
Abstract
Thyroid hormone deficiency during perinatal development results in significant alterations in neurological functions. The relationship between such events and brain metabolism is not completely understood. The aim of this study was to investigate the effects of hypothyroidism on leucine, mannose, glucose and lactate metabolism in rat cerebellar slices. Experimental hypothyroidism was induced by exposing mothers and pups to propylthiouracil (PTU) until weaning - postnatal day 21. Metabolic analyses were performed in postnatal day 10 (PND10) and 21 (PND21) animals. A matching group of animals received the same oral treatment also after weaning until adulthood PND60 with T3 supplement during lactation (P1-P21). In PND21 animals, PTU treatment significantly increased the rate of leucine oxidation to CO2, although glucose and lactate oxidations were not affected. PTU treatment also increased the oxidation of leucine to CO2 at PND60 (adult animals). PND10 hypothyroidism animals showed a decrease in conversion of mannose to glycolipids and glycoprotein compared with the control group. However, PTU treatment increased the conversion of mannose to glycolipids and glycoprotein in PND21 animals. The replacement of T3 normalized mannose and leucine metabolism in adult rats. These results indicate that deficits in thyroid hormones during lactation could delay or alter brain development and metabolism.
Introduction
Thyroid hormones are essential for the appropriate development of the mammalian central nervous system (CNS). 1 They regulate several neurodevelopmental processes, including neurogenesis, myelination, dendrite proliferation and synapse formation.2,3,4 Hypothyroidism during perinatal development results in profound neurological deficits at least in part due to impaired brain metabolism. 5 Several studies indicate that thyroid hormones modulate the expression and/or activity of crucial metabolic players in the brain, such as glucose transporters, 6 tyrosine hydroxy-lases, 7 glutamine synthetase 8 and enzymes of ketone body metabolism. 9
Propylthiouracil (PTU) has been widely used as an antithyroid agent. PTU inhibits both the synthesis of thyroid hormones and the conversion of thyroxine (T4) to its active form triiodothyronine (T3), causing thyroid disorders. 10 During the first few months of pregnancy, the fetus does not produce thyroid hormones itself and therefore depends totally on the supply of thyroid hormones from the mother. 11 Thus, developmental exposure to PTU normally causes delayed development and reduced growth, persistent neurobehavioral changes including increased spontaneous activity and impaired memory function. 12
The brain requires nutrients such as amino acids, glucose and other sugars and lactate to be used in metabolic pathways and for developmental processes. The branched chain amino acid (BCCA) leucine is rapidly transported across the blood–brain barrier 13 to brain tissue where it may be used as a source of energy or alternatively as a nitrogen donor. 14 Accordingly, the brain has a marked activity of BCAA aminotransferase,15,16 Since BCCA are essential amino acids, their uptake into neural cells is crucial for proper protein and neurotransmitter synthesis.17,18
Mannose, a sugar monomer, is involved in an extensive series of metabolic pathways by being incorporated into glycoproteins and glycolipids. 19 In the brain, mannose can be converted to fructose, which is rapidly incorporated into the glycoproteins of nerve endings 20 and brain lipids from Gaunosine diphosphate-mannose. 21 High-mannose oligosaccharides are considered essential components influencing neuroplasticity.22,23 Although the relevance of thyroid hormones for normal brain physiology is well documented (protein and neurotransmitter synthesis, energetic metabolism),24,25 studies regarding the effects of hypothyr-oidism on brain metabolism are still limited.
In this study we aimed to investigate the time profile effect of PTU-induced hypothyroidism on postnatal metabolism of leucine, mannose, glucose and lactate. The results suggest that hypothyroidism impairs amino acid metabolism and delays mannose incorporation into glycolipids and glycoproteins during lactation.
Material and methods
Chemicals
Chloroform, formic acid and methanol were obtained from Merck SA (Porto Alegre, RS, Brazil). Sodium hydroxide was purchased from J.T. Baker Chemical Company (Phillisburg, NJ, USA). L[U-14C]leucine (323 mCi/mmol), D[U-14C]glucose (250–360 mCi/mmol), L[U-14C]lactate (56–186 mCi/mmol), D[2-3H]mannose (27.4 Ci/mmol) were from Perkin-Elmer (Boston, MA, USA). PTU was obtained from Sigma-Aldrich, St Louis, MO, USA.
