE against Al-induced oxidative stress and impaired folate-dependent MS activity in SH-SY5Y cells. The outcomes of this study will help to understand the different neuroprotective attributes of PEE.
Materials and methods
Chemicals and supplies
All chemicals, α-minimum essential media (α-MEM) culture medium, penicillin–streptomycin–fungizone (PSF), trypsin-ethylenediaminetetraacetic acid (EDTA) solution, fetal bovine serum (FBS) and modified Kreb's solution were purchased from Sigma Chemical Co. (St Louis, MO, USA).
PEE preparation
Fresh green papaya was washed and the epicarp was separated by manual peeling using a sharp knife. The epicarp was dried in an air convection dryer at 40°C for 18 h and then placed in desiccators containing silica gel at 20°C for one week. The water content of the epicarp was 77.4 g/100 g skin. The dried epicarp was grinded into powder using a hammer mill with sieve size of 1.0 mm (Model MF 10 Basic; IKA Works, Wilmington, NC, USA). The dried powder was equilibrated for four weeks in a desiccator at 20°C containing a saturated lithium chloride solution in a beaker to maintain the relative humidity of the environment at 11.3%. The equilibrated dried sample reached a water activity of 0.113. The dried powder was mixed with distilled water (10 g/150 mL) and the mixture was stirred on a magnetic stirrer for 4 h at room temperature. The mixture was then centrifuged at 2000
Cell culture
Human SH-SY5Y neuronal cells were purchased from American Type Culture Collection (ATCC; Manassas, VA, USA) and were grown as a monolayer in 10 cm Petri dishes that contained 10 mL of α-MEM supplemented with 1% PSF and 10% FBS. Cultured cells were placed in an incubator chamber containing humidified 95% air and 5% CO2 at 37°C. For passage, confluent cells were detached with 1.0 mL trypsin-EDTA solution. They were then re-suspended in 10 mL of cell culture medium and incubated for 24 h before conducting the experiment.
The experimental design of this study was to incubate SH-SY5Y cells for 60 min with different doses (10–12–10–4 mol/L) of Al salt (AlCl3) in the presence or absence of 30 μg/mL of PEE. After treatments, the cells were washed twice with modified Kreb's solution, scraped, pelleted and re-suspended in 1 mL of 100 mmol/L phosphate buffer, pH 7.4. Cell membranes were disrupted by sonication on ice and the cell lysates were centrifuged at 4°C, 10,000
Cell viability using MTS assay
MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium], in the presence of phenazine methosulfate, is chemically reduced into formazan product which is soluble in tissue culture medium and its color absorbance can be measured at 490–500 nm. 19 A colorimetric method using MTS Assay Kit (Cell Titer 96® Aqueous Non-Radioactive Cell Proliferation Assay; Promega Corporation, Madison, WI, USA) was carried out using 96-well microplates and the production of formazan was read at 490 nm using a standard 96-well plate reader. The intensity of the color produced was proportional to the number of living SH-SY5Y cells.
Protein measurement
The protein content of cell lysates was measured by the method of Lowry et al. 20 Bovine serum albumin was used as the standard and the protein contents were expressed as mg/mL of sample.
MS activity measurement
After treatments, SH-SY5Y cells were scraped. Approximately 108 cells were suspended in 1 mL of 100 mmol/L phosphate buffer (pH 7.4). Cell membranes were disrupted by sonication on ice and the homogenate centrifuged at 4°C. Enzyme assays were performed under anaerobic conditions. The reaction mixture contained 100 mmol/L potassium phosphate (pH 7.2), 500 μmol/L HCY, 152 μmol/L S-adenosylmethionine, 2 mmol/L titanium citrate, 250 μmol/L (6R,S)-5-14CH3-THF (tetrahydrofuran) and the enzyme in a final volume of 500 μL. The reaction was initiated by addition of CH3-THF, incubated for 60 min at 37°C and terminated by heating at 98°C for two minutes. Radio-labeled methionine was separated on a Dowex 1-X8 column (Sigma-Aldrich Corporation, St Louis, MO, USA), which was eluted with 2 mL of water. Control assays, in which sample enzyme was omitted, served as blanks.
