
Abstract
Summary
Unintentional infection of laboratory rodents can compromise scientific research as well as the health of the animals and animal handlers. The source of contamination often is unknown, but may be introduced by wild rats from surrounding environments. To determine whether rats in Baltimore, Maryland, USA carry infectious agents commonly found in laboratory rodent colonies, we live-trapped 162 rats during 2005 to 2006 and screened them for a panel of viruses, bacteria and parasites. Antibodies against rat coronavirus/sialodacryoadenitis virus (91.7%), Mycoplasma pulmonis (72.9%), cilia-associated respiratory bacillus (52.1%), rat parvovirus/rat minute virus (29.2%), Kilham rat virus (10.4%), Toolan's H-1 virus (10.4%), Sendai virus (4.2%) and Theiler's mouse encephalomyelitis virus (4.2%), were detected in wild-caught Norway rats. Antibodies against reovirus and pneumonia virus of mice were not detected in wild Norway rats. Endoparasites, including Nippostrongylus braziliensis (71.6%), Rodentolepis nana or Hymenolepis diminuta (34.4%), Hetarakis spumosa (24.1%) and Trichuris muris (14.8%), as well as ectoparasites (14.8%), were identified in wild-caught rats. The risk of pathogen transmission from wild-caught rats to laboratory colonies needs to be mitigated by minimizing exposures rather than assuming wild animals represent a minimal hazard.
Unintentional introduction of infectious agents into laboratory rodent colonies may cause serious complications in biomedical research (Baker 1998). In addition to compromising data recovered from infected rodents, the health of laboratory colonies and animal handlers may be at risk during an outbreak. Infection of laboratory rodent colonies occurs worldwide and the mode of pathogen introduction often is unknown (Baker 1998, Jacoby & Lindsey 1998, Zenner & Regnault 2000, Livingston & Riley 2003, Gaertner 2004, Clifford & Cosentino 2006, Schoondermark-van de Ven et al. 2006). To determine whether wild rats may be implicated in outbreaks in laboratory rodent colonies, we screened wild-caught Norway rats (Rattus norvegicus) in Baltimore, Maryland, USA for several rodent-borne pathogens, including rat coronavirus/sialodacryoadenitis virus (RCV/SDA), rat parvoviruses (either rat parvovirus [RPV] or rat minute virus [RMV]), Kilham rat virus (KRV), Toolan's H-1 virus (H-l), Sendai virus, Theiler's mouse encephalomyelitis virus (TMEV), reovirus, pneumonia virus of mice (PVM), Mycoplasma pulmonis, cilia-associated respiratory bacillus (CARB), Nippostrongylus braziliensis, Rodentolepis nana or Hymenolepis diminuta, Heterakis spumosa and Trichuris muris, pinworms (i.e. Syphacia muris) and ectoparasites.
Methods
Wild-caught rats
Adult male and female R. norvegicus were live-trapped (Tomahawk Trap Co, Tomahawk, WI, USA) in East Baltimore, Maryland, USA. Rats were trapped from July 2005 to April 2006. All trapping locations were in urban areas in alleys behind residential dwellings. Traps were baited with peanut butter and set at locations approximately 1–2 h before sundown. Details of sampling procedures have been described previously (Glass et al. 1988). Rats were collected and processed the next morning. Rats were euthanized using CO2, weighed, sexed and bled by cardiac puncture. Serum was stored at –80°C until serological analysis. Each rat was examined for ectoparasites using a fine comb and ectoparasites were speciated visually using light microscopy (100× final magnification). Faecal and caecum content samples were collected for helminth ova analysis. The Johns Hopkins Animal Care and Use Committee (protocol no. RA05H6) approved all procedures described in this study.
