
Abstract
Erythropoietin (EPO) has been well recognized as a tissue protective agent by inhibiting apoptosis and inflammation. The tissue protective effect of EPO, however, only occurs at a high dosage, which may elicit severe side-effects at the meantime. Helix B surface peptide (HBSP), a novel peptide derived from the non-erythropoietic helix B of EPO, plays a specific role in tissue protection. We investigated effects of HBSP and the expression of its heterodimeric receptor, beta common receptor (βcR)/EPO receptor (), in a murine renal ischemia reperfusion (IR) injury model. HBSP significantly ameliorated renal dysfunction and tissue damage, decreased apoptotic cells in the kidney and reduced activation of caspase-9 and 3. The βcR/EPOR in the kidney was up-regulated by IR, but down-regulated by HBSP. Further investigation revealed that the expression and phosphorylation of Akt was dramatically enhanced by HBSP, but strongly reversed by wortmannin, the PI3K inhibitor. Wortmannin intervention improved βcR/EPOR expression, promoted caspase-9 and 3 activation, and increased active caspase-3 positive cells, while renal function and structure, and apoptotic cell counts scarcely changed. This result indicates a significant contribution of PI3K/Akt signaling pathway in the renoprotection of HBSP. The therapeutic effects of HBSP in this study suggest that HBSP could be a better candidate for renal protection.
Keywords
Introduction
Erythropoietin (EPO) has been fully proved as an endogenous protein to exert tissue protective effects for a wide range of organs.1,2 However, a very high dose of EPO is required for peripheral administration to achieve tissue protection, 3 which consequently increases the incidence of hypertension and hypercoagulation.4,5 These undesirable risks of EPO are attributed to the homogeneous dimeric EPO receptor (EPOR)2 that mainly plays an erythropoietic role.
A heterodimeric receptor has been lately proved to mediate tissue protection in a more specific manner. It is composed of EPOR monomer and beta common receptor (βcR, CD131), the latter also shared by granulocyte– macrophage colony-stimulating factor, interleukin (IL)-3 and IL-5 in signaling pathways.6,7 Helix B surface peptide (HBSP) is a linear peptide derived from the amino acid sequence located on the aqueous surface of the helix B of EPO 8 and has been proved to have no interaction with the erythropoietic receptor. 8 Therefore, HBSP may play a tissue protective role without erythropoietic side-effects.
Tubular epithelial apoptosis is one of the main changes in the renal ischemia reperfusion (IR) injury, during which caspase family members such as caspase-9 and caspase-3 are activated and involved in executing cell death. On the other hand, antiapoptosis signaling pathways also exist as a feedback to restrain the damage and balance the tissue homeostasis. The PI3K/Akt signaling pathway is one of the most important pathways for cell survival. An interesting connection between the protective receptor subunit, βcR, and PI3K has been recently revealed. 9 Therefore, we propose that HBSP plays a protective role in renal IR injury through the heterodimeric receptor βcR/EPOR and PI3K/Akt pathway.
In the present study, protective effects of HBSP were investigated through renal function and structure, and apoptosis in the kidney. The βcR/EPOR expression in the kidney was detected. Furthermore, the impact of HBSP on the PI3K/Akt pathway was also evaluated by PI3K inhibitor, wortmannin.
Material and methods
Materials
HBSP was synthesized by Scipeptide Biotechnology, Ltd, Shanghai, China according to a previously published sequence containing 11 amino acids: QEQLERALNSS, produced by Brines M et al.; 8 and validated by its molecular weight using HPLC. All other materials used in this study were specified at appropriate places.
Renal IR injury
Male BALB/c mice (weighing 20-25 g) were obtained from Shanghai Slac Lab Animal, Co., Ltd, and bred in an experimental animal room of SPF grade. All animal procedures were performed according to the guidelines of the Care and Use of the Laboratory Animal Ethical Commission of Fudan University. Mice were randomly divided into four groups (n = 6): (1) sham group; (2) vehicle group: IR injury with phosphate-buffered saline vehicle intraperitone-ally injected; (3) HBSP group: IR injury with HBSP of 8 nmol/kg intraperitoneally injected at one minute, six hours, 12 h after the operation; (4) HBSP + wortmannin group: HBSP administered as in the HBSP group, and treated with one dose of wortmannin (Beyotime, China), 1 mg/kg, intraperitoneally, 30 min before operation. 10 For the renal IR model, the mice were anesthetized with pento-barbital at a dosage of 0.1 g/kg body weight intraperitone-ally. Core body temperature was maintained at 37°C using a homeothermic pad during surgery. The abdominal cavity was exposed via a midline incision, both kidneys were exposed and the renal pedicles were carefully isolated. Bilateral renal occlusion for 30 min was performed using non-traumatic vascular clamps. Occlusion was confirmed by observing blanching of the entire kidney surface. After removing the renal clips, the kidneys were observed for a further five minutes to ensure color change indicating blood reperfusion. Afterwards, 1 mL saline at 37°C was injected into the abdomen and the incision was sutured in two layers.