Animals and treatments
Adult Wistar rats (male and female), 90-d old, from our own breeding colony, were maintained at 22±2°C (mean 6 SD), on a 12:12-h light:dark cycle, with free access to food (Nuvilab CR-1 Nuvital, Curitiba, Paraná, Brazil) and water. The breeding regimen consisted of grouping three virgin females with 1 male for five days. Pregnant rats were selected and housed individually in opaque plastic cages.
The experimental hypothyroidism model was performed using PTU (Sigma-Aldrich, St Louis, MO, USA) in drinking water (0.05% w/v) according Gravina et al.26 The solution was measured to assess the water consumption of animals and replaced daily with fresh PTU solution. PTU enters the bloodstream and passes through the mother's breast milk to the rat pups. Under normal conditions, the enzyme thyroperoxidase covalently binds iodide to thyro-globulin, ultimately resulting in the formation of thyroxine. However, with the addition of PTU, thyroperoxidase actions are blocked, resulting in a reversible model of thyroid hormone deficiency.
On the first day after parturition (postnatal day [PND] 1), dams were randomly assigned to one of three groups – control (euthyroid), hypothyroid (PTU) and reverted hypothyroid (PTU + T3). Pups (8 per litter) were maintained with their mothers during the treatment period (PND10–21, lactational period). An additional group (PND60) received PTU treatment until P22; the pups were then weaned at P25, separated by gender and housed in groups of 3–4 animals until PND60 (post-lactational period).
Dams from the hypothyroid group received PTU diluted in tap water (0.05% w/v), ad libitum, during the lactational period (PTU group). The control (euthyroid) group included weanling rats whose mothers received just tap water, ad libitum, during the lactational period.
Dams from the reverted hypothyroid (PTU + T3) received PTU diluted in tap water (0.05% w/v), ad libitum, during the lactational period (PTU group) and pups received daily injections of T3 (daily subcutaneous injections, 20 μg/kg, dissolved in 4 mmol/L NaOH, 10 mL/kg). Control pups receive vehicle injection (4 mmol/L NaOH, 10 mL/kg). Liquid and solid ingestions by mothers were monitored daily.
Measurement of CO2 production, lipid and protein synthesis
The animals were killed by decapitation. The cerebellum was isolated, kept at 4°C, weighed and cut into 0.3-mm slices using a McIlwain tissue chopper. All experiments were conducted in accordance with the Guiding Principles in the Use of Animals in Toxicology, adopted by the Society of Toxicology in July 1989 and all experiments were approved by our ethics committee for animal use at the Federal University do Rio Grande do Sul.
CO2 production
For the measurement of lipid or protein synthesis and CO2 production, cerebellum slices (between 60 and 70 mg) were incubated in 1.0 mL Krebs Ringer bicarbonate (KRb) buffer pH 7.4, containing either (i) 5.0 mmol/L D-mannose + 0.2 mCi D[2-3H]mannose (ii) 5.0 mmol/L D-glucose + 0.2 mmol/L Lleucine + 0.2 μCi L[U-14C] leucine; (iii) 10.0 mmol/L L-lactate + 0.2 μCi L[U-14C]lactate; (iv) 5.0 mmol/L glucose + 0.2 μCi D[U-14C]glucose. Before incubation, the reaction medium was gassed with a 95% O2: 5% CO2 mixture for one minute. Flasks were sealed with rubber caps, and the slices were incubated at 37°C for one hour in a Dubnoff metabolic shaker (60 cycles/ min) according to the method of Dunlop et al.27 The incubations were stopped adding 0.25 mL 50% tricarboxylic acid (TCA) through the rubber cap. Then 0.20 mL of 1 mol/L sodium hydroxide was injected into the central wells. The flasks were shaken for an additional 30 min at 37°C to trap CO2. Afterwards, the contents of the central well were transferred to vials and assayed for CO2 radioactivity in a liquid-scintillation counter. The flask contents were homogenized and transferred to tubes. 28
Lipid and protein synthesis
After centrifugation, the precipitate was washed 3 times with (1 mL) 10% TCA, and lipids were extracted with chloroform/methanol (2:1). The chloroform/methanol phase was evaporated in vials and radioactivity was measured. The resulting precipitate was dissolved in concentrated formic acid and radioactivity was measured. This radioactivity represents protein synthesis from radioactive amino acids. All the results were expressed with respect to the initial specific activity of the incubation medium.