Serological analyses
Serum samples were submitted to a commercial laboratory (BioReliance Invitrogen Bioservices, Rockville, MD, USA) for CARB, H-1, KRV, M. pulmonis, PVM, RCV/SVA, reovirus, RPV, Sendai virus and TMEV serology using the methods outlined in Table 1. For RPVs, samples that were positive for RPV by immunofluorescent assay (IFA) and negative for H-1 and KRV by haemagglutination inhibition test (HAI) were considered serologically positive for RPV/RMV by default; these samples, however, could represent a yet undescribed parvovirus. The prevalence for RPV/RMV may be underestimated because serum that was positive for H-1 or KRV could also be positive for RPV/RMV, but the diagnostic test used could not distinguish it. A TMEV-like virus has recently been reported in rats, and due to cross-reactivity with murine TMEV (i.e. GDVII), serology results for TMEV have included samples that may be positive for TMEV or rat TMEV-like virus (Ohsawa et al. 2003).
Serological screening programme (Bioreliance, Inc)
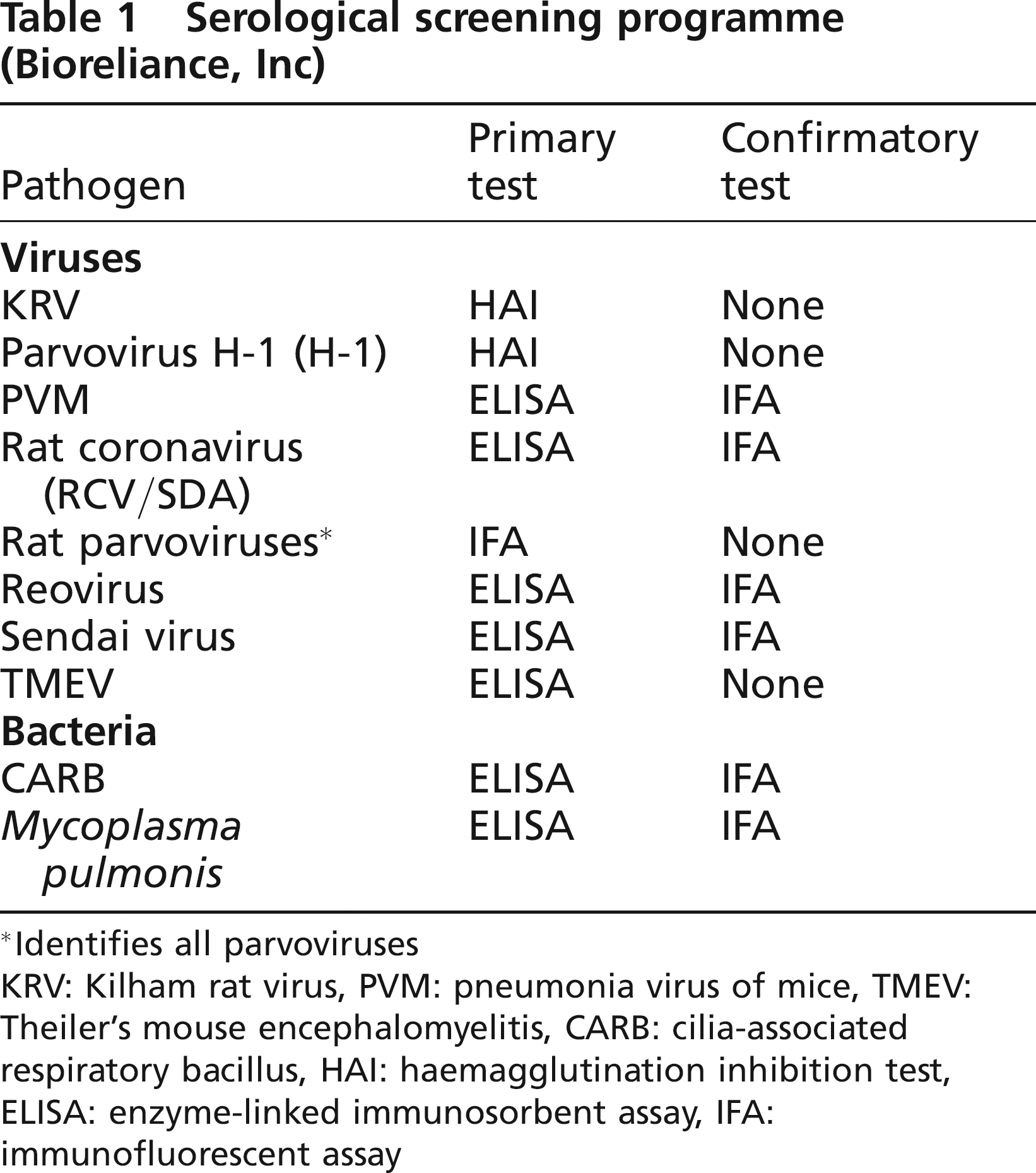
Identifies all parvoviruses
KRV: Kilham rat virus, PVM: pneumonia virus of mice, TMEV: Theiler's mouse encephalomyelitis, CARB: cilia-associated respiratory bacillus, HAI: haemagglutination inhibition test, ELISA: enzyme-linked immunosorbent assay, IFA: immunofluorescent assay
Helminth identification
Helminth ova were identified following faecal and caecum content floats. Briefly, faecal and caecum content samples were homogenized in zinc sulphate buffer (400 g/L) in glass test tubes and filled to the brim with buffer. A coverslip was placed on top for 15 min and transferred to a slide for microscopic evaluation (400 × final magnification). Clear tape was applied to the anus, transferred to a glass slide and evaluated under the microscope for pinworm (Syphacia sp.) eggs (400× final magnification) (Flynn 1973, Kohn & Barthold 1984).
Statistics
Differences in pathogen prevalence by age and seasonality were evaluated by χ2-tests. Body mass was used as a correlate of age as follows: juveniles were <200 g (n = 31), young adults were 200–399 g (n = 71) and adults were >400 g (n = 98). Rats over 200 g were sexually mature as indicated by the descent of testes in males and development of vaginal openings in females. Seasons were defined as: winter (December–February), spring (March–May), summer (June–August) and fall (September–November). Relationships between pathogens were analysed using Pearson product moment correlation analyses. Comparisons were considered statistically significant at P < 0.05.