Animals were ethically sacrificed at 48 h after IR injury and the whole blood drawn from the heart was centrifuged at 4°C, 2500 rpm, for 25 min to obtain the serum sample. The level of serum creatinine (Scr) and blood urea nitrogen (BUN) was measured by the automatic biochemistry analyzer (Hitachi 7060, Hitachi Ltd., Tokyo, Japan). Bilateral kidneys were harvested and transversally cut at the mid-plane. One half of each side was fixed with 10% buffered formalin for histological assessment and immunohisto-chemistry, while the rest was frozen for Western blot and immunoprecipitation analysis.
Histological assessment
Hematoxylin and eosin (H&E) staining was performed to assess the histological injury. The tissue sections were blind-labeled and reviewed by a renal pathologist. A histological score system 11 was used to estimate the renal damage, which was graded by the percentage of tubule injury: 0 (<1%); 1 (1-10%); 2 (11-20%); 3 (21-40%); 4 (41-60%); 5 (61-75%); 6 (>75%). The scores represented the severity of tubular injury (including loss of proximal tubule brush border, cell swelling or vacuolization and cell necrosis): the score ranges of 1-2 represented mild injury, 3-4 represented moderate injury and 5-6 represented severe injury.
In situ end-labeling apoptotic cells
Five micrometer paraffin sections were used to label fragmented DNAs in situ with digoxigenin-deoxyuridine (dUTP) by terminal deoxynucleotidyl transferase (TdT) using a TUNEL Apoptosis Detection Kit (Millipore, Billerica, MA, USA). Briefly, sections were digested by 40 μg/mL proteinase K (EMD Chemicals, Gibbstown, NJ, USA) for 15 min at 37°C, incubated with TdT and digoxi-genin-dUTP at 37°C for 60 min and transferred to wash/ stop buffer for 30 min. After adding anti-digoxigenin-peroxidase complex for 30 min, these sections were developed by 3'-amino-9-ethylcarbazole (AEC, DAKO, Carpinteria, CA, USA, dark red color) substrate. Apoptotic cells were examined at × 400 magnification over 20 fields for semi-quantitation.
Immunohistochemistry of beta-common receptor
For immunohistochemistry staining, 5 μm paraffin sections were incubated overnight at 4°C with primary antibodies against beta-common receptor (1:100, Santa Cruz Biotechnology, Santa Cruz, CA, USA). The antibody binding was revealed by AEC. All of the sections were blind-labeled and the percentage of positive cells was determined in 20 fields of each section on a digital microscopic system (Olympus BX51/DP72, Tokyo, Japan).
Immunoprecipitation assay for EPOR combined beta-common receptor
The kidneys of mice were harvested and homogenized in a homogenizing buffer (containing 125 mmol/L NaCl, 1% Nonidet P-40, 1 mmol/L phenylmethanesulfonyl fluoride, and 5 μg/mL aprotinin, and 5 μg/mL leupeptin). For EPOR immunoprecipitation, tissue homogenates were incubated with 2 μg of anti-EPOR antibody (Santa Cruz Biotechnology) for two hours at 4°C on a rotator. Then, 30 μL protein A/G beads (Santa Cruz Biotechnology) containing the secondary antibody to anti-EPOR antibody was added to each sample and incubated overnight at 4°C on a rotator. The samples were spun briefly in a microcen-trifuge and washed three times in the homogenizing buffer. EPOR monomer and βcR/EPOR heterodimer, therefore, were purified from the protein homogenate and also excluded βcR monomer. Twenty microliters of loading buffer was added to each sample and boiled for five minutes. During this process of denaturation, the heterodimer of βcR/EPOR was dissociated. The samples were then separated by sodium dodecyl sulfate (SDS)-polyacrylamide gels, transferred onto nitrocellulose membranes, and probed with anti-βcR antibody (anti IL-3/IL-5/GM-CSFRβ antibody, Santa Cruz Biotechnology). The level of βcR in this last step of Western blot detection represented the level of βcR/EPOR heterodimer.