Oxidative stress biochemical assays
The following biochemical markers were measured in the cell lysates according to the manufacturers’ instructions: GSH concentration with a glutathione assay kit (K251; Biovision, Mountain View, CA, USA), total antioxidant capacity (TAC) using a Randox assay kit (Randox Laboratories, Crumlin, UK). All antioxidant enzymes assay kits were purchased from Sigma Chemical Co.: catalase (CAT) assay kit (Catalog number CAT 100), GSH peroxidases (GPX) assay kit (Catalog number CGP1), superoxide dismutase (SOD) assay kit (Catalog number 19160).
Statistical analysis
Statistical analysis was performed using GraphPad Prism (version 5.03; GraphPad Software Inc., San Diego, CA, USA). The results are expressed as means ± standard error of means. Comparisons among groups were performed with one-way analysis of variance, followed by Tukey's test. The Student's unpaired t-test was used for pair-wise comparisons. The values at the concentration of P < 0.05 were considered to be significantly different.
Results
In this study, we investigated the Al-induced morphological changes in SH-SY5Y cells and measured the folate-dependent MS activity, TAC, intracellular GSH and antioxidant enzymes of the SH-SY5Y cells.
Effect of Al and PEE on SH-SY5Y cell viability
Our previous study reported that the presence of PEE at a concentration of 30 μg/mL exerted a significant protective effect against oxidative stress-induced insult in human SH-SY5Y neuronal cells. 18 Therefore, 30 μg/mL PEE was applied to explore its antioxidant properties against the Al-induced neurotoxicity.
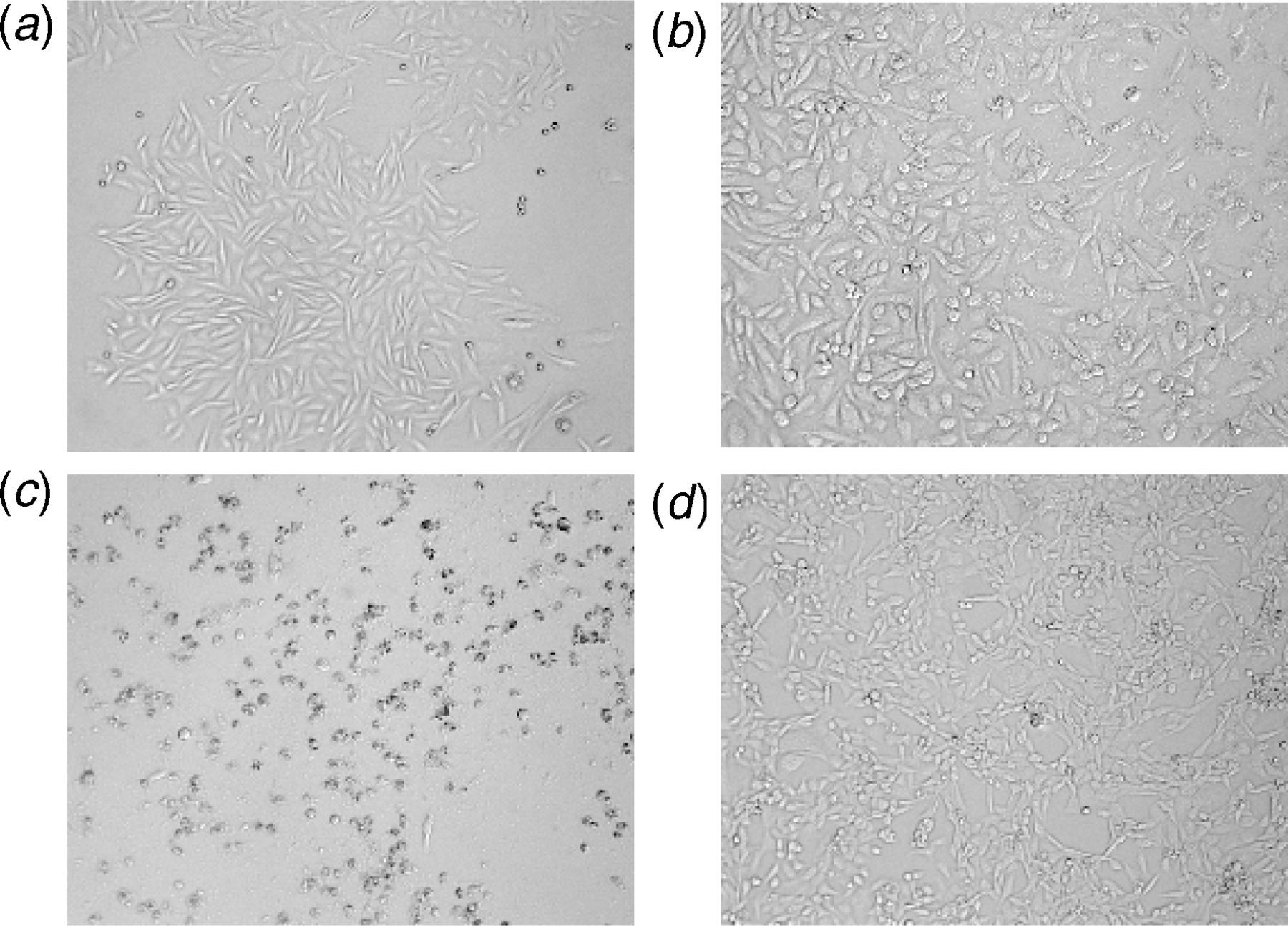
