
Abstract
Metabolic syndrome is often accompanied by development of hepatic steatosis and less frequently by non-alcoholic fatty liver disease (NAFLD) leading to non-alcoholic steatohepatitis (NASH). Replacement of corn oil with medium chain triacylglycerols (MCT) in the diets of alcohol-fed rats has been shown to protect against steatosis and alcoholic liver injury. The current study was designed to determine if a similar beneficial effect of MCT occurs in a rat model of NAFLD. Groups of male rats were isocalorically overfed diets containing 10%, 35% or 70% total energy as corn oil or a 70% fat diet in which corn oil was replaced with increasing concentrations of saturated fat (18:82, beef tallow: MCT oil) from 20% to 65% for 21 days using total enteral nutrition (TEN). As dietary content of corn oil increased, hepatic steatosis and serum alanine amino transferases were elevated (P < 0.05). This was accompanied by greater expression of cytochrome P450 enzyme CYP2E1 (P < 0.05) and higher concentrations of polyunsaturated 18:2 and 20:4 fatty acids (FA) in the hepatic lipid fractions (P < 0.05). Keeping the total dietary fat at 70%, but increasing the proportion of MCT-enriched saturated fat resulted in a dose-dependent reduction in steatosis and necrosis without affecting CYP2E1 induction. There was no incorporation of C8-C10 FAs into liver lipids, but increasing the ratio of MCT to corn oil: reduced liver lipid 18:2 and 20:4 concentrations; reduced membrane susceptibility to radical attack; stimulated FA β- and ω-oxidation as a result of activation of peroxisomal proliferator activated receptor (PPAR)a, and appeared to increase mitochondrial respiration through complex III. These data suggest that replacing unsaturated fats like corn oil with MCT oil in the diet could be utilized as a potential treatment for NAFLD.
Introduction
Medium chain triacylglycerols (MCT oil) consists of saturated fatty acids (FA) of chain length consisting of predominantly 8–10 carbons. 1 Unlike long-chain fats, MCTs are absorbed directly from the diet into the hepatic portal vein, and are thought to enter the liver cells and mitochondria by minimal use of normal FA transport systems or by diffusion where they are rapidly degraded to increase ther-mogenesis.2,3 In addition, MCT FAs are poorly esterified into cellular triacylglycerols in adipose tissue and do not induce adipocyte differentiation 2 and have been suggested to increase satiety and reduce food intake. 4 As a result, diets high in MCT have been utilized clinically to prevent obesity.2–6 Interestingly, MCT has also been shown to prevent alcohol-induced liver injury in animal models7–10 and clinically in patients with cirrhosis. 11 The protective effect of MCT on alcoholic liver injury has variously been suggested to involve increased FA degradation, decreased lipid peroxidation, reduced gut permeability and inhibition of endotoxin-induced Kupffer cell activation.7–10 Since alcoholic steatohepatitis and non-alcoholic fatty liver disease/ non-alcoholic steatohepatitis (NAFLD/NASH) share a very similar pathology, it has been suggested that dietary supplementation with MCT might have a similarly beneficial effect on NAFLD/NASH pathology. However, to date, this has not been extensively investigated. A study by Charles Lieber's group in 2008 12 suggested that complete replacement of dietary long-chain triacylglycerols (LCT) with MCT could dramatically ameliorate NASH development produced by ad libitum feeding of a liquid diet containing 70% fat for 21 days as a result of improved insulin and adipokine status, reduced steatosis, reduced inflammation and increased hepatocyte proliferation. However, interpretation of these data was complicated by the substitution of MCT for two different sources of LCT, corn oil and olive oil, and by reductions in caloric intake produced by MCT. 12 In this same study, the authors observed increased rather than decreased hepatotoxicity when MCT only partly replaced LCT unless the MCT substitution was accompanied by caloric restriction. 12
We recently developed a new rat model of NAFLD in which overfeeding a liquid diet high in the polyunsaturated LCT corn oil via an intragastric cannula (total enteral nutrition, TEN), resulted in dose- and time-dependent progression of NAFLD to NASH. 13 The advantage of this model is that diets of substantially different composition can be overfed isocalorically to produce obesity, metabolic syndrome and NASH while eliminating the variable of different levels of caloric intake between groups. We utilized the model to demonstrate the importance of oxidative stress in NASH progression, 14 and herein examine the effects of iso-caloric substitution of corn oil with an MCT-enriched mixture of saturated fats (MCT:beef tallow, 82:18) on body composition, development of liver pathology, induction of CYP2E1, liver FA composition and hepatic lipid homeostasis.
Materials and methods
Animals and experimental design
Male Sprague–Dawley rats (175 g) were purchased from Harlan Sprague-Dawley (Indianapolis, IN, USA). Animals were housed in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) approved animal facility. Animal maintenance and experimental treatments were conducted in accordance with the ethical guidelines for animal research established and were approved by the Institutional Animal Care and Use Committee at the University of Arkansas for Medical Sciences. Rats had an intragastric cannula surgically inserted and were allowed seven days to recover before infusion of TEN diets as described previously.
13
Animals had ad libitum access to water throughout the experiments. Rats were randomly assigned to groups of n = 7–8 and were overfed 220 kcal/ kg3/4/d diets (17% above that required to match the growth of ad libitum chow fed rats) containing 10%, 35% or 70% corn oil (CO) for 21 days to produce obesity and dose-dependent development of NAFLD. In the other experimental groups the total % energy from fat was kept constant at 70%, but a saturated fat mixture of 82:18 MCT oil:beef tallow (
Liver pathology
Steatosis as determined by Oil Red O staining of lipid droplets was quantified by image analysis as described pre-viously. 13 Triglyceride was extracted from whole liver homogenates with chloroform:methanol (2:1, v:v) and analyzed using Triglyceride Reagent (IR141; Synermed, Westfield, IN, USA). Inflammation was assessed by measurement of tumor necrosis factor (TNFα).