Immunostaining of cleaved Caspase-3
Immunohistochemistry staining of cleaved caspase-3, recognizing 17 kDa activated subunit, was undertaken on paraffin sections using a DAKO ChemMate EnVision Detection Kit (DAKO). Antigen was blocked by peroxidase blocking reagent. The sections were labeled by an anti-cleaved caspase-3 antibody (1:2000, Cell Signaling Technology, Boston, MA, USA) at 4°C overnight or normal rabbit immunoglobulin G (DAKO) at the same protein concentration of primary antibody as negative control. The antibody binding was revealed by AEC. Active caspase-3 positive cells in the cortex of kidneys were semi-quantitatively scored in 20 fields at × 400 magnification.
Western blot analysis
Renal tissue homogenates were prepared and then the supernatant was obtained at 4°C. Thirty micrograms of protein from each sample were separated by SDS– polyacrylamide gels and transferred onto polyvinylidene fluoride membranes. The primary antibodies were added and incubated at 4°C overnight including anti-cleaved caspase-3 (Asp175), anti-PI3K p85, anti-Akt or anti-phospho-Akt (Ser473, Thr308) (Cell Signaling Technology), followed by one hour incubation with peroxidase-conjugated secondary antibodies (Jackson Immuno-Research, Malvern, PA, USA) at room temperature. Immunoreactive bands were visualized using ECL Western Blotting Substrate (Thermo Fisher Scientific, Rock-ford, IL, USA). For the loading control, the same membranes were probed with anti- glyceraldehyde-3-phosphate dehy-drogenase (Cell Signaling Technology) or anti-β actin (Abcam, Cambridge, MA, USA) at the same time. The signals were quantified by scanning densitometry using a Bio-Image Analysis System (Bio-Rad, Hercules, CA, USA). The results from each experimental group were expressed as relative integrated intensity compared with that of control measured with the same batch. The appropriate negative control experiments were run for each target protein, as well as beta actin or GAPDH, by replacing primary antibodies with the normal immunoglobin of same species at the same protein concentration when the experimental condition was optimized.
Statistical analyses
Data are presented as mean ± standard error of the mean (SEM). Statistical analysis of the data was performed with the two-tailed independent t-test between two groups, and one-way analysis of variance (ANOVA) among more than three groups, using SPSS 18.0 software (SPSS Inc, Armonk, NY, USA). P < 0.05 was considered as statistically significant.
Results
Renal function and histological changes
Scr and BUN were markedly increased by IR injury compared with that in the sham group (119.80 ± 6.41 versus 14.69 ± 1.72μmol/L, sham, P<0.01; 50.33 ± 5.09 versus 14.87 ± 1.67 mmol/L, sham, P < 0.01). HBSP treatment significantly reduced the level of Scr and BUN (23.83 ± 1.32 versus 119.80 + 6.41 μmol/L, vehicle, P < 0.01; 10.62 ± 0.79 versus 50.33 ± 5.09 mmol/L, vehicle, P < 0.01). However, wortmannin treatment significantly reversed the protective effect of HBSP, as evidenced by the extremely high level of Scr (112.16 ± 12.37 versus 23.83 ± 1.32 μmol/L, HBSP, P < 0.01) and BUN (49.86 ± 2.86 versus 23.83 ± 1.32 mmol/L, HBSP, P<0.01) compared with the HBSP group (Figure 1a).
Evaluation of the renal protection. All mice were subjected to 30 min of ischemia and 48 h of reperfusion except the sham group, and the serum was obtained at 48 h from each group to detect serum creatinine (Scr) and blood urea nitrogen (
Histological assessment in each group was performed in H&E stained sections. Semi-quantitative analysis using a histological scoring system revealed that the tissue structure in the HBSP treated group was well protected, with mild tubular dilation and interstitial edema, fewer protein casts in tubular lumens and cellular infiltration, without tubular epithelial vacuolation or detachment, compared with that in the vehicle group (1.17 ± 0.31 versus 4.50 ± 0.43 per × 200 field, P < 0.01). However, wortmannin inhibited the tissue protection of HBSP (4.17 ± 0.48 versus 1.17 ± 0.31 per × 200 field, P < 0.01) (Figure 1b).
The number of apoptotic cells in tubulointerstitial areas was significantly decreased by HBSP treatment, as determined by the TUNEL (terminal dUTP nick end labeling) positive cells in renal tissue sections (3.83 ± 1.40 versus 24.17 ± 1.45 per × 400 field, P < 0.01, Figure 2a).