The CO2 production rate and the lipid and protein synthesis were constant over 30, 60 and 90 min of incubation. 29
Statistical analyses
Biochemical data are expressed as the mean±SEM. All analyses were performed using the Statistical Package for the Social Sciences (SPSS 16.0, SPSS Inc., Chicago, IL, USA) software. P values were considered significant when P < 0.05. Differences in experimental groups were determined by one-way analysis of variance, followed by the Tukey multiple range test or t test.
Results
PTU-induced hypothyroidism had no effect on mean weight gain from PND1–7; however, weight gain from PND7–60 was significantly lower when compared with control animals. Weight gain in animals treated with T3 was similar to that in the PTU group, being different only in PND14 – 21 (Table 1; P < 0.05). The average weight of the brain and cerebellum from animals in the PTU and PTU + T3 groups were significantly lower than control animals.
Effects of hypothyroidism (PTU) on the body and central nervous system weight in (postnatal day 10) PND10, PND21 and PND60 in experimental groups

Values as expressed as mean ± SEM (n = 8 per group. One-way analysis of variance, followed by the Tukey multiple range test.
Control >PTU and PTU + T3 (P < 0.05);
PTU <control and PTU + T3 (P< 0.05)
Figure 1 shows the oxidation of leucine, glucose and lactate to CO2 in cerebellar slices from PND10 and PND21 rats. PTU treatment increased leucine oxidation in PND10 and PND21 by approximately 150% and 200%, respectively, (Figure 1a; P < 0.05) but had no effect on glucose and lactate oxidation (Figures 1b and c).
Effects of hypothyroidism (PTU) on the oxidation of leucine, glucose and lactate to 14CO2 in cerebellum slices from postnatal day 10 (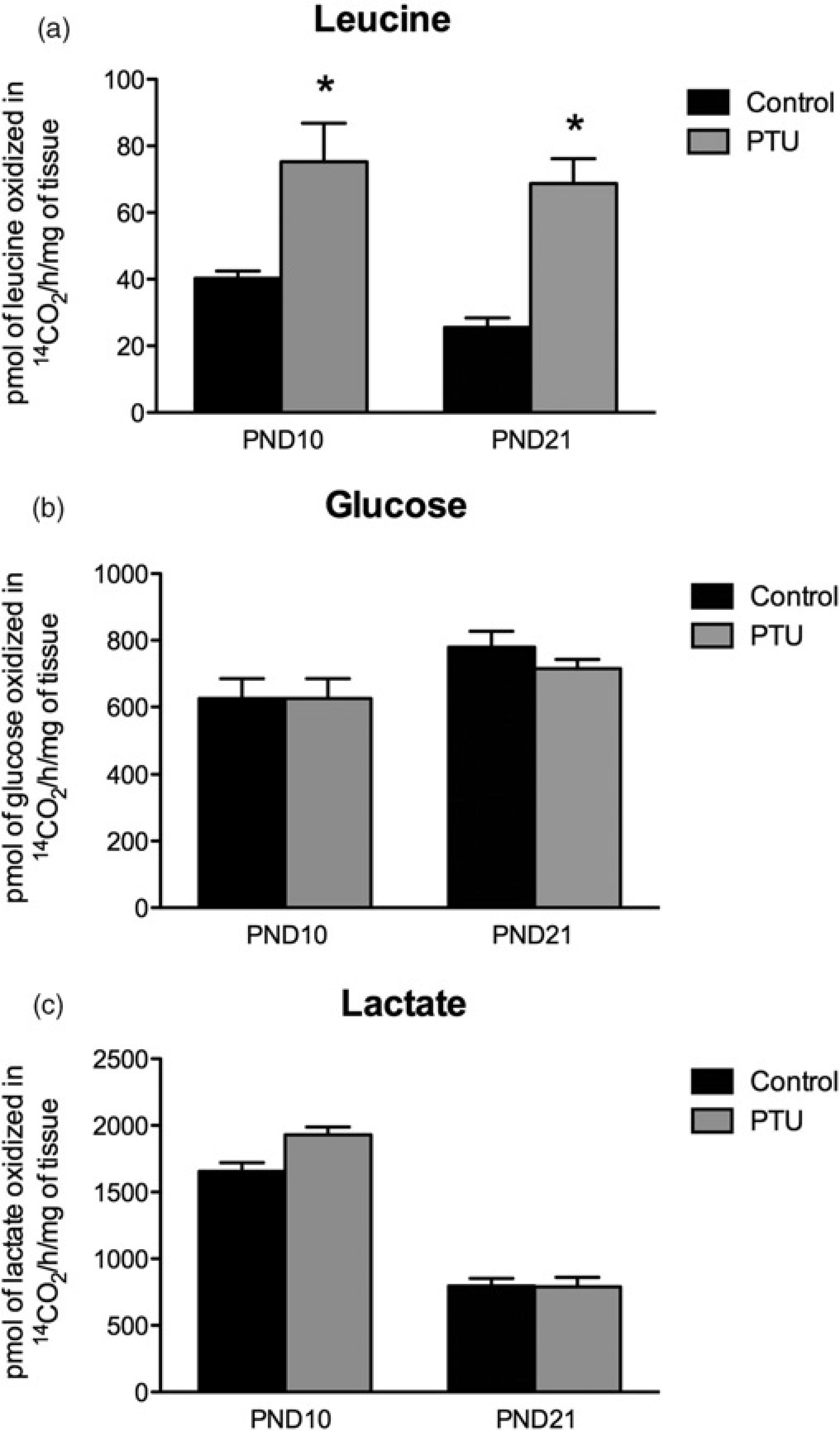
To analyze if the replacement of T3 during development could affect adulthood metabolism, we analyzed oxidation in PND 60 cerebellar slices. PTU treatment increased cerebellar leucine oxidation in PND60 (Figure 2a; P < 0.05); however, the replacement of T3 normalized the oxidation of leucine in PTU treated animals (Figure 2a; P < 0.05). Glucose and lactate oxidations in adulthood were not affected by either PTU or T3 replacement treatments (Figures 2b and c).