Results
Prevalence of rodent-borne pathogens
A total of 162 wild rats were trapped as part of the study. The panel of pathogens for this study included the panel used for sentinel rats at the Johns Hopkins University Medical Institutions (JHMI) in Baltimore, Maryland. Pathogens that could be visually identified (i.e. nematodes and ectoparasites) also were included. For those pathogens that were screened for the sentinel panel of pathogens, cost constraints limited the numbers of individuals that could be tested; for those agents a subsample of individuals (n = 48) was selected to be equally distributed among seasons. The prevalence of antibody or rodent-borne pathogen for the agents tested was generally high – indicating these agents commonly circulate in wild populations (Table 2). Antibody against RCV/SDA was detected in almost all rats tested (91.7%; 44/48). Nearly three-quarters of the rats were infected with rat hookworm, N. braziliensis (71.6%; 116/162), and had antibody against M. pulmonis (72.9%; 35/48). More than one-quarter of the rats tested had detectable antibody against CARB (52.1%; 25/48) and RPV/RMV (29.2%; 14/48). Common endoparasites, including R. nana or H. diminuta (34.4%; 55/162), H. spumosa (24.1%; 39/162) and T. muris (14.8%; 24/162), were present in more than 10% of the rats. Antibodies against KRV (10.4%; 5/48), H-1 (10.4%; 5/48), Sendai virus (4.2%; 2/48) and TMEV (4.2%; 2/48) were detected less commonly in rats. None of the Norway rats had pinworms (S. muris; 0/162) or detectable antibody against PVM or reovirus (0/48 for both). Ectoparasites were collected from 14.8% (24/162) of the rats; the spiny rat mite (Laelaps echidninus) was the most prevalent ectoparasite and two cat fleas (Ctenocephalides felis) and three tropical rat mites (Ornithonyssus bacoti) also were collected.
Prevalence of pathogens in wild-caught Norway rats

IFA positive for RPV/RMV and HAI negative for KRV and H-1
Crossreacts with a rat TMEV-like virus
See Table 1 for abbreviations
Age
The distribution of rats by age was variable for the helminth panel (juveniles, n = 20; young adults, n = 56; and adults, n = 86) and the serological screening panel (juveniles, n = 1; young adults, n = 15; and adults, n = 32). Of all pathogens included in this study, only the prevalence of N. braziliensis increased with age (χ2 = 8.27, P = 0.016).