Examination of tubular apoptosis and caspases activation in the kidney. (a) Terminal dUTP nick end labeling positive cells were detected (×400 magnification), and the semi-quantitative result showed the renal tubular apoptosis was significantly decreased by the administration of helix B surface peptide (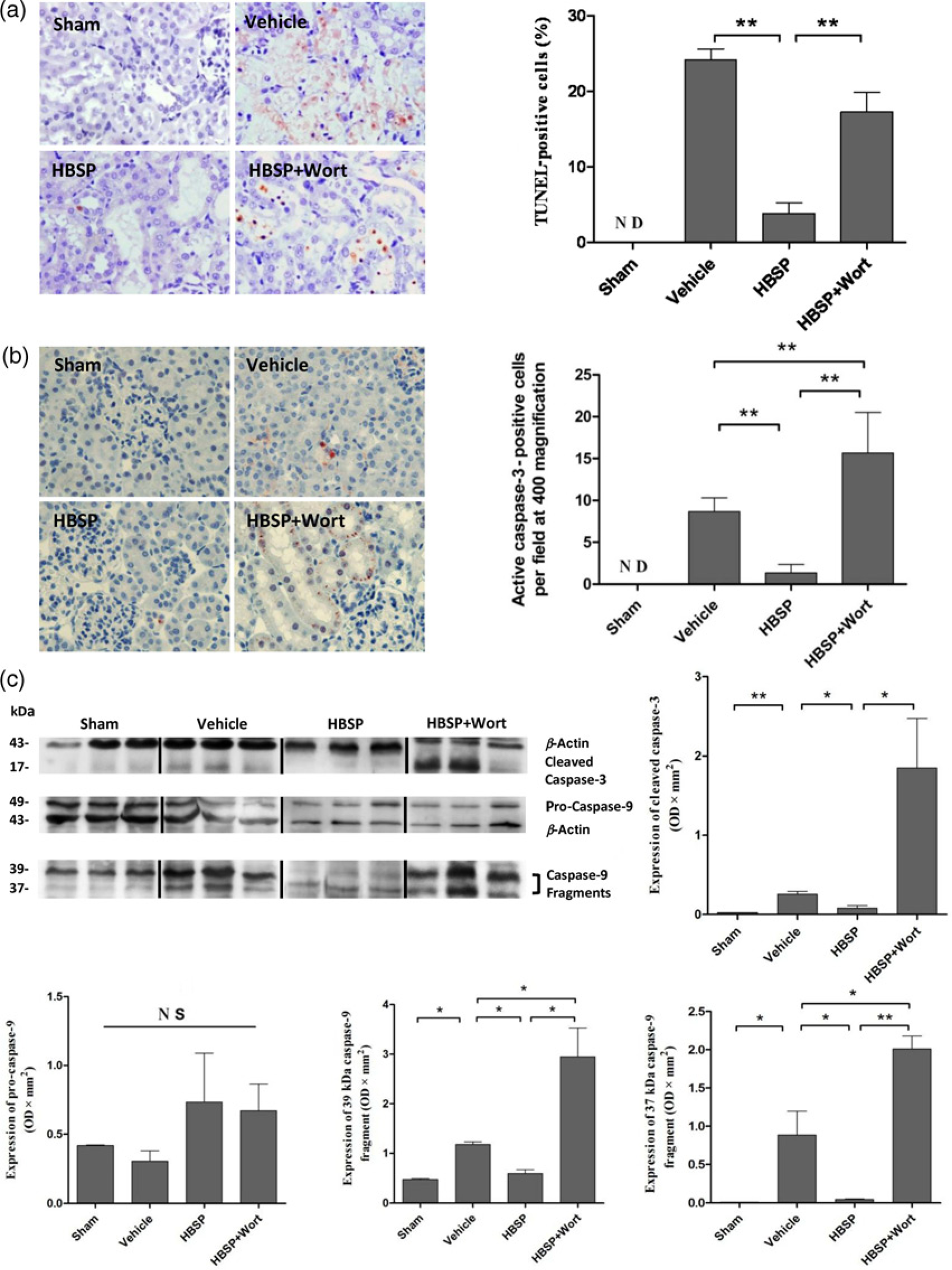
Wortmannin did not further increase Scr and BUN, histological damage and the number of apoptosis in contrast to the vehicle group.
Impact of HBSP on caspase-9 and -3 activation
The distribution of active caspase-3 in the IR injured kidneys was also detected by immunohistochemistry. The active caspase-3 positive cells were hardly seen in the HBSP treated group, but revealed an increase in the renal tubular lumens after IR injury (1.33 ± 0.42 versus 8.67 ± 0.67, P < 0.01, Figure 2b). Wortmannin treatment not only significantly reversed the effect of HBSPbut also further increased the number of active caspase-3 positive cells (15.67 ± 1.98, P < 0.01, Figure 2b) compared with the vehicle group.