Effects of hypothyroidism (PTU) on the oxidation of leucine, glucose and lactate to 14CO2 in cerebellum slices from postnatal day 60 (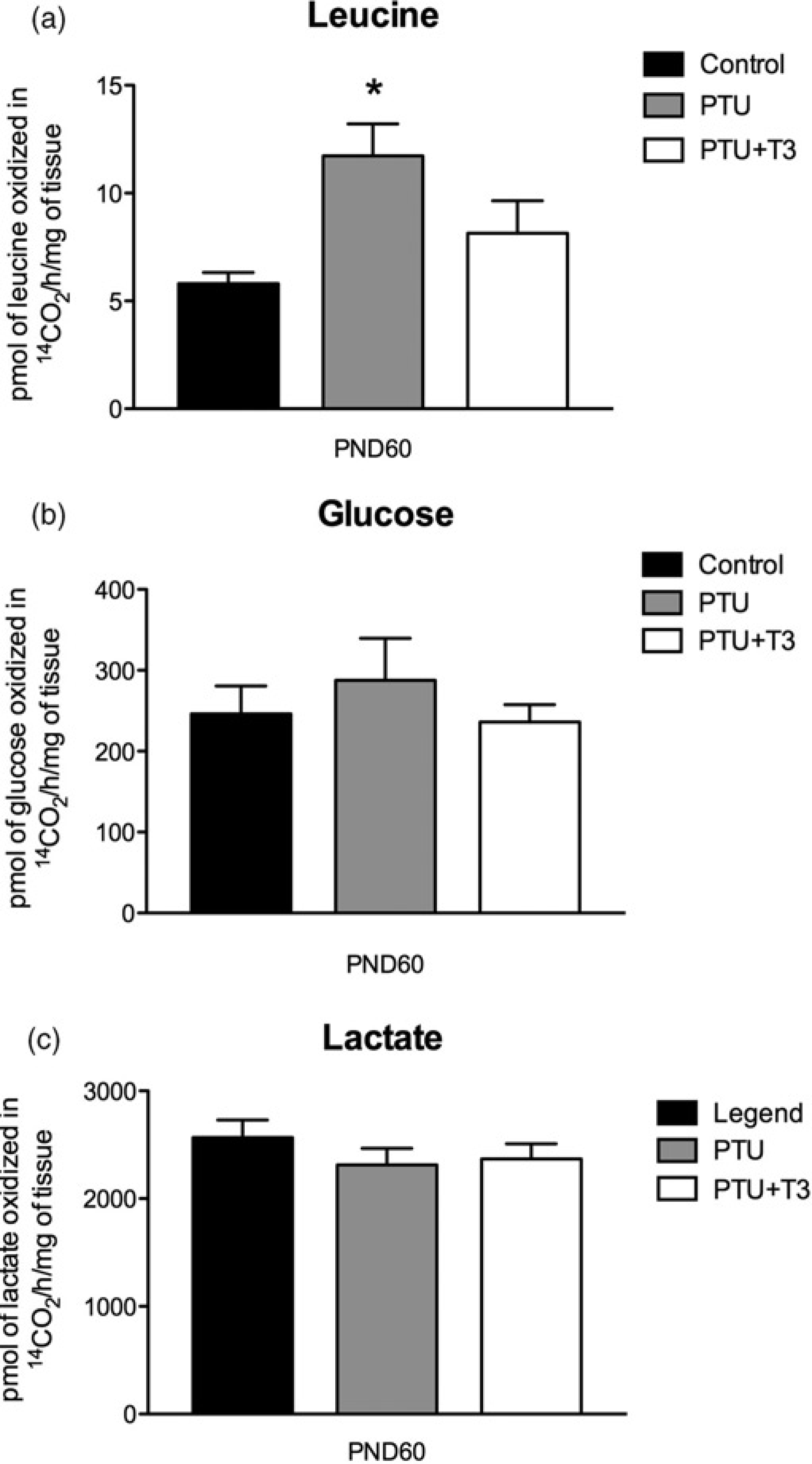
Next, we evaluated the effect of hypothyroidism on mannose metabolism. Figure 3 shows the incorporation of mannose into lipids and proteins in cerebellar slices. The incorporation of mannose into lipids and protein significantly decreased in cerebellar slices from PND10 animals treated with PTU, while in PND21 and PND60 there was an increase in incorporation of mannose into lipids and protein (Figure 3a and b, P < 0.05). The replacement of T3 during development normalized the incorporation of mannose to lipids and protein in PND60-PTU rats (Figure 3a and b, P < 0.05).
Effects of hypothyroidism (
Discussion
Here we showed that PTU affected cerebellar metabolism by increasing leucine oxidation and decreasing mannose incorporation into lipids and proteins. These data suggest impaired brain development and reinforce the relevant role of thyroid hormones during perinatal period for normal brain development.
PTU treatment interferes with the incorporation of iodine into tyrosyl residues of thyroglobulin (TG) and inhibits the coupling of iodotyrosyl residues to form iodothyronine, thus inhibiting the synthesis of thyroid hormones. 30 When PTU is administered to mothers during pregnancy or lactation periods, the fetuses are affected by hypothiroidism and its undesirable consequences because they are dependent of mother's supply. 11 One important issue regarding this PTU-model is the fact that after cessation of medication the rats are able the re-establish thyroid hormone levels. Indeed, a recent study addressing PTU-induced developmental hypothyroidism showed a significant decreased serum T3 and T4 levels in rats at PND10 and PND21 compared with controls which returned to similar levels as controls at PND90. 31