mRNA expression using realtime reverse transcriptase polymerase chain reaction (RT-PCR) as described below. 14 Necrosis was assessed biochemically by measurement of serum alanine amino transferase (ALT) activity at sacrifice using the Infinity ALT liquid stable reagent (Thermo Electron Corp., Waltham, MA, USA) according to sthe manufacturer's protocols. Cellular proliferation was measured indirectly by realtime RT-PCR assessment of mRNA encoding Ki67 which is only expressed in dividing cells 15 and by Western immunoblot analysis of the hepatic expression of proliferating cell nuclear antigen (PCNA) as described previously. 16
Biochemical analysis
Western immunoblot analysis of apoprotein expression for CYP2E1 and CYP4A1 was conducted as previously described and normalized against total protein loaded based on ponceau staining of the membranes. 16 CYP2E1 was a gift from the laboratory of Dr Magnus Ingelman-Sundberg (Karolinska Institute, Stockhom, Sweden). 17 CYP4Al was detected using a polyclonal sheep antibody to rat CYP4A1 18 which was a gift from Dr Gordon Gibson (University of Surrey, Guildford, UK). CYP2E1 activity was assessed directly by measurement of microsomal p-nitrophenol hydroxylase activity. 19 Susceptibility of cellular membranes to attack by free radicals generated by CYP2E1 was determined by assessing the rate of formation of thiobarbituric reactive lipid peroxidation products in microsomes incubated with carbon tetrachloride. 20 Lipid FA profile analysis of liver homogenates was conducted by gas chromatography mass spectrometry as previously described in our laboratory. 21 Peroxidizability index of liver free FA, triglycerides and phospholipid fractions was calculated based on FA profiles as described by Lambert et al. 22 Mitochondrial protein extracts were prepared using a Mitochondrial Isolation kit for Tissue (Pierce, Rockford, IL, USA) and immunoblotting was performed for oxidative phosphorylation complexes I–V (MitoSciences, Eugene, OR, USA). Mitochondrial numbers were estimated indirectly by realtime RT-PCR analysis of expression of the mitochondrial transcription factor mtTFAM mRNA. In addition, sirtuin (SIRT)3 mRNA expression and expression of the uncoupling protein UCP2 mRNA were analyzed by realtime RT-PCR. 23
Realtime RT-PCR
Total RNA was extracted from livers using TRI Reagent and cleaned using RNeasy mini columns (Qiagen, Valencia, CA, USA). RNA quality was ascertained spectrophotometrically (ratio of A260/A280) and also by checking ratio of 28S to 18S ribosomal RNA using the RNA Nano Chip on a 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). Total RNA (1 μg) was reverse transcribed using the iScript Reverse Transcription kit (Biorad Laboratories, Hercules, CA, USA) according to the manufacturer's instructions. The reverse transcribed cDNA (10 ng) was utilized for realtime PCR using the 2 × SYBR green master mix and monitored on a ABI Prism 7000 sequence detection system (Applied Biosystems, Foster City, CA, USA). Gene-specific probes described previously 13,14,20,21,23,24 were designed using Primer Express™ Software (Applied Biosystems, CA and the relative amounts of gene expression were quan-titated using a standard curve according to the manufacturer's instructions. Gene expression was normalized to 18S rRNA as described previously. 13