Further investigation showed that there were no statistical differences in the expression of 49 kDa full-length pro-caspase-9 among the groups. However, the activated forms, including the 39 and 37 kDa caspase-9 fragments and the 17 kDa cleaved caspase-3 were all markedly reduced by HBSP in comparison to the high level in the vehicle group (39 kDa caspase-9: 0.60 ± 0.07 versus 1.18 ± 0.05 OD × mm2, P < 0.05; 37 kDa caspase-9: 0.05 ± 0.01 versus 0.89 ± 0.31, P < 0.05; caspase-3: 0.08 ± 0.03 versus 0.26 ± 0.04, P<0.05, Figure 2c). Furthermore, the inhibited expression of two active caspase-9 subunits (2.95 ± 0.58, P < 0.05; 2.01 ± 0.17, P < 0.01) as well as active caspase-3 (1.85 ± 0.62, P<0.05, Figure 2c) was reversed and further enhanced by the treatment of wortmannin.
Expression of tissue protective receptor
The protective subunit βcR revealed by immunostaining was constructively expressed on renal tubule cells and was up-regulated by IR injury (94.83 ± 4.19 versus 130.67 ± 13.80 per × 400 field, P < 0.05, Figure 3a).
Expression of the tissue protective receptor in the kidney. (a) The subunit beta-common receptor (βcR) was shown on the renal tubular cells in a great number, and significantly stimulated at injury (× 400 magnification). (b)The 130 kDa band of βcR represented the level of βcR/EPOR heterodimer. The expression of EPOR combined βcR increased during the ischemia reperfusion injury, but remarkably lowered with helix B surface peptide (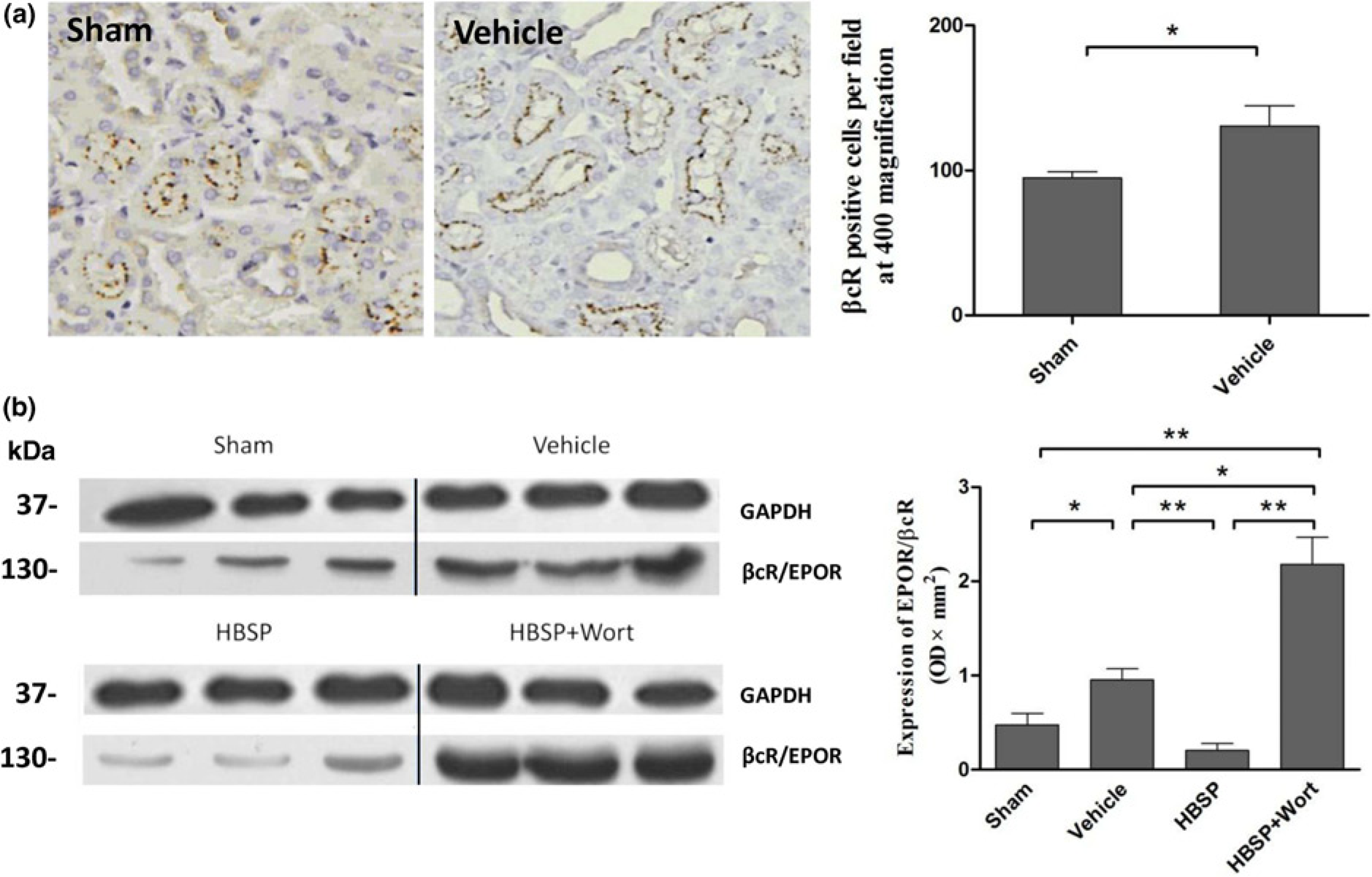
The expression of βcR and EPOR in a heterodimeric form was further detected by the immunoprecipitation assay. As shown in Figure 3b, the EPOR precipitated βcR was clearly increased by IR injury (0.95 ± 0.12 versus 0.47 ± 0.12 OD × mm2, P < 0.05), but fell back markedly upon HBSP treatment (0.20 ± 0.07, P < 0.01). In addition, with the treatment of wortmannin (2.18 ± 0.29), the expression of EPOR precipitated βcR was more than doubled in comparison to the vehicle group (P < 0.05), the HBSP group or the sham group (both P < 0.01, Figure 3b).