Leucine crosses the blood-brain barrier rapidly,
32
and therefore is a potential nitrogen source for glutamate and glutamine synthesis. One-third of the
The energetic contribution of the sugar mannose as an energetic substrate for brain oxidative metabolism is low; however, its major physiological role in the CNS appears to be linked to its incorporation into lipids and proteins to build up cell-surface molecules involved in cell migration, neurite outgrowth during development 36 and, in the maintenance of synaptic activity in the adult CNS.37,38 Of particular importance is that glycoproteins and glycolipids play crucial roles in the processes of brain development and neural communication. 39 Our results showed that PTU-induced hypothyroidism during the perinatal period caused a decreased capacity to incorporate mannose into glycoproteins (20%) and glycolipids (35%) at PND10; however, a higher incorporation of mannose into glycoproteins (58%) and glycolipids (18%) was observed at PND21 and PND60, suggesting a delayed formation and assembly of these molecules into the framework structures essential to synaptogenesis and myelinization. This postulation is reinforced by evidence suggesting that hypothyroidism in developing rats impairs synaptic neuro-transmission and causes devastating and permanent deleterious effects on neurological functions. 40 Here, the replacement of T3 reversed PTU-induced decrease in mannose incorporation in glycolipids and glycoproteins, pointing to the importance of this hormone on normal brain physiology.
In conclusion the data of the present study provide evidence that PTU-induced hypothyroidism during the perinatal period elicits disturbances in leucine oxidation and delays mannose incorporation into glycolipids and glyco-proteins in the rat cerebellum. These alterations may delay brain maturation secondary to hypothyroidism.
Footnotes
Acknowledgements
This study was supported by grants from CNPq, PROPESQ/UFRGS, IBN-Net/FINEP # 01.06.0842-00 and INCT-EN/CNPq.