EMSA for assessment of peroxisome proliferator activated receptor alpha (PPARα) binding
Complementary oligonucleotide probes specific for the peroxisome proliferator response element (PPRE) on the acyl CoA oxidase gene were synthesized by Integrated DNA Technologies Inc. (Coralville, I A, USA). Hepatic nuclear extracts were used for electrophoretic mobility shift assay (EMSA) analysis of PPARα binding as described previously. 24
Statistical analysis
Data for continuous outcomes are presented as mean ± standard errors of the mean (SE). One-way analysis of variance (ANOVA) was used to test overall mean differences of each outcome across corn oil concentrations. Post hoc comparisons of means were performed using an all-pairs-Tukey-Kramer comparison test and considered significant if P ≤ 0.05. The Jonckheere-Terpstra test, a non-parametric trend test, was used to test for a significant overall increasing or decreasing change in each outcome with varying corn oil concentrations. Statistical analyses were performed using the Stata statistical package version 12.0 (Stata Corp, College Station, TX, USA).
Results
Isocaloric feeding of MCT dose-dependently reduces adiposity
Overfeeding diets containing different proportions of CO resulted in comparable body weight and obesity with an ending body composition of ~20% body fat (Table 1). Isocaloric substitution of
Effects of dietary fat composition on body weight and adiposity

CO, com oil,
Data are mean ± SEM. Means with different superscripts are significantly different P < 0.05 a < b < c
Determined by nuclear magnetic resonance
CO dose-response
Liver pathology is reduced by increasing dietary content of MCT
Increasing the %CO in the diet resulted in a dose-dependent development of NAFLD, as indicated by elevated hepatos-teatosis and serum ALT concentrations (Tables 1 and 2). Although liver size was slightly reduced (P < 0.02), appearance of lipid droplets as measured by Oil Red O staining (Figure 1) was paralleled by increased liver triglyceride content in 70% CO fed rats compared with 10% CO fed rats (trend: P<0.01; Table 2). Analysis of alterations in the fatty acid (FA) composition of hepatic lipid fractions: free FA, triglycerides and phospholipids is shown in Table 3. The major FAs in the free FA and triglyceride fractions were 16:0,18:1,18:2 and 20:4. In the phospholipid fraction, 18:0 was also present in a significant proportion of the lipid. Other FA species made up only a small portion of the lipid fractions (data not shown). Increasing dietary %CO resulted in a dose-dependent rise in polyunsaturated 18:2 (linoleate) content in all three lipid fractions (trend: P < 0.01). In addition, the amount of arachidonate (20:4) was greater in the free FA and triglyceride fractions (trend: P < 0.02). Small elevations were observed in the total amount of 16:0 and 18:1 FA triglycerides and 18:0 phospholipids as the %CO in the