
Abstract
The rapid evolution of HIV-1 is a major obstacle to viral eradication. Early antiretroviral therapy (ART) during primary HIV-1 infection could limit viral diversity. Eighteen patients recently infected with HIV-1 were selected. Nine initiated ART soon after enrolment and nine remained untreated. Replication-competent (RC) viruses were quantified at baseline and after one year of follow-up. Viral diversity in the C2V5 envelope region was evaluated from plasma, peripheral blood mononuclear cells (PBMCs), and cell culture at both time points. The amount of RC virus in the treated group declined (median −5.42 infectious units per million [IUPM]) while it remained stable or increased in the untreated group (median +0.87 IUPM). At one year post infection, we observed a significant increase in diversity for the C2V5 (+0.150%) region, specifically in the hypervariable loops V4 (+0.73%) and V5 (+0.77%), in the untreated group. More importantly, viral diversity did not significantly increase in treated individuals during the first year post infection. Genetic diversity during primary infection remains low through the first year of infection. Early treatment could contribute to a decrease in RC viruses from PBMCs and to limitation of viral diversification in the viral reservoir. These findings may have relevance for the rational design of specific immunotherapeutic strategies.
BACKGROUND
HIV sequence diversity is a major challenge for the design of a vaccine that could protect against a broad range of heterologous viruses. HIV-1 evolves rapidly because of its high replication rate and the low fidelity of its reverse transcriptase. Viral populations found in newly infected hosts are highly homogeneous, although continuous viral evolution generates multiple and distinct variants that remain highly related throughout the course of infection. 1–5 Several studies have linked viral diversity with disease progression and the development of AIDS. 6–9 Following acute HIV infection, the effect of treatment on viral diversity is not well documented. Early antiretroviral treatment (ART), by limiting viral replication, could attenuate viral diversity, which could protect from immune escape, chronic activation and exhaustion of the immune system. On the contrary, a low antigenic diversity could prevent the initiation of a broad immune response in the context of a more preserved immune system.
Envelope protein (ENV) is the most variable HIV-1 gene and the C2V5 region is particularly variable in sequence and length. 10,11 An earlier study has shown an increase in viral diversity of 1% per year to reach a plateau of 5% after a few years of infection. 12,13 This region is also highly glycosylated and therefore mutations in glycosylation sites can affect sensitivity to neutralizing antibodies. 14–16 We investigated the effect of early initiation of ART on the evolution of ENV in the context of the very homogeneous viral population observed during acute HIV-1 infection.
STUDY DESIGN
Study population
Eighteen subjects were selected randomly from a prospective cohort of individuals participating in the primary HIV-1 infection cohort in Montreal, Canada, based on treatment criteria. 17 They were recruited from several clinical sites and were followed for one year. Primary HIV-1 infection was first assessed by clinical and epidemiological history. Patients were enrolled in the cohort according to acute infection criteria, which consisted of a negative HIV enzyme immunoassay (EIA) serology with positive p24 antigen or a positive EIA with indeterminate Western blot and positive p24 antigen. In order to confirm acute infection, sera were tested with a detuned EIA (Abbott, Santa Laurent, QC, Canada). 18 Eight patients initiated ART in the first month following HIV-1 infection diagnosis (range from 1–25 days) and one patient initiated two months after diagnosis (57 days). Nine subjects deferred the initiation of ART during the study. The decision as to whether ART was to be initiated or not was made by the physician and the patient, based on clinical judgement and patient preference. Seven patients experienced symptoms during the acute infection phase, from both the treated and untreated groups. All patients initiating ART remained on therapy for the duration of the study. There was no treatment interruption or drug vacation for the duration of the study. Plasma HIV-1 RNA quantification was obtained by a PCR-based assay from Chiron (version 3.0), which has a lower limit of detection of 50 copies per mL.
