
Abstract
Five decades after the first documented use of a laser for wound healing, research in light therapy has yet to elucidate the underlying biochemical pathways causing its effects. The aim of this review is to summarize the current research into the biochemical mechanisms of light therapy in order to better direct future studies. The implication of cytochrome c oxidase as the photoacceptor modulating light therapy is reviewed, as are the predominant hypotheses of the biochemical pathways involved in the stimulation of wound healing, cellular proliferation, production of transcription factors and other reported stimulatory effects.
Keywords
Introduction
Numerous studies over the past several decades have shown beneficial effects of light therapy. These effects include stimulation of neuronal growth,1–3 retinoprotective effects, 1 cardi-omyocyte protective effects postischemia, 4 stimulation of DNA and RNA synthesis, promotion of cell adhesion, 5 improved neurological function post-traumatic brain injury (TBI) and stroke,6–10 acceleration of wound healing,1,11 cellular and extracellular matrix proliferation 12 including fibro-blasts, 13 collagen production13,14 and granulation tissue formation,13,14 and reduction of the inflammatory response.13,15,16 While many of these phenomena and effects have been researched, no study has ever been able to prove the underlying stimulatory etiology responsible for the beneficial effects. Light therapy for photobiomodulation remains an alternative medicine for this reason.
Studies in light treatment over the past 30 years have primarily utilized three model systems: formic acid poisoning of the optic nerve, neuronal bridging and proliferation, and most predominantly, wound healing. Formic acid, the toxic metabolite of methanol, is known to be inhibitory in the mitochondrial respiratory chain,1,17 effectively starving optic cells to death. A study utilizing 670 nm visible red light on methanol-intoxicated rats, has shown 72% electro-retinogram (ERG) functionality of control in the light-treated group with only 28% functionality in the untreated for rods and M-cones. 1 Similarly, ultraviolet cones showed 61% and 30% ERG functionality of control for light therapy and untreated groups, respectively. Attenuation of formic acid's toxic optic effects by light therapy is suggested to manifest in increased cytochrome c oxidase functionality to counteract the competitive binding of formic acid.1,17,18
Neuronal stimulation by light therapy has been demonstrated in a myriad of studies in both the peripheral (PNS), as well as the central nervous system (CNS). Contusion and hemi-section damage models have been used to study light therapy's proliferative effect on the PNS, with results including increased neuronal count,3,19 decreased de-nervated muscle atrophy, 20 and increased bridging axonal thickness. 2 CNS models have included spinal cord damage,21,22 TBI and stroke, demonstrating functional and behavioral improvement with the addition of light therapy.6–10 Irradiation wavelengths for neuronal studies are summarized in Table 1. Investigations into neur-onal recovery with light therapy have provided little to no information on the mechanism of action; however, they show promise for future treatments.
Irradiation parameters studied in nervous system injuries

PNS, peripheral nervous system; CNS, central nervous system
The majority of all light therapy studies have focused on wound healing, providing a wealth of information on its proliferative effects on the cellular reconstruction process. Each of the wound healing stages has been shown to benefit from light's stimulatory effects as seen in Figure 1.
Wound healing stages light therapy is cited to act upon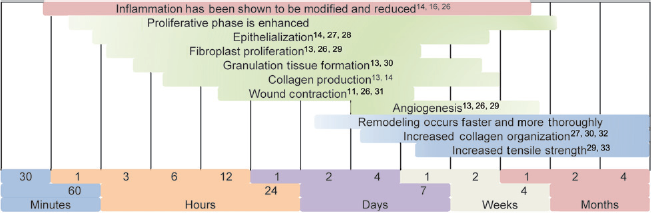
While these wound healing studies contribute little to the fundamental understanding of the mechanism of light therapy, they contribute two important concepts. Many studies attribute light's stimulatory effects to its initial ability to modify the inflammatory response.34,35 More importantly, studies conducting varying control models show the possibility of a systemic response, not simply a local effect16,26 mediating proliferative effects at wound sites distant from that of irradiation.
Studies that have focused specifically on the mechanism of light therapy have shown increases in cellular metabolites and signaling molecules including ATP, reactive oxygen species (ROS) and nitric oxide (NO), leading to the currently accepted belief that the mitochondrial respiratory chain enzyme cytochrome c oxidase is the chromophore, receiving photostimulation. Action spectra reported by several laboratories support this photoaccepting ability by the interchangeable redox states of the chromophore, indicating that light provides the energy necessary to switch from one conformation to the other.