Impact of HBSP on PI3K/Akt signaling pathway
The PI3K/Akt pathway inhibitor, wortmannin, was used to further investigate the necessity of the PI3K/Akt pathway in HBSP induced tissue protection. There was no difference in the expression of PI3K p85 among groups (Figure 4b). However, as shown in Figure 4, HBSP treatment did significantly enhance the expression of Akt compared with the vehicle group (0.444 ± 0.044, versus 0.213 ± 0.040 OD × mm2, P < 0.05) as well as the sham group (0.112 ± 0.004, P < 0.01), although IR itself failed to statistically increase the total Akt in comparison to the sham group (Figure 4c).
The change of PI3K/Akt pathway. The expression of 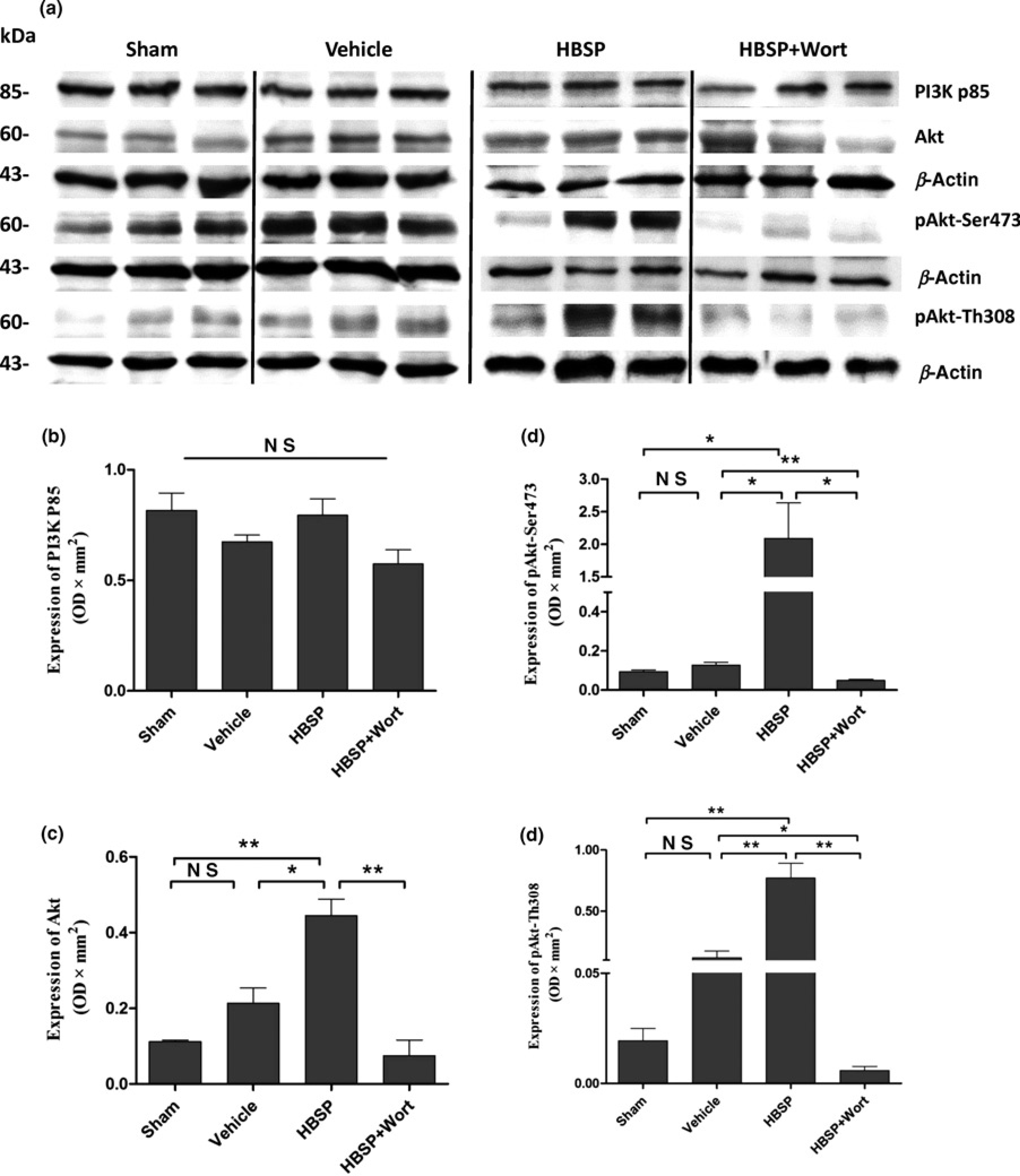
The changes in the phosphorylated Akt represented by phosphorylation of Ser473 (2.089 ± 0.446, HBSP, versus 0.128 ± 0.013, vehicle, P < 0.05; versus 0.094 ± 0.009, sham, P < 0.05, Figure 4e) and Th308 sites (0.770 ± 0.121, HBSP, versus 0.118 ± 0.055, vehicle, P < 0.01; versus 0.019 ± 0.006, sham, P < 0.01, Figure 4c) were consistent with that of Akt.
Wortmannin significantly reversed and inhibited the expression of Akt (0.075 ± 0.041 versus HBSP, P < 0.01) and the phosphorylation in both Ser437 (0.049 ± 0.005 versus HBSP, P < 0.05) and Th308 sites (0.006 ± 0.002 versus HBSP, P < 0.01) caused by HBSP treatment (Figures 4c-e).
Discussion
In this study, we showed that HBSP, a non-erythropoietic peptide derived from EPO, played a protective role by ameliorating renal dysfunction and tissue damage in a murine renal IR injury model. This effect was attributed to the existence of the heterodimeric receptor βcR/EPOR in the kidney, as well as PI3K/Akt signaling pathway stimulated by HBSP for the inhibition of IR-induced apoptosis.
HBSP, derived from the hydrophilic helix B of EPO, was synthesized according to the sequence of amino acids first published by Brines et al.7,8 As an oligopeptide, HBSP possesses characteristics of rapid absorption, distribution and clearance in vivo. It has been reported that HBSP presented the same tissue protective potency as EPO without the erythropoietic effect. 12 Therefore, HBSP is supposed to be a better candidate than EPO for therapeutic application. In this murine bilateral 30 min ischemia and 48 h reperfusion model, it clearly revealed that 8 nmol/kg HBSP given one minute, six hours and 12 h after insult improved renal function and histological changes.