diet increased (P < 0.05) along with a greater total triglyceride concentration in the liver. However, the proportion of 16:0 and 18:1 FA were reduced relative to the proportion of 18:2 FA. This resulted in dose-dependent increases in the peroxidizability index of the free FA and triglyceride fractions (trend: P < 0.0001) and greater peroxidizability of the phospholipid fraction from the 70% CO group relative to the 35% CO group (P < 0.05; Table 4). Other FA species were present in hepatic lipid fractions in small amounts, but even when combined, contributed to only a small percentage of the total (data not shown). Development of steatosis was accompanied by induction of the cytochrome P450 enzyme CYP2E1 at the level of apoprotein and activity, which is a known source of reactive oxygen species
25
(P < 0.05; Figures 2 and 3). Steatosis was also associated with greater serum ALT values, indicative of the development of necrosis (P < 0.05; Table 2). After this relatively short period of overfeeding of high polyunsaturated fat diets, there was a tendency to increase expression of the inflammatory marker TNFα mRNA, but this did not reach statistical significance (trend: P = 0.08; Table 2). Increasing dietary CO from 10% to 70% had no significant effect on hepatic expression of Ki67 mRNA (Table 2) but significantly suppressed expression of PCNA protein (P < 0.05; Figure 4) suggesting a small decrease in cellular proliferation. Isocaloric substitution of CO in 70% fat diets with MCT oil-rich saturated fat resulted in a dose-dependent increase in liver weight (trend P < 0.0001), but a dose-dependent decrease in hepatic pathology including disappearance of lipid droplets (Figure 1, Table 2), decreased triglyceride content, reduced ALT values and decreased expression of Ki67 mRNA (trend: P < 0.01; Table 2) and PCNA protein (Figure 4). Interestingly, no incorporation of C8-C12 FAs was observed in any of the three hepatic lipid compartments after
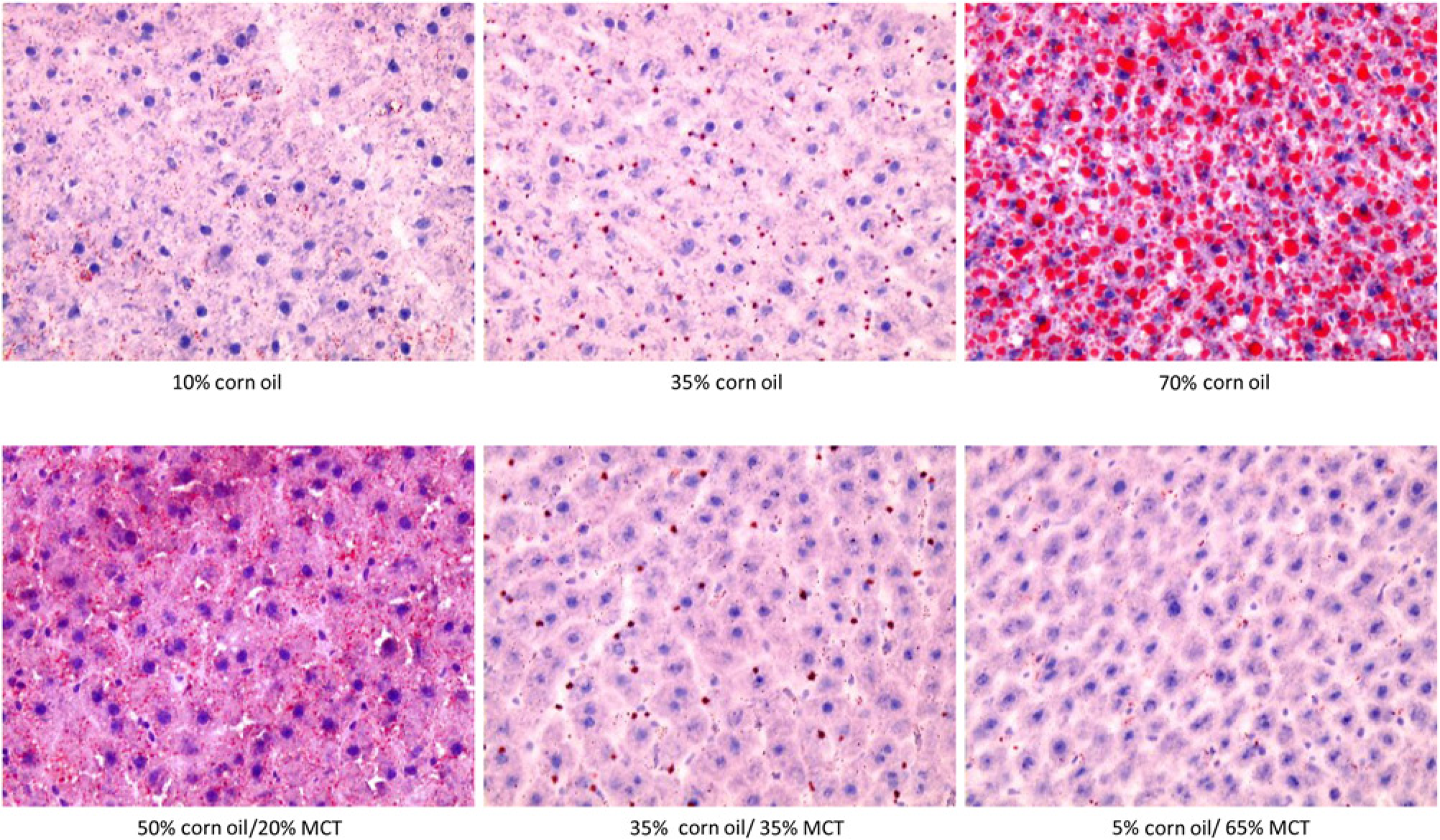
Representative Oil Red O stained liver sections illustrating the effects of dietary fat composition on accumulation of hepatic lipid droplets in the TEN model
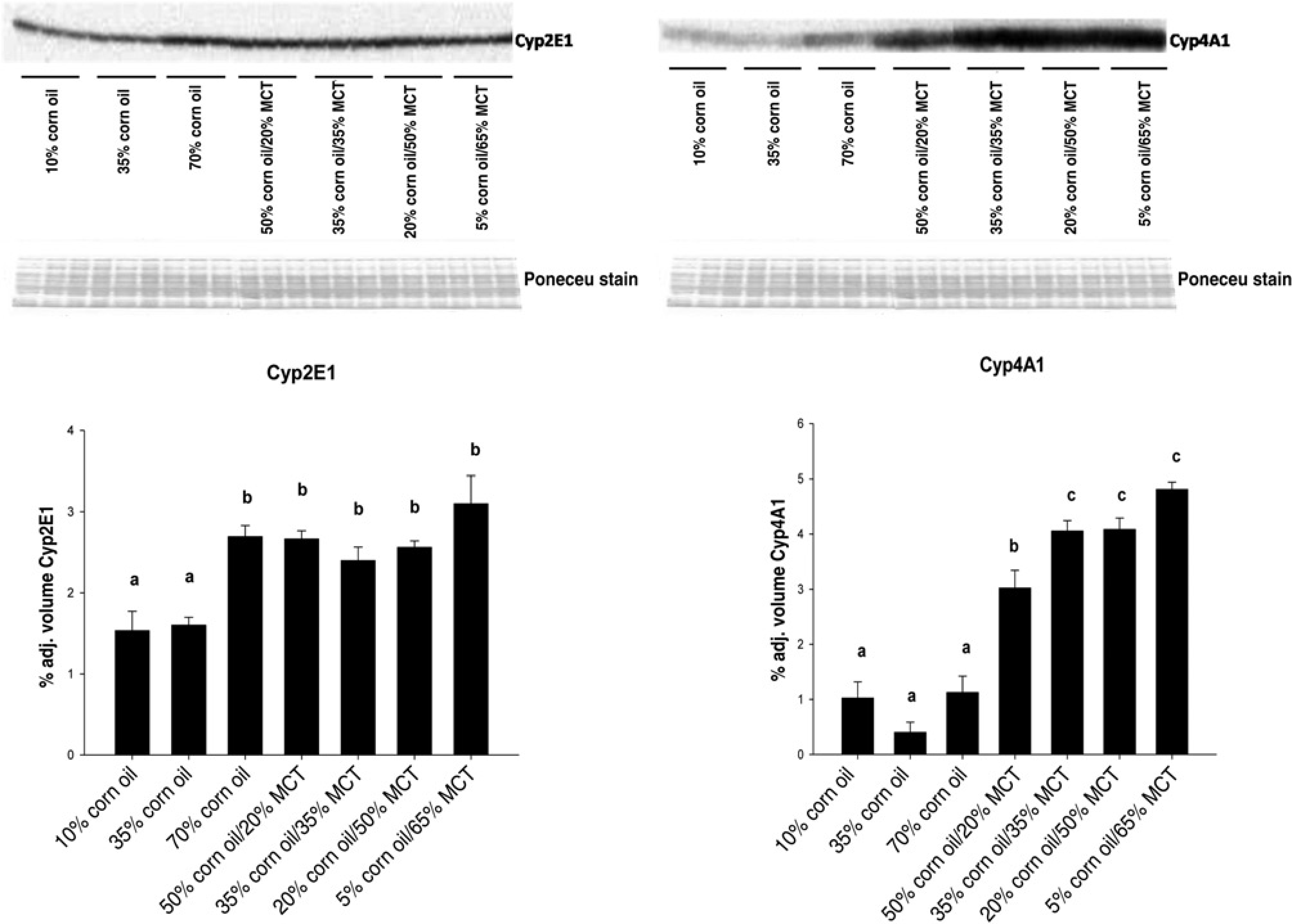
Effects of increasing dietary corn oil (CO) content and substitution of medium chain triacylglycerol (MCT) for CO on expression of CYP2E1 and CYP4A1 apoproteins in hepatic microsomes from rats overfed via total enteral nutrition. Immunoquantitation data are mean ±
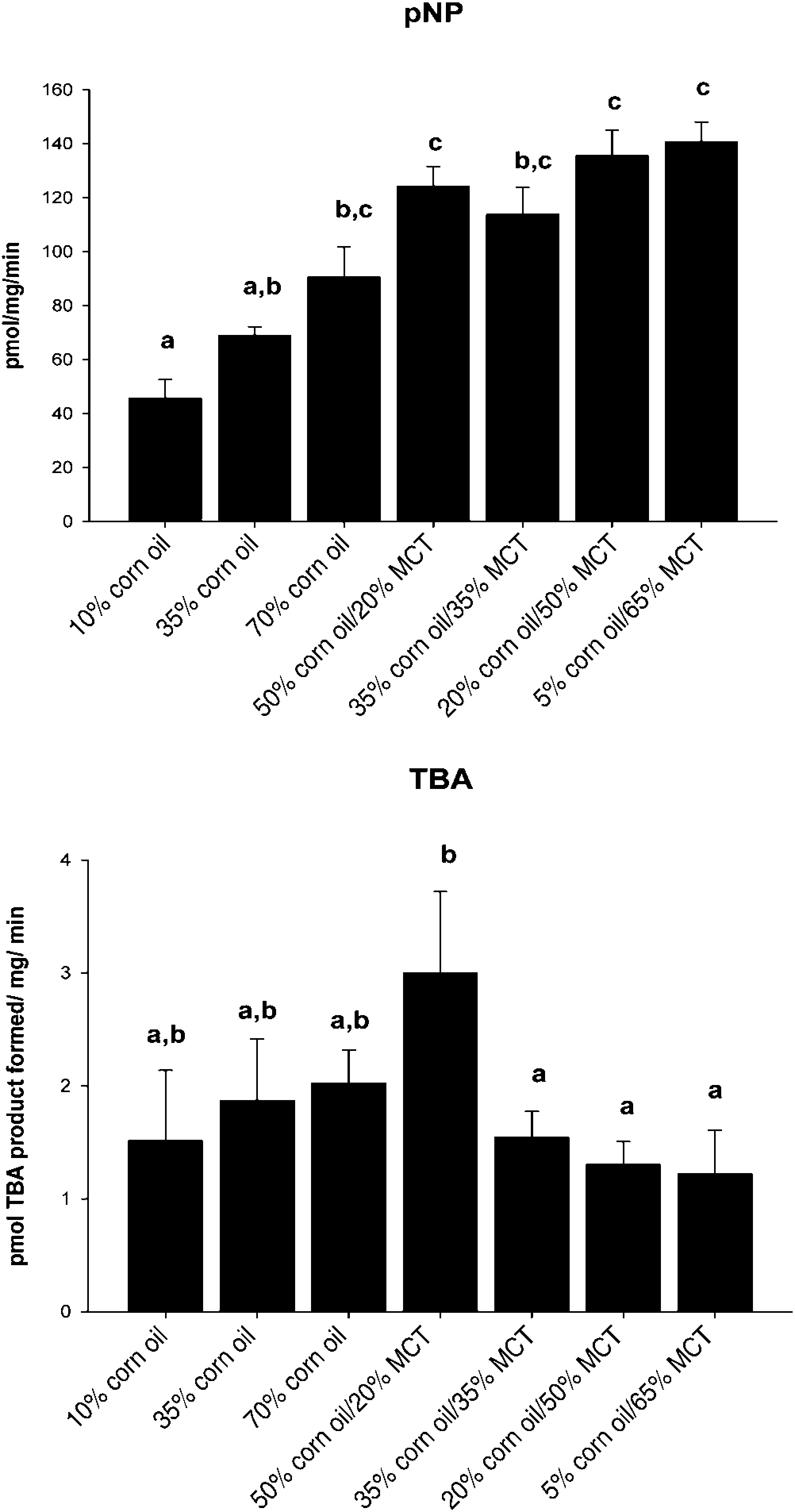
Effects of increasing dietary corn oil (CO) content and substitution of medium chain triacylglycerol (MCT) for CO on CYP2E1-dependent micro-somal monooxygenase activity. Top panel: p-nitrophenol (pNP) hydroxylase activity; bottom panel: carbon tetrachloride-dependent lipid peroxidation as measured by generation of thiobarbituric acid reactive products (TBARS). Data are mean ±
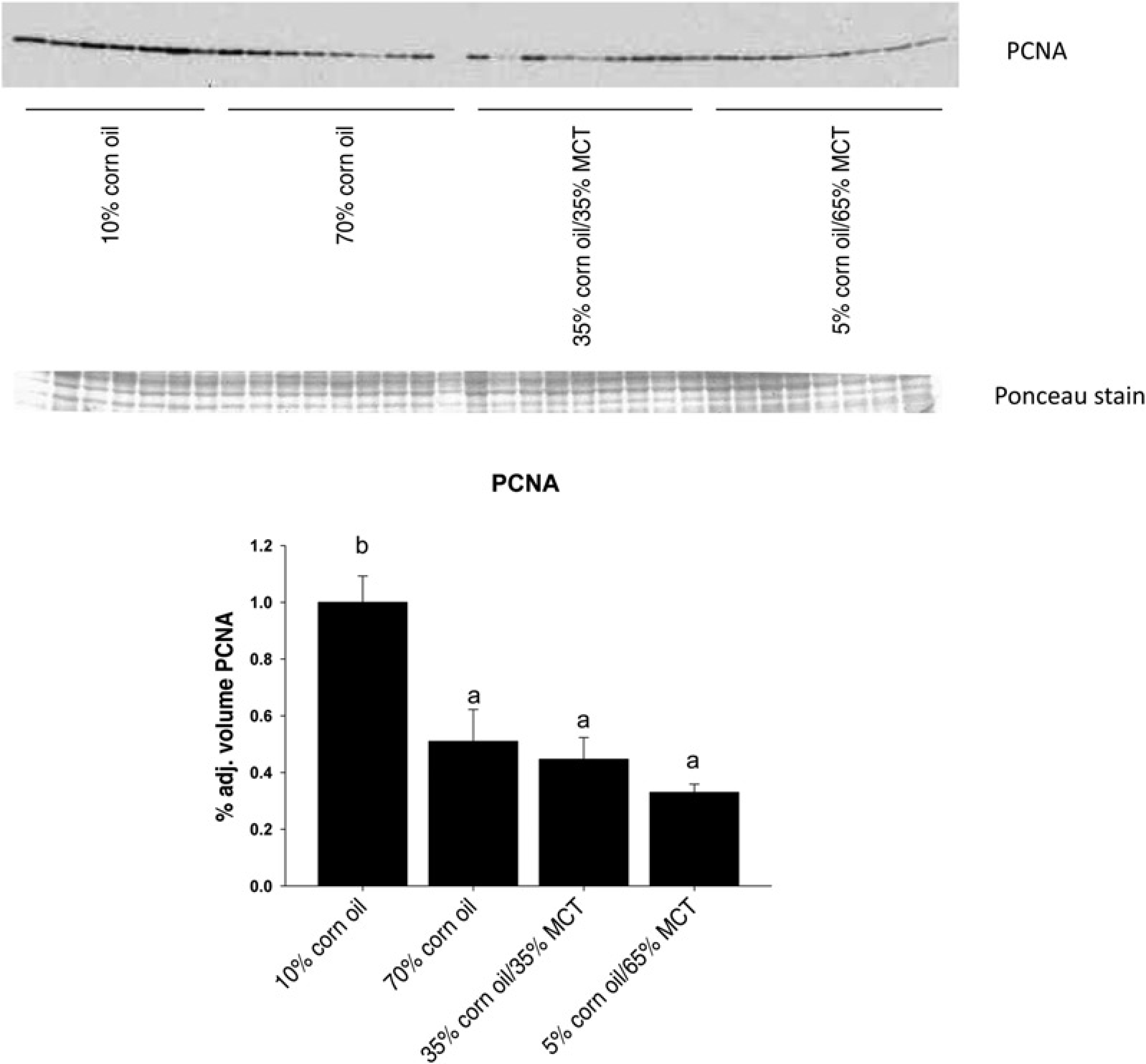
Effects of increasing dietary corn oil (CO) content and substitution of medium chain triacylglycerol (MCT) for CO on expression of proliferating cell nuclear antigen protein in liver homogenates from rats overfed via total enteral nutrition. Immunoquantitation data are mean ±