Examinations of the mechanisms of ATP production have led to the discovery of a correlation with ROS production and the currently predicted model of elevated chemiosmotic gradient as the primary effect of light stimulation. The production of NO is examined as a secondary signaling molecule responsible for vascular dilation and growth, as well as antibacterial effects and cytokine and growth factor regulation.
Cytochrome c oxidase as a photoacceptor
The theory that light therapy is mediated by cytochrome c oxidase stems from the basic knowledge of chromophores. In order for light energy to be absorbed by a biological system, it must contain one of the few natural light modulating structures. Because cytochrome c oxidase contains two copper centers (CuA and CuB) and two iron centers (Heme a and Heme a3) responsible for electron transfer to molecular oxygen, 36 it is understandable that early experiments looked to the d-orbital valence electrons of these transition metals as possible mediators of light therapy.
A majority of the evidence for cytochrome c oxidase as the photoacceptor over the years has come from experiments on ‘functionally inactivated’ (cytochrome c oxidase inhibited) respiratory chains and the partial resuscitating ability of light. Cytochrome c oxidase inhibitors azide, 37 potassium cyanide, 38 formate,1,17,18 paraquat, rotenone and antimycin A 39 have been used in combination with light therapy to show increases in levels of HeLa cell adhesion from inhibited to 150% of control, 37 increases in ATP levels in neurons from inhibited to 95% of control 38 and protective effects from methanol toxicity in multiple retinal cases.1,17,18
Action and absorption spectra are becoming more widely used as evidence for a cytochrome c oxidase phototherapy mechanism due to the work of Karu et al., and it is important to understand the fundamental differences between the two. An absorption spectra indicates the wavelengths of light attenuated or removed from the spectrum by any particular molecule or complex, while the action spectra indicates the intensity of a biological response as mediated by a wavelength along the spectra; there is no typical action spectra criteria, and the results will always be representative of a secondary response to the stimulation.
In the ideal scenario of a primary response to light stimulation, the action spectra would mimic the absorption spectra in both intensity and wavelength. As a primary response is as of yet unobtainable from the advanced multi-meric protein complex of cytochrome c oxidase in an in vivo situation, secondary observations of DNA and RNA synthesis and cell adhesion to glass matrices have been used in quantifying light therapy, and it has been shown with multiple experiments a wavelength-dependent photostimulatory effect on the proliferation of each of these factors.5,40,41 Figure 2 illustrates the possible excitatory pathway by which light stimulates the electron transport chain to higher functionality.
Effects of light therapy on the mitochondrial electron transport chain. Cytochrome c is stimulated to switch redox states, transferring electrons to cytochrome c oxidase at a higher rate, increasing proton transport and subsequently the mitochondrial membrane potential. Increase in hydrogen ion gradient drives ATP synthase to increase ATP catalysis. Electron transport follows decreased energy states  , while electron leakage reduces oxygen to reactive oxygen species
, while electron leakage reduces oxygen to reactive oxygen species
Early studies on the adhesion of HeLa cells in different states of oxidation led to absorbance curves indicating wavelength trends to redox states of the respiratory chain. It was found that upon irradiation, cells in moderately oxidized or reduced states would switch to the opposite, as indicated by the absorption bands of that region. Wavelengths in the 710–790 nm range were found to be characteristic of a reduced photoacceptor while those in the 560–680 nm region were characteristic of one that is oxidized, leading to the 760/665 nm redox span. 40
The action spectra studies of Karu et al. can be summarized into the resolution of four distinct peaks in the red-infrared spectrum. By cross-referencing with spectra from cytochrome c oxidase structure-determining studies, each peak may be resolved to a redox state of the copper atoms CuA and CuB. The resolved wavelengths are shown in Table 2.