Apoptosis plays an important role in IR injury. Oxygen deficit due to ischemia may exhaust the reserved ATP and disrupt the cell membrane integrity, undermining intra-cellular homeostasis which is necessary for survival.13–15 Moreover, energy abnormality may lead to mitochondria dysfunction, which further initiates the activation of apoptotic mediators.16,17 In this study, a great number of apoptotic cells were found in the tubulointerstitial area of the vehicle treated kidneys, whereas those in the HBSP-treated kidneys were significantly lower. Further investigation showed suppressed activation of caspase-9 and -3 in the HBSP treated kidneys. Caspase-9 is a crucial up-stream initiator enzyme in the caspase cascade. Once activated, the full length caspase-9 was cleaved into 37/39 kDa fragments to catalyze down-stream caspases including the major effector enzyme caspase-3, ultimately resulting in the execution of apoptotic cell death. 18 This study displayed that the 17 kDa subunit of caspase-3 and activated caspase-3 positive cells increased after IR injury, but were significantly inhibited by HBSP. Our result that HBSP alleviated apoptosis in renal injury accords with the antiapoptotic effect of HBSP in other organs. 19
It has been revealed in previous studies that βcR and EPOR combined as the heterodimeric receptor of HBSP to mediate tissue protection. Knock out of βcR exacerbated IR damage in myocardial cells and neurocytes. 6 In this study, immunohistochemistry showed the constitutive expression of βcR in the renal tubular epithelial cells and its up-regulation during IR injury. Moreover, given the necessity of the integrity of βcR with EPOR to perform protective roles, these two subunits in an associated form were identified by the co-immunoprecipitation assay, and the EPOR co-precipitated βcR was also significantly increased by renal IR injury, but decreased by HBSP treatment.