Effects of dietary fat composition on liver pathology
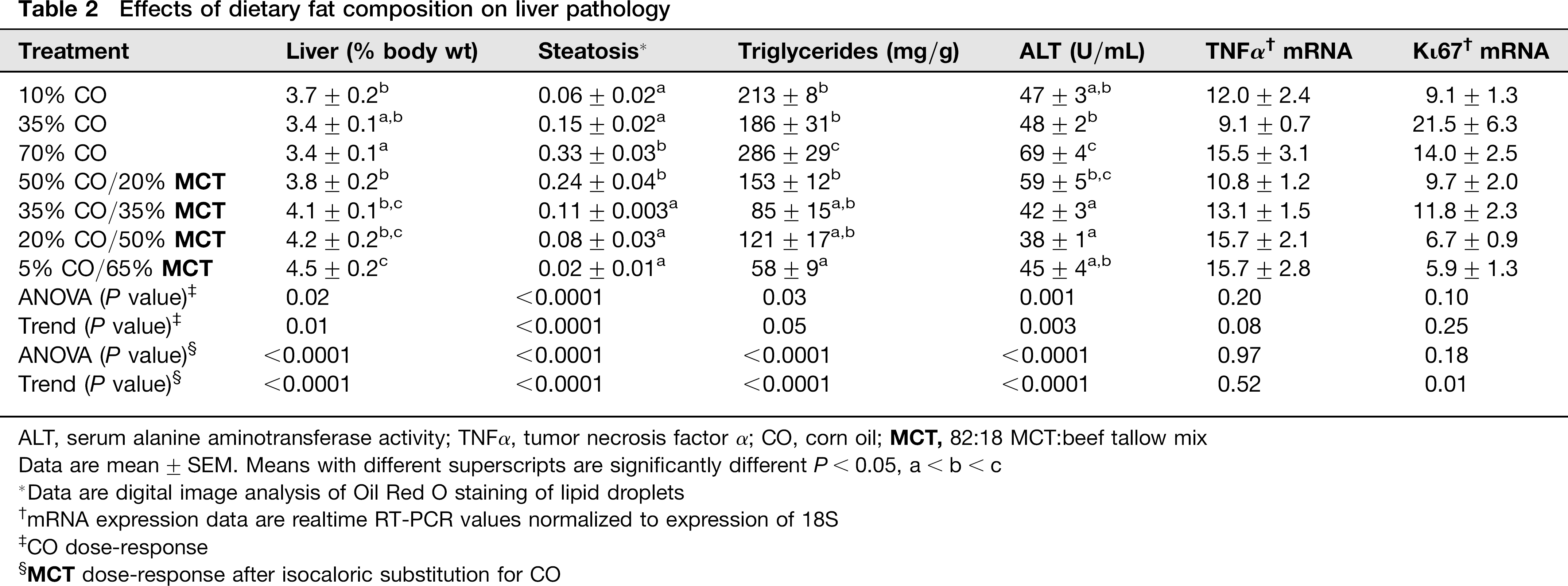
ALT, serum alanine aminotransferase activity; TNFα, tumor necrosis factor α; CO, corn oil;
Data are mean ± SEM. Means with different superscripts are significantly different P< 0.05, a < b < c
Data are digital image analysis of Oil Red O staining of lipid droplets
mRNA expression data are realtime RT-PCR values normalized to expression of 18S
CO dose-response
Effects of dietary fat composition on hepatic FA composition (μg/g liver)

CO, corn oil;
Data are mean ± SEM. Means with different superscripts are significantly different P < 0.05, a < b < c
CO dose-response
Effects of dietary fat composition on peroxidizability index of hepatic lipids *
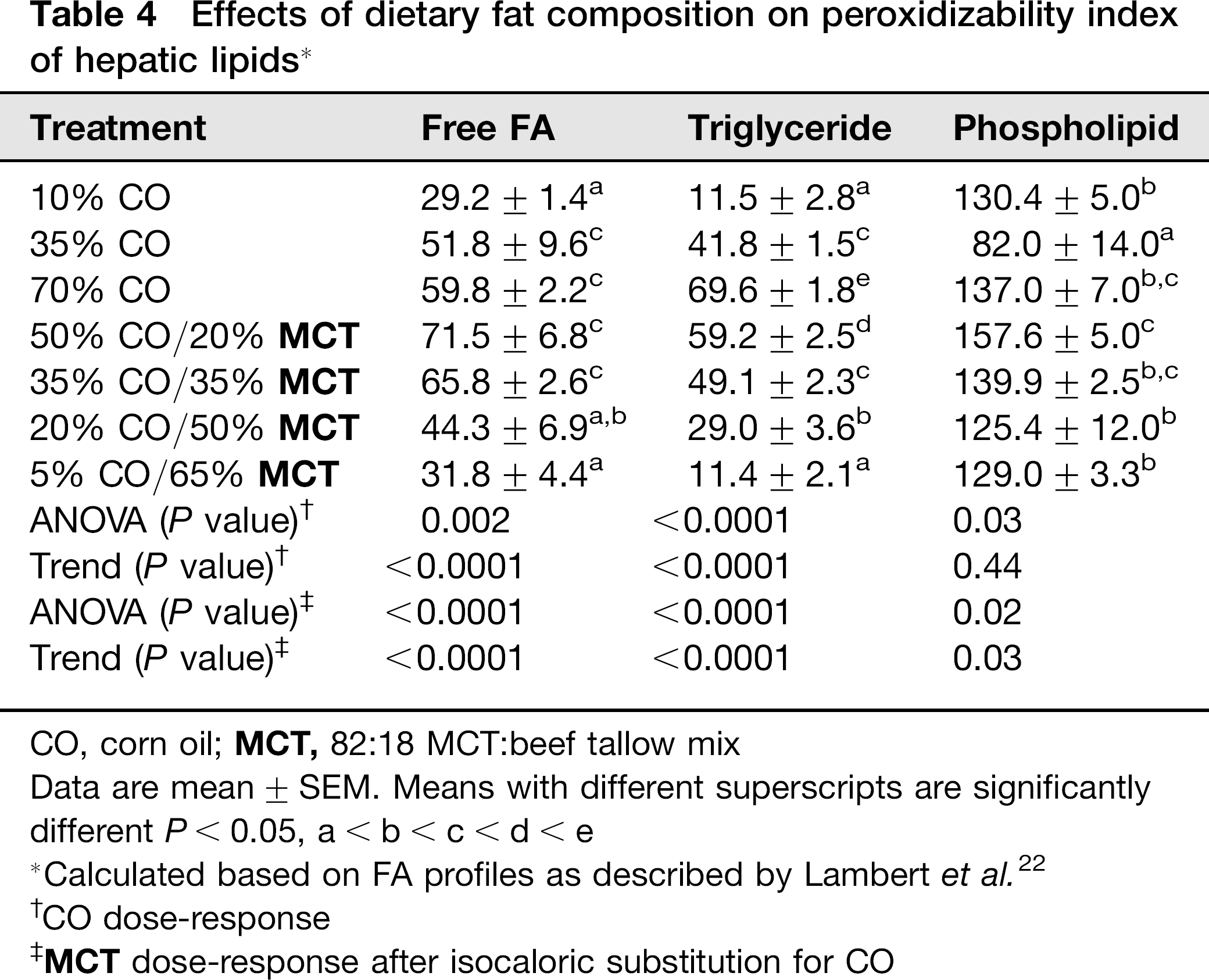
CO, corn oil;
Data are mean ± SEM. Means with different superscripts are significantly different P < 0.05, a<b<c<d<e
Calculated based on FA profiles as described by Lambert et al. 22
CO dose-response
Hepatic FA homeostasis is altered by increasing dietary CO and substitution with MCT
Overfeeding of diets with increasing %CO resulted in greater expression of mRNAs encoding the hepatic FA transporters CD36
26
and FATP2
27
(trend: P < 0.05; Table 5). In addition, as %CO increased there was a dramatic suppression in expression of adiponutrin (PNPLA3) mRNA (trend: P < 0.0001; Table 5). Adiponutrin is a hepatic triglyceride hydrolase for which inactivating mutations in humans have been linked to appearance of fatty liver disease.
28
In contrast, as hepatic steatosis appeared with increasing %CO, expression of hepatic mRNA for the transcription factor sterol regulatory element binding protein (SREBP-1c) and downstream genes encoding enzymes involved in de novo FA synthesis and desaturation
29
(fatty acid synthase [FASN], acyl CoA carboxylase [ACC1] and steroyl CoA desaturase [SCD-1]) appeared to be highly suppressed (trend: P < 0.002; Table 5). No effects of increasing %CO were observed on expression of CYP4A1 which catalyzes FA ω-oxidation
18
(Figure 2), but there was a two-fold greater expression of mRNA encoding ACO, the rate-limiting enzyme in the peroxisomal β-oxidation of FAs,
30
in the 70% CO group relative to 10% and 35% CO (trend: P < 0.01; Table 5). Although there were no significant effects on expression of apoprotein B100 (ApoB100) mRNA, increasing %CO did elevate expression of the mRNA encoding microsomal triglyceride transport protein which regulates formation of very low-density lipoprotein (VLDL) for triglyceride export out of the liver
31
(trend: P < 0.003; Table 5). Substitution of Top panel: Electrophoretic mobility shift (EMSA) assay of nuclear protein binding to the peroxisome proliferator response element element on the acyl CoA oxidase (ACO) promoter in the 70% CO and 5% CO/65% MCT groups. Competition lanes show 5% CO/65% MCT samples with or without excess cold oligonucleotide to demonstrate specificity of binding. Bottom panel: densitometric quantitation of 