As presented in Figure 1, Al potently decreased the SH-SY5Y cell viability in a dose-dependent manner with a threshold of inhibition at 1 μmol/L (10−6 mol/L). Al at a concentration of 10 pmol/L (10−11 mol/L) did not (P > 0.05) affect the SH-SY5Y cell viability and only caused a change in the morphology of SH-SY5Y cells (Figure 2a) as compared with untreated cells (Figure 2a). However, the treatment of cells with Al at a concentration of 1 μmol/L (threshold limit of inhibition) caused the normally spindle-shape SH-SY5Y cells to condense and round-up as an indication of apoptosis (Figure 2c). Interestingly, the concomitant treatment of SH-SY5Y cells with 1 μmol/L Al and 30 μg/mL PEE (Figure 2d) protected cells and maintained their normal appearance and growth as compared with treated cells (Figure 2c). We concluded that Al at a concentration of 10 pmol/L was not cytotoxic and did not induce any cell death; therefore, it was used as a treatment dose for all the experiments included in this study, and hence the observed Al-induced oxidative stress is not attributed to SH-SY5Y cell death.
Dose-dependent effect of aluminum (Al) and papaya epicarp extract (PEE) on SH-SY5Y cell viability. Cells were treated for 60 min with different concentrations of Al in the presence or absence of 30 μg/mL PEE and then subjected to a MTS assay. Treatment with 10−10–10−4 mol/L Al significantly decreased cell viability (P < 0.05) as compared with 10−12–10−11 mol/L and control, untreated cells. Incubation of cells with 30 μg/mL PEE significantly reduced the Al-induced cell death in a dose-dependent manner. Cell viability is expressed as percent of control. Results are the means ± SEM of six measurements. MTS, 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium Changes in SH-SY5Y neuronal cell morphology. (a) Control, normal untreated cells. (b) Cells treated for 60 min with 10 pmol/L aluminum. (c) Cells treated with 1 μmol/L aluminum. (d) Cells concomitantly treated with 1 μmol/L aluminum and 30 μg/mL papaya epicarp extract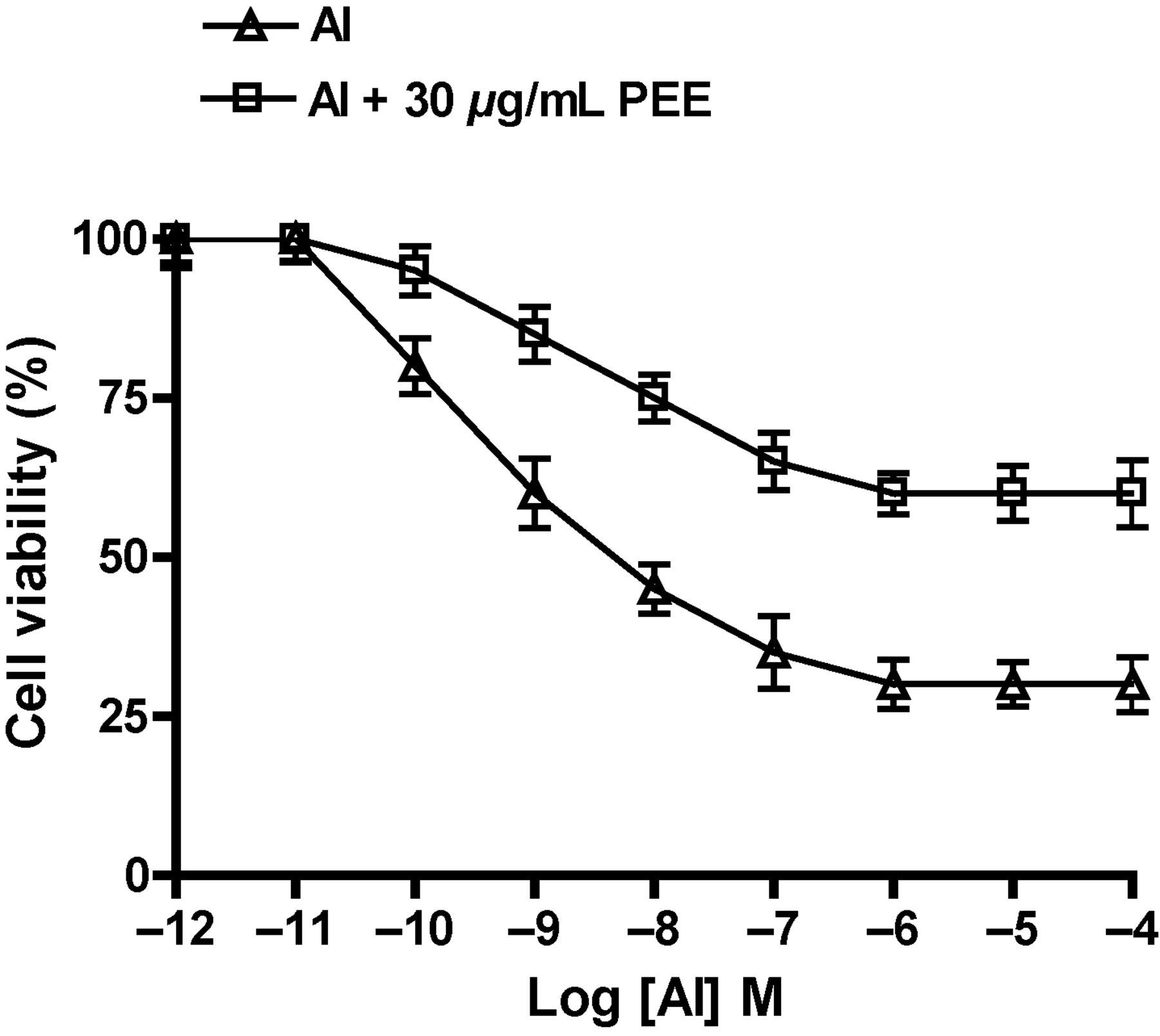
Effect of Al and PEE on redox status of SH-SY5Y cells