PI3K is an intracellular signaling molecule, composed of a catalytic subunit (p110) and a regulatory subunit (p85). 20 Only when the p85 is phosphorylated and subsequently separated from p110 during activation, can the p110 subunit catalyze and initiate the subsequent phosphorylation of the Akt pathway. 21 A variety of studies have proved that constitutive activation of Akt is potent enough to inhibit the apoptosis in many kinds of cells.22–24 In this study, neither the total nor the phosphorylated Akt was influenced by IR injury alone, but the expression and phos-phorylation of Akt was remarkably increased by HBSP. Moreover, inhibition of the whole pathway by wortmannin even in the presence of HBSP further confirmed the involvement of Akt activation. Therefore, the PI3K/Akt pathway was one of the intracellular signaling pathways that were stimulated by HBSP and led to a protective effect on apoptosis. In addition, administered 10 μg/kg HBSP to rats subjected 30 min renal ischemia and 48 h reperfusion attenuated renal and tubular dysfunction through enhanced phosphorylation of Akt and glycogen synthase 3-β, and decreased NF-κB activity and inflammation. 25 In a heart 45 min ischemia and four hours reperfusion murine model, the expression of βcR/EPOR and the level of phos-phorylated Akt were not significantly changed by IR injury alone, but increased by carbamylated EPO in comparison to the IR group. 26
Another interesting finding among our results was that the expression of heterodimer receptor βcR/EPOR was up-regulated by IR injury, but down-regulated by HBSP. Moreover, the expression of βcR/EPOR was significantly further increased by wortmannin treatment to a level of more than double of that in the IR group. This indicated that the PI3K/Akt pathway might be involved in a negative feedback regulation of βcR/EPOR after 48 h reperfusion in this study. Moderate IR injury increased the expression of this protective heterodimer receptor. Without HBSP, the up-regulated βcR/EPOR by IR alone failed to fully initiate the PI3K/Akt pathway, whereas with HBSP treatment, the PI3K/Akt pathway adapter proteins were activated and subsequently involved in the tissue protection such as inhibiting apoptosis.
Nevertheless, there may be a time differentia between the expression of βcR/EPOR and PI3K/Akt pathway adapter proteins as the level of βcR/EPOR might be higher at the early stage of reperfusion with HBSP treatment in order to activate the PI3K/Akt pathway, 26 and were then reversed into a low level after 48 h reperfusion, but the expression of PI3K/Akt pathway adapter proteins was still remained at a high level. This needs a further study with more time points to confirm. Furthermore, interruption of PI3K/Akt signaling pathway by wortmannin led to remarkable increasing of the heterodimeric receptor, which indicates that the feedback regulation of βcR/EPOR requires an involvement of PI3K/Akt signaling pathway. In addition, the activation of caspases was not only reversed, but also further raised by wortmannin, although the level of Scr and BUN, the score of renal tissue damage, as well as the number of apoptosis did not further increase. In this study, therefore, wortmannin might have side-effects, which need to be further explored.
The justification of using GAPDH and β-actin as loading controls cannot be neglected, because GAPDH appeared not to be influenced by experimental conditions, but β-actin seemed to be affected. Caron et al. demonstrated that renal ischemia strongly increased the expression of β-actin and α-tubulin, especially in nuclear fractions. The data suggest that increased and redistributed β-actin and microtubule cytoskeleton components participated in adaptive responses to cellular lesions. 27 In this study, the loading controls were not specifically chosen for desired purposes, apart from referring the molecular weight of target proteins to comparable loading controls and Western blots generated by two laboratories according to their routing practice. In addition, there are no ‘gold standards’ for selecting loading controls; and relative ratios normalized by loading controls, rather than absolute values, were used for the comparison of target proteins between groups. Therefore, both GAPDH and β-actin should be acceptable, although GAPDH was more constant than β-actin and could be a more suitable loading control for this experimental setting.
In summary, HBSP demonstrated convincing protective effects on renal structure and function in IR injury by ameliorating the activation of caspase-9, -3 as well as apoptosis. The PI3K/Akt pathway was one of the involved signaling pathways, and activated by HBSP via heterodimeric βcR/EPOR. The therapeutic effects of HBSP in this study suggest that HBSP is a better candidate for renal protection.
Footnotes
Acknowledgements
The authors also thank the grants provided by the National Nature Science Foundation of China (No. 81270832, 81270833 and 81170689) and Science and Technology Commission of Shanghai Municipality (No. 12ZR1405500).