Resolved active wavelengths of cytochrome c oxidase chromophores 42

Typical ROS production
Although an efficient form of energy transduction, oxidative phosphorylation releases a small percentage of electrons from the matrix membrane-bound protein complexes during chemiosmosis. Oxygen, a strong oxidizing agent, readily accepts these electrons, producing superoxide (O2-1) and peroxide (O2-2), respectively, upon reduction. 43 These charged free radicals in turn reduce proteins and DNA to form mutation and nucleotide modifications, 44 hindering protein functionality and replication, and in some cases leading to apoptosis of the cell. 45 In order to combat these destructive radicals, cells are equipped with superoxide dismutase (SOD), catalase and other dismutative enzymes capable of converting superoxide and peroxide down to benign water and oxygen molecules. 44
ROS signaling
It is well known that at significant concentrations, ROS initiate the apoptotic cascade though the degradation of the mitochondrial membrane potential, cytochrome c release, caspase activation and DNA digestion. 46 At concentrations closer to that of physiological, ROS have been shown to have stimulatory and signaling abilities. Aside from SOD up-regulation, ROS have been shown to be involved in stem cell differentiation, 47 ion channel interference, including increases in calcium flow, 48 cellular growth, 49 mitogen-activated protein kinase (MAPK) activation and Janus kinase transcription activation. 50
ROS in light therapy
ROS have gained much attention in light therapy studies as they have been observed to increase in concentration as a result of light treatment.39,49,51 Interestingly, ATP has been shown to have the same correlation.12,25,38,39,52,53 The major difficulty in quantification of ROS and the reason why few studies have investigated this effect is the shortlived nature of the oxygen radical. Rarely in nature do charged particles exist, and when they do, their redox potential causes them to participate in neutralizing reactions nearly immediately after they are formed.
Several methods have evolved, embracing or circumventing the free radical nature for ROS studies. Spin trapping, a technique utilizing radical binding agents TEMPO or DEPMPO capable of detection with electron paramagnetic resonance spectroscopy indicates not only the presence of radicals, but also the type.49,51 Dosage information may be obtained through the addition of external radicals to culture; 51 and lastly, circumventing radical metrics, secondary transcription factors up-regulated by ROS can also be quantified more readily. 39
Investigations into the ROS signaling pathway have implicated calcium ion flow as a secondary messenger of light therapy, and as such, this flux has been quantified for dose-dependent mechanisms. Addition of external free radical peroxide (H202) in place of light therapy has been shown to produce identical levels of calcium stimulation, matching doses of light irradiation. 51 The ability to illicit an identical response indicates ROS production (though not the source) to be the primary effector in a calcium ion pathway.
Studies on the production of ROS and ATP with and without electron transport chain (ETC) inhibitors have provided some insight into the concentrations of each produced upon irradiation, and not surprisingly, results indicate a significantly elevated production of ROS upon late ETC inhibition with antimycin A. However, and more importantly, ATP and ROS production without inhibition are shown to increase nearly 30% and 300%, respectively. 39 While typical ROS production is small, less than 1% of electron transfer, a three-fold increase can have significant effects in the number of reduced proteins as a result of light therapy.
Mitochondrial membrane potential
Over the last several decades, numerous studies in light therapy have indicated increased production of ATP from mitochondria.12,25,38,39,52,53 A more recent analysis has shown by flow cytometry that there is a simultaneous increase in mitochondrial membrane potential (ΔΦmt) with the addition of light treatment. 12 The significance of changes to this gradient is not only in the functionality of ATP synthase and the production of ATP, but the typical flow of electrons in the ETC membrane proteins. The increase in chemiosmotic gradient responsible for accelerated ATP synthase activity appears to be balanced by an increased production of ROS from straying electrons. Under these conditions, it would appear that the mechanism of light therapy is based in the ability to accelerate the functionality of the electron transport chain. However, with the higher ROS/ATP production ratio, it may be apparent that the mitochondrion is incapable of effectively utilizing a higher chemiosmotic gradient.
NO production
Production of NO is mediated by three constitutive nitric oxide synthase (NOS) isoenzymes: neuronal (nNOS), endothelial (eNOS) and inducible (iNOS). While all NOS isoforms produce NO from arginine, they differ in their activation pathways: nNOS and eNOS lie cytosol and membrane-bound, respectively, and are both calcium (Ca2+)- and calmodulin-dependent. iNOS expression is brought about by exposure to a wide array of cytokine and bacterial products, and operates independent of calcium or calmodulin. 54
Cytochrome c oxidase-dependent nitrite oxidation to form NO
A second method of NO production has been elucidated more recently from studies on cytochrome c oxidase under anoxic conditions. In the presence of oxygen, the typical reduction pathway of cytochrome c oxidase follows:

In anoxic conditions, cytochrome c oxidase has been shown to function with catalyzing ability for nitrite NO2-, oxidizing it to NO55,56

Light stimulation of cytochrome c oxidase oxidation
Isolation of this reaction in anoxic/NOS inhibitory conditions on mitochondria has demonstrated light therapy in the green to red band of visible light to be significantly effective in catalyzing the ability of cytochrome c oxidase to produce NO 57 A similar study on the hypoxic/reoxygenation of cardiomyocytes found that upon the addition of a universal NOS inhibitor, the protective effect of near infrared (NIR) light was attenuated, but not as dramatically as when NO scavengers were added. 4 While light has been effectively demonstrated to have an influence on the production of NO through a cytochrome c oxidase oxidation alternative mechanism, its dependence on in vivo nitrite has yet to be seen. Simultaneously, this mechanism cannot be ruled out, as NIR therapy has proven to remain effective in the absence of NOS enzymes, also potentially supportive of a secondary pathway.