Protective effect of papaya epicarp extract against aluminum-induced impairment of oxidative stress indices in SH-SY5Y neuronal cells
Al, aluminum; PEE, papaya epicarp extract; GSH, glutathione; TAC, total antioxidant capacity; CAT, catalase; GPX, glutathione peroxidases; SOD, superoxide dismutase
GSH and TAC are expressed as nmol/mg protein. The unit of CAT, GPX and SOD enzymes activities is μmol/min/mg protein
SH-SY5Y neuronal cells were incubated with Al (10 pmol/L) in the presence or absence of PEE (30 μg/mL) for 60 min
Results are the means ± SEM of six measurements
PEE was compared with the control group, and there was no significant difference, P > 0.05
Al + PEE, PEE and Al groups were compared with the control group. Data with (a and b) superscripts are significantly lower than the control group, P < 0.05
Al + PEE, PEE groups were compared with the Al group. Data with hash (#) superscripts are significantly higher than the Al group, P < 0.05
Effect of Al and PEE on MS activity of SH-SY5Y cells
Protective effect of papaya epicarp extract against aluminum-induced inhibition of methionine synthase activity in SH-SY5Y neuronal cells
Al, aluminum; PEE, papaya epicarp extract
Methionine synthase (MS) activity is expressed as pmol/min/mg protein
ND, no detectable MS activity
SH-SY5Y neuronal cells were incubated with Al (10 pmol/L) in the presence or absence of PEE (30 μg/mL) for 60 min. Results are the means ± SEM of six measurements. Al + PEE and PEE groups were compared with the control group, and there was no significant difference, P > 0.05
Discussion
Our study is the first report that demonstrated the amelioration of Al-induced neurotoxicity by PEE using SH-SY5Y cells as an in vitro model. Considering that PEE exhibits an antioxidant action, 18 it is an additional advantage that it could be used as an adjunct therapeutic agent to protect against oxidative-stress associated diseases.