Quantification of RC virus
Peripheral blood mononuclear cells (PBMCs) were resuspended at a concentration of 106 cells/mL in RPMI 1640 medium (Sigma, St Louis, MO, USA) supplemented with 10% heat-inactivated fetal calf serum (Sigma), penicillin (50 U/mL), streptomycin (50 mg/mL), L-glutamine (2 mmol/L), HEPES buffer (10 mmol/L) and 100 U of recombinant human interleukin-2 (Hoffmann-La Roche, Nutley, NJ, USA) per mL, and six fivefold dilutions of PBMCs were cultured (25 million to 8,000). The 25- and 5-million PBMC dilutions were cultured in T25 flasks in duplicate, and the higher dilutions were cultured in 24-well plates in quadruplicates. A CD3-CD8-bispecific monoclonal antibody (MAb), which selectively depletes CD8 cells while activating CD4 cells, was added at a final concentration of 1 mg/mL. Cell cultures were incubated at 37°C in a humidified 5% CO2 atmosphere and maintained for a 21-day period with twice-weekly medium exchanges. Supernatants were collected weekly before the medium change for the measurement of HIV-1 p24 antigen by an enzyme-linked immunosorbent assay. The number of infectious units per million (IUPM) PBMC was calculated from the pattern of positive wells by the method of maximum likelihood.
PCR amplification
Viral clones were obtained from proviral DNA from PBMCs, from plasma RNA and from replication-competent viruses recovered from cultured PBMCs. Biological clones were obtained from extraction of RNA from all positive culture wells. Proviral DNA was extracted from PBMCs and molecular clones were obtained from serial dilutions and multiple independent wells were selected for single genome sequencing. RT-PCR and nested PCR was performed covering the C2V5 region generating a 718-base pair fragment with primers described elsewhere. 19 Reverse transcription was carried out at 50°C for 30 minutes. Amplification conditions were: 94°C for five minutes followed by 35 cycles at 94°C for 30 seconds, 55°C for 30 seconds and 68°. Sequencing was performed at Génome Québec sequencing facilities on a 3730 × l DNA analyser system from Applied Biosystems (McGill University and Genome Quebec Innovation Centre, Montreal, Quebec, Canada). All nucleotide sequences were submitted to GenBank and provided with accession numbers GQ507803–GQ508254.
Phylogenetic analysis and statistical analysis
Sequences were aligned using Clustal W version 1.83 20 followed by manual alignment editing using BioEdit version 7.0.4.1. Pairwise nucleotide distances within each patient were estimated with a Kimura two-parameter model using a 2.0 transition/transversion ratio as implemented in the Mega 3.1 software 21 with a 1000-bootstrap value. We used the non-parametric Mann–Whitney method to compare sequence evolution within all patients at baseline and year one within untreated and treated groups. We compared the diversity for each group at each time point with the Wilcoxon test. Pearson's χ 2 and/or correlation coefficient test were used to test the relationships between variables. All analyses were performed using SPSS version 15 (SPSS Inc., Chicago, IL, USA) with a significance level of 5%.
RESULTS
Patient characteristics
Both groups shared similar characteristics: in the treated group, 78% were men who have sex with men (MSM), median age was 36.0 ± 8.3 years and 78% experienced symptomatic acute infection. For the untreated group, 89% were men, 67% acquired HIV from MSM contact, median age was 43.1 ± 7.77 years and 78% had symptoms at onset. There was no difference between groups according to median CD4 count and viral load at baseline. After one year, the median increase in CD4 count was +180 cells/mm3 in the treated group, while it decreased or remained stable in the untreated group (−130 cells/mm3). All treated patients achieved undetectable viral loads within the first year of infection while the median viral load was 26,237 copies/mL in the untreated group.
Viral quantification
Using an ultrasensitive culture technique, virus was recovered from patients at baseline and follow-up. There was no significant difference in IUPM PBMCs for both groups at baseline (P = 0.436). We observed a decline in the IUPM in 8/9 treated patients and in 3/9 untreated patients (P = 0.016). The magnitude of the decline in the treated group was of −5.42 IUPM and an increase of +0.87 was observed for the untreated group (P = 0.001). IUPM decline was not correlated with viral load at baseline (P = 0.830) (Figure 1).