Effects of dietary fat composition on expression of genes involved in hepatic lipid homeostasis *
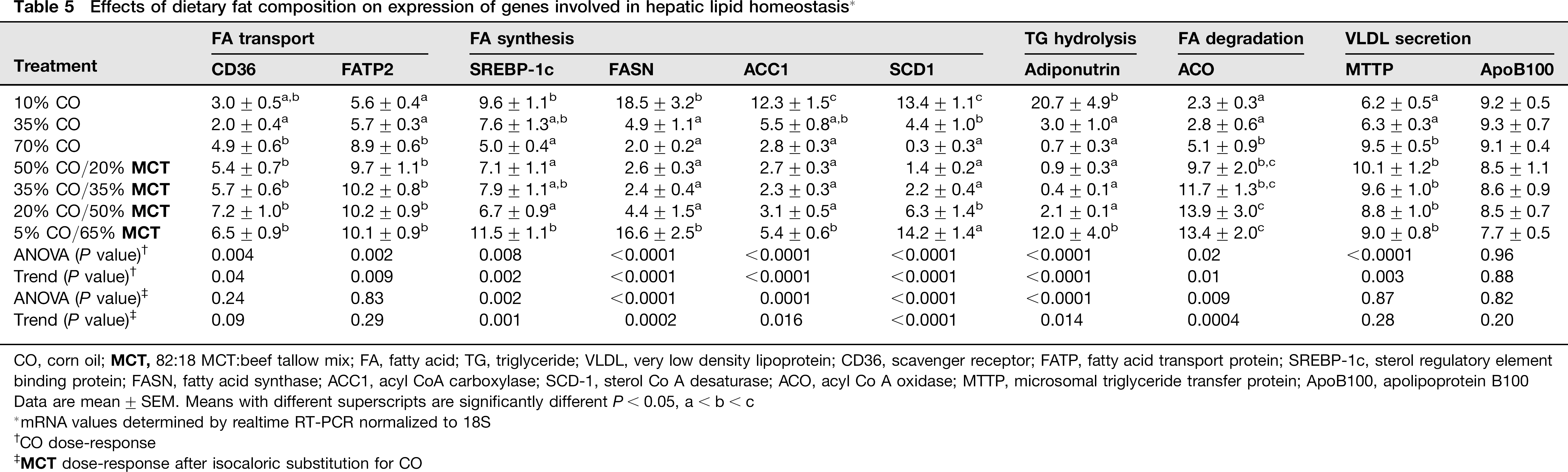
CO, corn oil;
Data are mean ± SEM. Means with different superscripts are significantly different P< 0.05, a < b < c
mRNA values determined by realtime RT-PCR normalized to 18S
CO dose-response
Effects of increasing dietary CO and substitution with MCT on mitochondrial respiration
No effects of manipulating dietary fat composition with CO or MCT were observed on expression of the mitochondrial marker mtTFAM mRNA. Moreover, no significant effects were observed on expression of UCP2 mRNA (data not shown). However, significant effects of manipulating dietary fat composition were observed on expression of proteins comprising the mitochrondrial respiratory complexes (Figures 6 and 7). Increasing the % of dietary fat as CO from 10% to 70% induced expression of Complex I, III and IV (P < 0.05), but a CO-dependent dose-response was only observed for Complex III (trend P < 0.035). Isocaloric substitution of

Representative Western immunoblots showing expression of hepatic mitochondrial respiratory complexes in rats overfed CO or CO/MCT diets via total enteral nutrition. Each lane represents a different animal (3/group). VDAC-1 expression was assessed as a loading control. CO, corn oil;
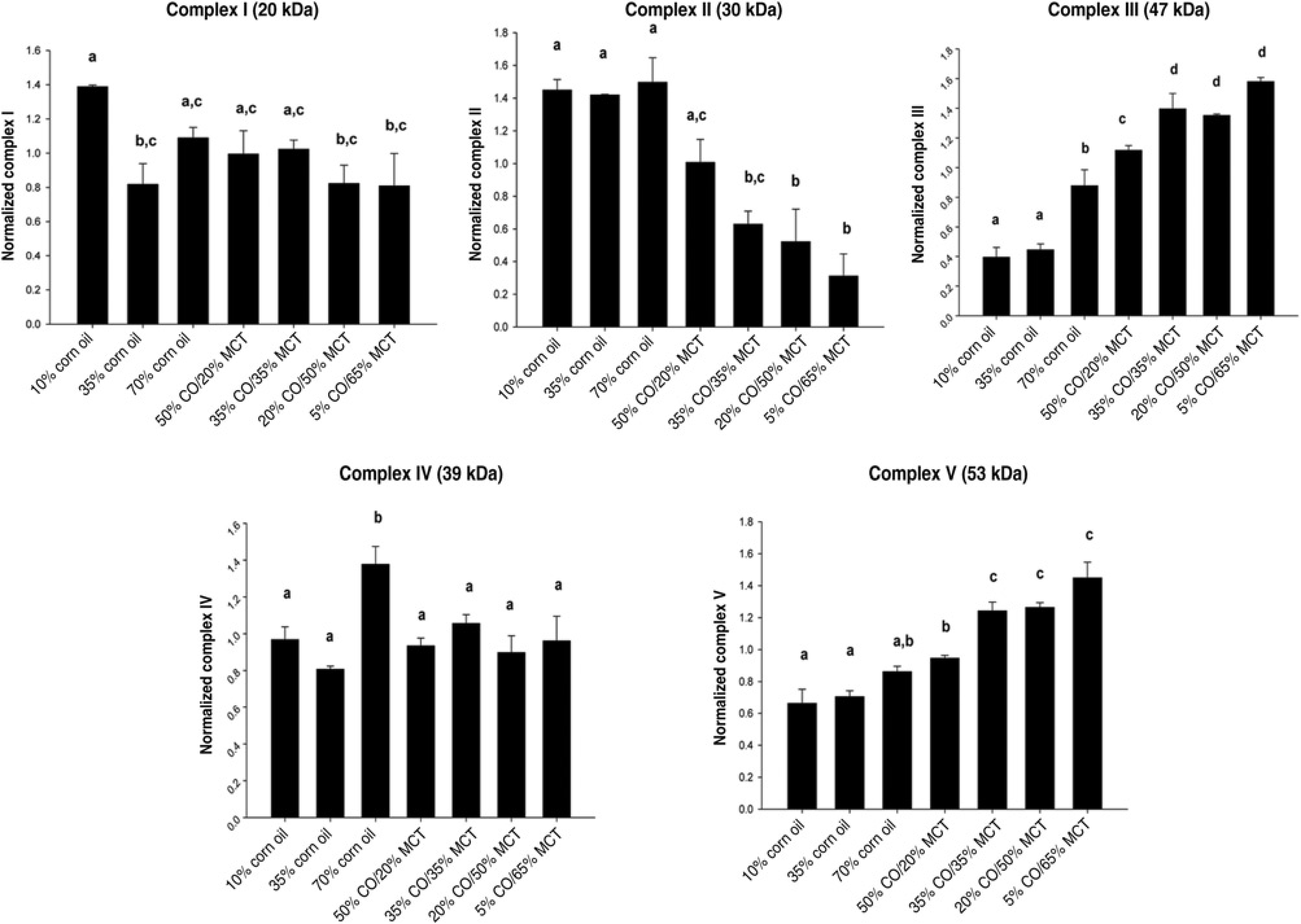
Immunoquantitation of hepatic mitochondrial respiratory complexes in rats overfed corn oil or corn oil/MCT diets via total enteral nutrition. Data are mean ±
Discussion
Although the beneficial effects of dietary medium chain fatty acids (MCFA) on energy expenditure and body composition have been examined extensively in both animal models and clinical studies,1–6 fewer studies have examined the effects of MCFA on accumulation of hepatic triglycer-ides after the feeding of high-fat diets or the effects of MCFA on the development of obesity-associated NAFLD. In the current study we observed an almost complete loss of liver pathology, including a marked dose-dependent reduction in steatosis, as the fraction of dietary fat from MCFA in the form of MCT oil was isocalorically increased. Decreases in markers of hepatic injury, such as ALTs, are most likely secondary to the decrease in steatosis. Interestingly, in contrast to the previous study of Lieber et al.,
12
we observed no evidence of increased hepatotoxicity or increased hepatocyte proliferation associated with either feeding MCT alone or as a mixture containing long-chain FAs. In contrast, increasing substitution of
Previous studies of MCFA effects on liver triglyceride accumulation in the experimental animal literature have resulted in disparate and contradictory findings. Buettner et al. 33 and Turner et al. 34 have reported significantly increased steatosis in rats and mice fed MCFA as part of pelleted high-fat diets fed ad libitum relative to animals fed low-fat diets or diets high in long-chain fats. Others, such as Baba et al., 35 reported no differences. In contrast, a recent report by Lieber et al. 12 found significantly reduced hepatic triglyceride accumulation using a liquid diet containing only MCFA