MS is a ubiquitous enzyme that regulates HCY-transmethylation into methionine and transsulfuration into GSH, the major intracellular antioxidant. Our study reported a reduction in the indices of oxidation stress markers (Table 1) as well as MS activity (Table 2) in association with Al-treatment. A reduction on MS activity was associated with a depletion of GSH, an index of cellular oxidative stress. This is consistent with previous reports that have reported that inhibition of MS activity is associated with a reduced cellular GSH concentration and proposed the hypothesis that the reduced MS activity is involved in the etiology of impaired cellular oxidation status. 11,14
Impairment of MS activity by Al toxicity suggests a plausible mechanism. However, the consequently elevated HCY concentration is itself an independent risk factor for oxidative stress and neuronal cell dysfunction. 21 A recent study indicated that pretreatment of SH-SY5Y cells with physiologically relevant concentrations of cyano-cobalamin (10–50 nmol/L) prevented the HCY-induced increases in O2 •− and cell death. 22 Folate supplementation, in another study, inhibited both the HCY and rotenone-induced mitochondrial O2 •− production. 23 The results of our study are in line with these findings and showed that PEE restored the inhibited MS activity in Al-treated SH-SY5Y cells. PEE also augmented GSH, TAC and antioxidant enzymes in SH-SY5Y cells after exposure to Al. Likewise, in our study, the Al-toxicity was evident by morphological changes and death of the treated cells. The human brain has approximately 100 billion neuronal cells that synchronize together for proper functions of neuromotor integration/regulation. Neuronal cell population death deregulates the synchronization process with a subsequent onset of various neurodegenerative disorders. Although this phenomenon is common during aging, it has also been reported that during early childhood development, certain environmental insults can induce oxidative stress in brain neurons and ultimately may lead to neuronal cell death/injury. 12
Our study is the first to report the reduction in Al-induced neuronal oxidative stress by PEE extract. The main outcomes of this study highlight the importance of PEE as an effective dietary intervention to minimize the Al-toxicity on human neuronal cells.
Conclusion
PEE exhibited an efficient antioxidant activity that could protect against the neurocytotoxicity of Al and, subsequently, neuronal cell damage. Further studies are, however, required to investigate the association of PEE consumption with the reduction of Al toxicity-related diseases among vulnerable population groups.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by HM Strategic grant (SR/AGR/FOOD/11/01), Internal Research grants (IG/AGR/FOOD/10/01 and IG/AGR/FOOD/11/01) from the College of Agricultural and Marine Sciences, Sultan Qaboos University (SQU), and by the International Islamic University Malaysia Grant, IIUM/504/RES/G/14/3/RMGS/08-04. The authors are deeply thankful to Dr Allal Ouhtit, Genetics Department at SQU, for providing chemicals and supplies, and the laboratory facility for conducting the experiments of this study. Special thanks to Amani Al-Rawahi for her technical help in PEE preparation.