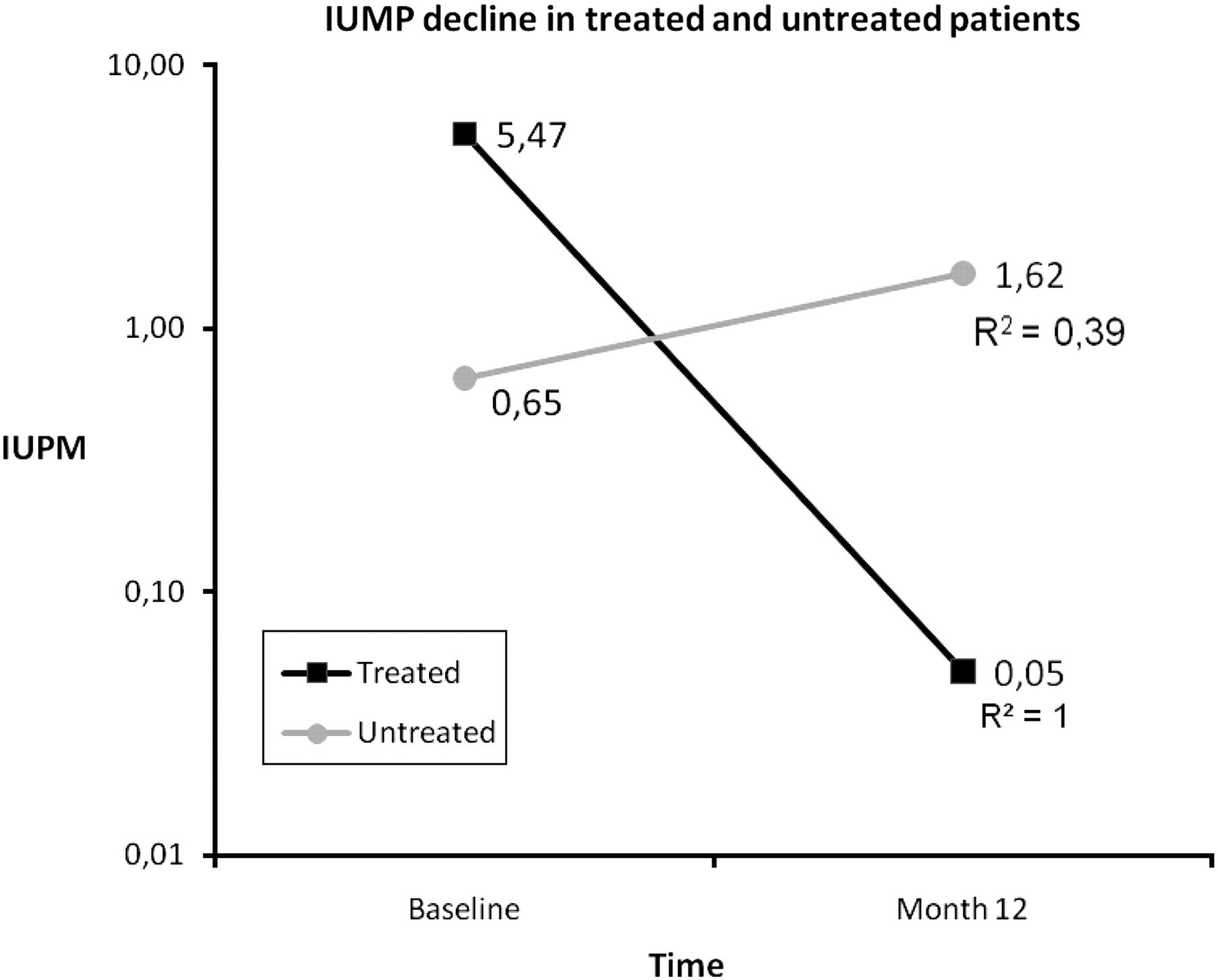
Decline in infectious units per million (IUPM) of peripheral blood mononuclear cells in treated and untreated patients during the first year post infection
Evolution of HIV-1 envelope C2V5 is limited in the year following acute infection
All HIV-1 isolates were subtype B. Highly specific clustering of autologous sequences from patients excluded possible cross-contamination among subjects or with laboratory strains. We measured intrapatient sequence diversity for all patients at the time closest to infection before ART initiation except for one patient. The median diversity at baseline was 0.19% and was not statistically different between patients who initiated ART compared with the patients who remained untreated (0.09% versus 0.23%, respectively, P = 0.93). There was no correlation between viral diversity and CD4 count (P = 0.64), viral load (P = 0.704), or IUPM (P = 0.830). During the first year of infection, sequence diversity in the entire C2V5 region increased +0.06% in the treated group (P = 0.678) and +0.15% for the untreated group (P = 0.028). The difference between the two groups was not statistically significant (P = 0.297) (Figure 2).