NO dissociation from cytochrome c oxidase inhibition
Not unlike cyanide (CN) and carbon monoxide, NO has been shown to bind to the heme a3-CuB active site of cytochrome c oxidase in an oxygen-displacing, competitive mechanism.58,59 Unlike the toxic effect of CN and CO, however, NO attaches and dissociates from cytochrome c oxidase in a regulatory mechanism controlled by its concentration in the mitochondrion, resulting from NO producers including NOS. 59 The a3-NO complex has been shown to be bound by a photolabile bond, capable of being photomodu-lated as seen by absorption spectra in the 597 nm region. 60 It has been suggested that light therapy photodissociates NO from the heme a3-CuB active site, decreasing the number of functionally inactivated electron transport chains and increasing metabolic turnover.57,59,61,62
NO signaling
NO, also known as endothelial derived relaxing factor, mediates vasodilation by relaxing vascular smooth muscle and increasing diameter. More recently, however, it has been implicated in a number of mechanisms mediating wound healing. NO modulates the inflammatory and immune response by inhibiting T- and B-cell proliferation, antibody production by CD5 B-cells, T- and B-cell diversification and leukocyte recruitment. 63 Immunoresponsive cells including phagocytes, glial cells, endothelial cells and fibro-blasts, up-regulate eNOS, iNOS or both for the production and secretion of NO, utilizing NO's radical oxidative ability to damage bacteria, viruses and other infectious agents.63,64 Studies have shown that light therapy modulates this expression of iNOS, 65 providing a potential explanation for the modified inflammatory response seen in light therapy wound healing studies.
NO is also responsible for the production and regulation of several cytokines, chemokines and growth factors regulating the pro- and anti-inflammatory response, as seen in Table 3.
Nitric oxide modulation of cytokines and growth factors 63

IL, interleukin; INF-γ, interferon-γ; TNF, tumor necrosis factor; TGF-β, transforming growth factor-β; MCP-1, monocyte chemoattractant protein-1; VEGF, vascular endothelial growth factor; MIP-1α, macrophage inflammatory protein-1α; G-CSF, granulocyte colony-stimulating factor; M-CSF, macrophage colony-stimulating factor
Angiogenesis, one of the critical processes in reperfusion and tissue growth, has been shown to be modulated by NO in an eNOS → vascular endothelial growth factor (VEGF) up-regulation pathway.66,67 NOS inhibition prevents both VEGF induction of endothelial cell growth and migration as well as induction of MAPK activation. 66 NOS enhancement by over-expression and inhibitory studies have shown NOS to be an important mediator of collagen and granulation tissue formation in the late stages of wound remodeling. Transforming growth factor (TGF)-β1 has been implicated in this mechanism as well as the angiogenic pathway. 66
Overproduction of NO is also known to cause damaging effects to the cell. The reaction of NO with superoxide produces an even more damaging radical peroxynitrite:
68
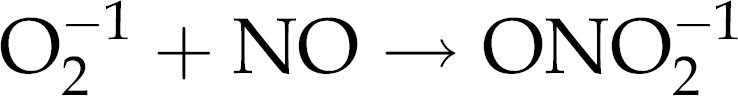
The damaging oxidative effect of peroxynitrite is attributed to its ability to interrupt iron metabolism, leading to mito-chondrial dysfunction and resulting in apoptosis. 68
Calcium modulation
Perhaps one of the newest suggestions to the light therapy mechanical pathway is the contribution of intracellular calcium as a signaling molecule. Calcium release has been shown in previous studies as having a direct correlation to the concentration of ROS, in particular hydrogen peroxide (H2O2). 51 A study examining this relationship found that the same increase in Ca+2 can be brought about by the addition of H2O2 or the application of light therapy. The response to each acted in a dose-dependent manner, with damaging effects resulting from high doses of either. 51
The relationship of ROS to calcium influx is thought to be brought about by either the phosphorylation of calcium transporters or ROS oxidation of thiols to disulfide bridges within their structure. This functional enhancement is shown to decrease upon the addition of verapamil, demonstrating that L-type calcium channels are at least partially responsible for the transport.48,51
A second suggested pathway involves the incorporation of phosphorous into the cellular membrane after light irradiation, and subsequent phosphatidylinositol 4,5-bispho-sphate (PIP2) production. PIP2 would then follow the typical signaling pathwayof phospholipase C catalysisto inositol tri-phosphate (IP3) and diacylglycerol (DAG) and subsequent Ca±2 release signaling to the endoplasmic reticulum. 69 A third suggested pathway includes the opening of the transient receptor potential vanilloid Ca+2 membrane channel upon cytosolic alkalization, resulting from increased mito-chondrial membrane permeability and ion influx. 70
Protein expression
As of yet most studies are not reporting protein expression as a product of light therapy; however, what is currently available can be seen in Table 4. Many of the studies reported in this paper have based their pathway hypothesis on these results and so they have been presented tabulated according to function.