compared with a liquid diet high in long-chain polyunsaturated fats. Part of the explanation for these inconsistent findings may lie in methodological differences such as use of solid versus liquid diets. Moreover, most previous studies have used ad libitum feeding where differences in food intake and caloric density result in significant differences in overall caloric intake. Additional studies in which TEN diets made with MCT are administered at different levels of calories to mimic the differences between ad libitum and isocaloric feeding would be required to address this. In addition, not all MCFA sources are the same. Both Buettner et al. and Turner et al. used hydrogen-ated coconut oil as their MCFA source and in this case the major dietary MCFAs are lauric acid (C12:0) and myristic acid (C14:0), 34 in contrast both Lieber et al. and the current study utilized MCT oil as the MCFA source. In this case the major dietary MCFAs are the shorter chain FAs C8:0 and C10:0.7,12
MCFAs are known to have different pharmacokinetics and utilize different metabolic pathways than long-chain FAs. MCFAs are not incorporated as part of chylomicrons, but instead are absorbed directly into the hepatic portal vein and have been reported to not require transporters such as CD36 or FATPs to enter the liver.2,3 Thus, in this regard, we were surprised that these transporters remained elevated in all the 70% fat diet groups compared with the 10% and 35% CO groups regardless of dietary FA composition.
The hepatic fatty acyl synthase required for triglyceride re-esterification has a chain length preference for FAs of 14 or more carbons.
5
Consistent with these data, there was little accumulation of MCFA into triglycerides or phospho-lipids in the current study. Increasing substitution of MCT for corn oil appeared to result in a substantial up-regulation of FA oxidation in the liver sufficient to significantly reduce the development of steatosis resulting from lipid oversupply. MCFA have been suggested to preferentially undergo mitochondrial and extra-mitochondrial oxidation compared with long-chain FAs.
2
Our data demonstrate a dose-dependent increase in expression of CYP 4A1 which catalyzes the microsomal ω-oxidation of FAs and in expression of mRNA encoding ACO, the rate-limiting step in peroxisomal β-oxidation of FAs when MCT was substituted isocalorically for corn oil. The increase in CYP4A1 expression is similar to that we have previously observed in alcohol-treated rats fed MCT-rich diets.
7
These enzymes are both downstream targets for the transcription factor PPARα.30,32 Our EMSA data demonstrate for the first time that dietary
The major pathway for FA oxidation is mitochondrial β-oxidation. MCFAs have been demonstrated not to require rate-limiting transformation into acylcarnitine by carnitine palmitoyl transferase for transport into the mito-chondria,
39