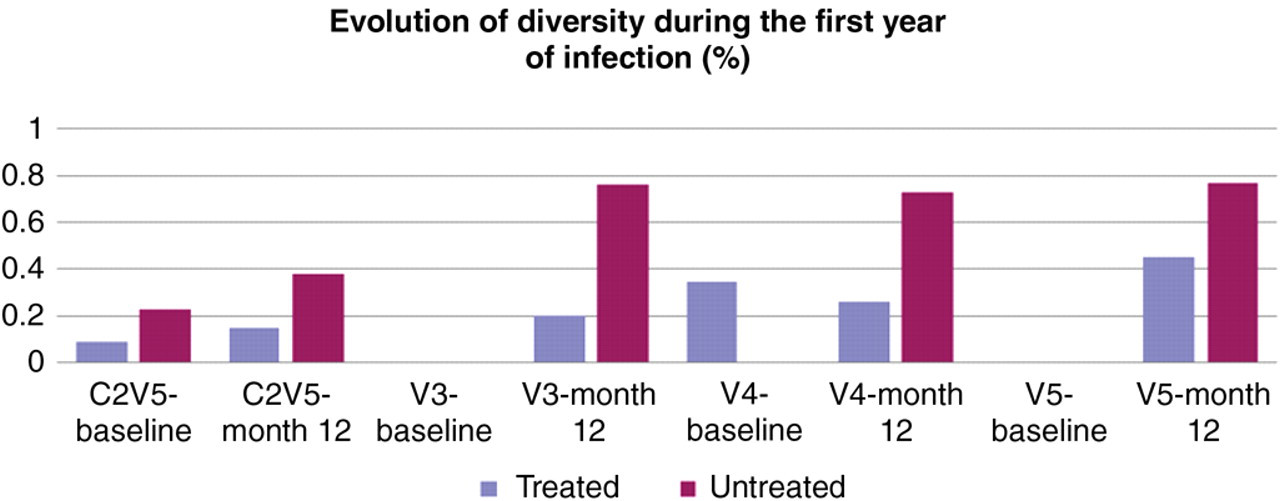
Evolution of diversity (%) for all patients in the hypervariable loops V3, V4, V5 and C2V5 at the time closest to infection (baseline) and after 12 months of follow-up (M12)
We also looked at changes in the potential N-glycosylation (PNG) sites during the first year of infection. The number of PNG sites increased from 11.0 to 12.0 in treated patients (P = 0.66) and from 11.4 to 11.90 in the untreated (P = 0.51), which was not significant.
Characterization of hypervariable loops V3, V4 and V5
Clonal analysis of the V3 loop revealed that all subjects harboured viruses with a single V3 loop length, most of them consisting of 35 amino acids, while three patients had 34 amino acid V3 loops. Two of these three subjects were in a transmission pair. Viral populations were extremely homogeneous in V3 at baseline for both groups and increased during our study to reach 0.20 for the treated patients and 0.76 for the untreated group. Viral diversity increased for both groups (+0.75%, P = 0.327) compared with the treated group (+0.20%, P = 0.674); however, those differences were not significant.
Sequence patterns for V4 were more complex. The lengths of V4 loops ranged from 28 to 38 amino acids, and four subjects had viral populations harbouring two different loop lengths, while one subject had three. Viral diversity for the entire population was 0.08% at baseline and was not significantly different between the two groups (P = 0.436). It increased to 0.49% after one year. However, in the group initiating ART, diversity decreased during the course of infection to reach 0.26% one year later, while in the untreated group diversity increased +0.73% (P = 0.046). Although there was a greater increase in viral evolution for the untreated group, the differences in rate of sequence evolution did not reach statistical significance between the two groups (P = 0.094).
All patients except one had a unique V5 length (11–16 amino acids). Baseline diversity was extremely low for the entire population. Overall, viral diversity in V5 did not significantly increase during the first year of infection (+0.56% P = 0.340). In the treated group, diversity increased from to 0.45% (P = 1.0) while viral diversity significantly increased to 0.77% in the untreated group (P = 0.046). The difference was not statistically different between the two groups (P = 0.34) (Figure 2).
DISCUSSION
HIV-1 diversification has been associated with an attempt of the virus to escape control by the immune system. However, some studies have linked high level of sequence diversity with slower disease progression and better immune responses. 22–26 Characterizing viral diversity during early infection is important in order to identify strains that are more likely to establish infection. Conserved epitopes within these ancestral strains could be targeted by vaccines to elicit strong cellular or humoral immune responses against viruses with the highest potential to initiate infection. In this study, we evaluated the effects of early ART initiated during acute HIV-1 infection on viral suppression and evolution, exploring the hypothesis that suppression of viral replication by ART might limit viral diversification and its potential to escape the immune system. Our results suggest that early ART does play a role in viral reservoir decay and may limit or slow viral diversification.