Reported protein expression

Discussion
Two important topics in light therapy that do not directly pertain to the mechanism of action are the skin's optical window and dosimetric optimization, both of which for completion, are necessary to address.
The optical window of skin can be defined as the ideal range of absorption, or inversely as the spectrum allowed to transmit through epithelial tissue. The penetration depth of light into skin is a function of the wavelength, with longer wavelengths penetrating deeper into tissue than short wavelengths. 72 Although studies have included multiple wavelengths in their experimental design, either in a diode array, or separate test groups of a single wavelength, single wavelength trials utilizing light in the red spectrum (630–690 nm) have been shown to elicit the most productive biological response;21,73 red light has therefore been adopted by the majority of all light therapy studies.
Epithelial and cutaneous tissue light filtration is an important factor to consider in in vivo studies. One pilot study measured the light penetration transcutaneously to the spinal cord of a rat irradiated with 150 mW of NIR light, but measured only 9 mW through the skin, a result of only 6% transmission. 21 Most studies have irradiated with fluencies significantly less than 150 mW, averaging between 10 and 40 mW, with the bi-phasic dose response reaching its upper therapeutic threshold at the latter. 13 These results raise the question of whether light can be applied transcutaneously to reach intended cells at the same fluency used in in vitro studies without eliciting the damaging maximum of the bi-phasic response in transversed tissue. However, with the more recent use of subcutaneously applied light-emitting probes, this matter may become irrelevant.
Conversely, both the wound healing model, as well as the in vitro culture dish model, are free of the optical window constraint; excluding them from epithelial wavelength filtering and fundamentally separating them from transcutaneous applications, allowing the pure science of light therapy to be analyzed with less convolution.
The second important topic of discussion is the dosimetric optimization of studies. As previously mentioned, the bi-phasic dose response of light therapy is a proposed range of energy similar to a drug response curve, with a therapeutic range and a toxic threshold. The difference with the light therapy curve is that there are two parameters to the dose of light and thus the curve must be represented in three dimensions instead of two. Laser energy is quantified in Joules per area since the beam of light has a defined diameter. Application of this energy over time yields the unit of Watts, or Joules per second. Studies report fluencies in both Joules as well as Watts. The reason for the three-dimensional dose response curve is the balance of energy output and irradiation time to produce the total power out. For example, studies utilizing a high-energy laser may irradiate for milliseconds to seconds, while a low-energy laser may be applied for several minutes, with both studies applying the same power (Watts).
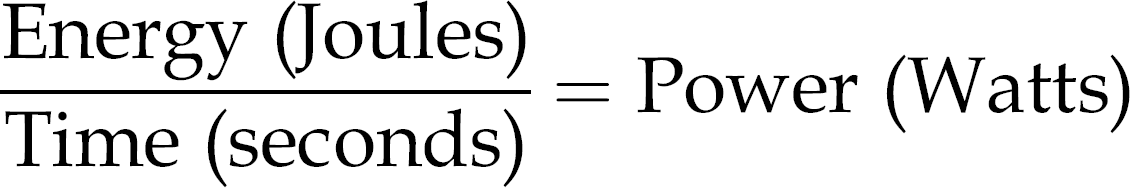
For this reason, it can be seen that unifying irradiation parameters is quite difficult stemming not only from variation in equipment power, but also the variability in laser spot size and application.
Although standardization of irradiation protocols has not yet been established, optimization of ideal wavelengths, as well as the development of the bi-phasic dose-response curve, have been great improvements to the field of light therapy over the past two decades. Large strides have been made in elucidating several steps of the light therapy mechanism as evidenced in this paper, and new advances are made each year. Though currently its use and application may not be fully optimized, light therapy is proving to be a low-risk, high-yield treatment for the improvement of wound healing and other currently untreatable defects like diabetic ulcers, toxin-related retinopathies and nerve damage.