but instead penetrate the mitochondrial membrane directly and are rapidly oxidized to acetyl CoA. Expression of both respiratory Complex III and V (ATP synthase) were substantially elevated in response to MCT substitution for corn oil in the 70% fat fed groups, while expression of complex II appeared to be suppressed. A likely explanation for these effects is that at this high level of dietary fat, oxaloacetate levels will be limiting since it will be shuttled into gluconeogenesis to supply the brain with glucose. In the absence of sufficient oxaloacetate to condense with the acetyl group of acetyl CoA to form citrate, acetyl CoA will be used to form ketone bodies which are known to be elevated after MCFA feeding
40
rather than fed through the Krebs cycle, and less FAD2 will be oxidized through Complex II. Since expression of respiratory complex I does not change, we speculate that electrons are being delivered to Complex III directly through Complex I and that additional electrons enter the respiratory chain at Complex III from MCFA derived glycerol 3-phosphate via glycerol-3-phosphate dehydrogenase and ubiquinone.41,42 The elevation of mitochondrial respiration did not appear to be associated with uncoupling of the respiratory chain or with increased mitochondrial numbers, since we observed no increase in expression of UCP2 or mtTFAM mRNAs. Moreover, although changes in SIRT3 expression have been linked to differences in expression of mitochondrial respiratory complexes,
23
we observed no effects of
In addition to oxidation and formation of ketone bodies, published data suggest that acetyl CoA derived from MCFA may be used to synthesize longer chain FAs and esterified.2,34 Our data are consistent with this since increasing substitution of
Interestingly, our results differ substantially from those of Turner et al., 34 who reported increased mitochrondrial respiratory complexes in the muscle of male C57BL6/J mice-fed MCFA from hydrogenated coconut oil compared with mice fed low-fat diets or diets high in long-chain fats. However, they observed no effects on expression of mitochondrial respiratory complexes in the liver relative to a low-fat diet and higher expression of hepatic complexes II and IV relative to mice fed long-chain fats. In addition, this study reported increased hepatic triglyceride content in the coconut oil-fed group compared with the other groups associated with dramatic increases in expression of lipogenic enzymes and de novo lipogenesis well above levels observed in mice fed low-fat diets. It is possible that these differences in biochemical and physiological responses to feeding MCFA reflect species and strain differences. Alternatively, it is possible that the shorter chain C8:0 and C10:0 MCFAs in MCT oil have distinct and different metabolic effects than the longer chain C12 and C14 FAs in coconut oil.
In conclusion, using a rat model to study NAFLD, we have demonstrated that isocaloric replacement of dietary corn oil (long-chain polyunsaturated fats) with
Footnotes
Acknowledgements
The authors thank Mat Fergusson, Tammy Dallari and Rene Till for technical assistance. In addition, we thank Dr Gazyna Nowak, University of Arkansas for Medical Sciences for providing valuable discussion of the mito-chondrial data. This work was funded in part by ACNC-USDA-CRIS 6251-51000-005-03S.