As expected, ART led to a rapid decline in viral load and recovery in CD4 cell counts. The level of PBMC IUPM was significantly lower in the treated group and no decline in IUPM was observed in the untreated group. After one year of suppressive therapy, RC viruses isolated from PBMCs represented predominantly virus from latent reservoirs. Most studies showing the stability of viral reservoirs over time have been performed in chronically infected individuals. Our subjects started therapy during the acute infection stage and therefore early treatment may have helped limit the replenishment of these reservoirs.
In order to study viral evolution, we chose to study the ENV gene. ENV encodes the surface glycoproteins gp120 and gp41 and exhibits the highest degree of evolution, particularly in C2V5. 2,3,12 This region is highly glycosylated, and changes have been associated with escape from neutralizing antibodies. 16 Mutations in ENV have also been shown to affect T lymphocyte cytotoxic epitopes. 27,28 In our study, viral diversity was low overall in agreement with recent studies showing that very few viral variants are transmitted to a new host, and infection is established from these initial variants. 12,29–31 At the end of study, there was a trend towards higher diversity in the untreated group compared with the treated group, albeit not statistically significant, suggesting that early treatment may limit viral evolution. The small sample size and the length of follow-up may have limited our ability to show statistical significance. Viral evolution in treated patients suggests that residual replication occurs in such individuals. C2V5 diversity did not correlate with viral load, which was consistent with a previous study, 32 although another study with subtype CRF02_AG infection did show a correlation. 33 There was no association between CD4 count and viral diversity, either at baseline or one year post infection, in contrast to a study by Delwart et al. 23 that showed reduced diversity in patients with rapid CD4 decline.
Several studies have shown that hypervariable loops V3, V4 and V5 accumulate mutations at glycosylation sites and that the number of PNG sites may vary during the course of infection. 15,16,34 This was not the case for our patients during the first year post infection. Although we observed an increase in PNG sites for both groups, our results did not reach statistical significance. Nevertheless, different patterns of PNG were observed, which could affect neutralization sensitivity. 35
Our results suggest that treatment may play a role in limiting C2V5 sequence diversity by either diminishing the level of viral replication, thus decreasing the virus's reverse transcriptase opportunity to generate new variants, and/or by diminishing viral antigen exposure to the immune system, limiting pressure on the virus to generate better-adapted quasispecies. Low viral diversity could be more amenable to viral control with immunotherapeutic strategies. Although our study was limited in size and length of follow-up, our results support further studies on early treatment during acute infection to evaluate whether clinical benefits can be derived from such a strategy.
Footnotes
ACKNOWLEDGEMENTS
We wish to thank all the patients, the study coordinator Mario Legault, our Network of Clinical Investigators and their research team: R G Lalonde, J Allan, N Gilmore, M Klein, J MacLeod, M Potter, C Tsoukas, J Falutz, J Cox, from the McGill University Health Center (MUHC), M A Charron, C Fortin, A de Pokomandy, A Talbot, for Centre Hospitalier de l'Université de Montréal (CHUM), F Asselin, M Boissonneault, L Charest, H Dion, Emmanuelle Huchet, S Lavoie, D Legault, D Longpré, P J Maziade, M E Morin, D Murphy, V K Nguyen, R O'Brien, V D Pham, S Vézina for Clinique médicale l'Actuel, M A Charron, S Dufresne, P Junod, M S Joyal, B Lessard, F Laplante, Y Parent, D Poirier, M Potter, A Talbot, D Tessier, È Sasseville for Clinique médicale du Quartier Latin and Roger LeBlanc for project LORI at Clinique médicale Goldberg, Leblanc, Rosengren, MacLeod, Gilmore. We also want to thank Genevieve Bélanger-Jasmin for her administrative assistance, Miguel Chagnon for help with statistical analysis and Dr Johnson Wong for providing the CD3, CD8 antibodies and medical lab technologist for blood processing and freezing at the network main repository. CT and JPR are clinical investigators receiving financial research support from the Fonds de la recherche en santé du Québec (FRSQ). This scientific work was supported financially by a research grant from the Canadian Institutes of Health Research (CIHR) HSH6388, HOP64507 and by the Réseau sida et maladies infectieuses (Réseau SidaMI) du Fonds de la recherche en santé du Québec (FRSQ). The authors declare no conflict of interests.