Season
Rat trapping success was not evenly distributed across seasons due to a rainy fall in 2005 (spring, n = 46; summer, n = 33; fall, n = 20; and winter, n = 63). Samples selected for serological testing were equally distributed across seasons (n = 12/season). Seasonal differences were observed for R. nana or H. diminuta, in which the prevalence was significantly lower in the spring (17.4%) as compared with summer (45.5%), fall (40.0%) and winter (38.1%) (χ2 = 8.38, 3; P = 0.04). The prevalence of N. braziliensis infection was significantly higher in the fall (100%) as compared with spring (57.7%), summer (65.6%) and winter (74.2%) (χ2 = 12.76, P = 0.005). The prevalence of ectoparasites was significantly higher in the summer and fall (31.2% and 31.6%, respectively) as compared with the winter and spring (4.7% and 4.4%, respectively; χ2 = 21.86, 3; P < 0.001).
Correlations
There was a correlation between infection with M. pulmonis and CARB (r = 0.542, P < 0.0001). Of the two samples that were positive for Sendai virus, one of those was also positive for TMEV. No correlations existed among prevalences of all other pathogens tested.
Discussion
Historically, pathogens reported in laboratory rodent colonies reflected those found in the wild because rodents were housed in open cages in large rooms with no barriers against the spread of infection between laboratory rodents or from wild rodents entering the facility (Lindsey & Baker 2006). Initial attempts were made to eliminate pathogens that caused apparent morbidity or mortality in rodents. Due to technical advances, several non-pathogenic infections were discovered in laboratory rodent colonies (Baker 1998). Although not pathogenic to rodents, infections were reported to alter research data (e.g. cellular and humoral immune responses); thus, modern research facilities have much lower prevalences of pathogens due to the availability of pathogen-free rodents from vendors, improved infection control procedures in research institutions, and routine use of surveillance programmes to diagnose colony infections (Compton et al. 2004).
As a consequence, recent reports indicate a major reduction in pathogens in laboratory colonies (Lindsey & Baker 2006). The most commonly reported rat-borne pathogens detected in laboratory rat colonies in the United States remain pinworms (i.e. Syphacia sp.), parvoviruses (RPV, RMV, H-1 and KRV), TMEV, coronaviruses and mesostigmatid mites (e.g. L. echidninus and O. bacoti) (Jacoby & Lindsey 1998, Livingston & Riley 2003, Chu & Couro 2005, Cole et al. 2005, Coons 2005, Clifford & Cosentino 2006). These organisms are difficult to eradicate because they survive in the environment for long periods (e.g. parvoviruses, pinworm eggs and TMEV), are highly contagious (e.g. coronaviruses) or are not well contained (e.g. pinworm eggs are resistant to all commonly-used disinfectants and mesostigmatid mites penetrate barrier caging and travel long distances to find new hosts) (Gaertner 2004).
Although outbreaks may be primarily caused by introduction of an infected laboratory animal or animal product (i.e. transplantable tumour or serum), the source of contamination often is unknown. Wild rodents may contribute to laboratory outbreaks and have been implicated in prior outbreaks of infectious agents in laboratory colonies (Skinner et al. 1977, Barthold 1998). In many urban areas, wild rats live in close proximity to universities and research centres, may frequent laboratory animal receiving areas and may even enter laboratory housing areas (e.g. during construction projects or in delivery trucks). Although wild rats are less likely to reside in laboratory buildings as compared with mice, they may transmit pathogens via fomites, resident commensal mice, aerosols or direct migration of parasites (Sullivan 2004).
The JHMI in Baltimore, Maryland are located in an urban environment and Norway rats are commonplace in the surrounding neighbourhoods (Easterbrook et al. 2005). Until 2004, JHMI maintained several breeding colonies of rats that had been in existence continuously for more than 20 years in unfiltered cages. Thus, they likely introduced infections common in laboratory rodents many years ago. Routine health surveillance of these colonies indicated infection with pinworms (S. muris), RCV/SDA, RPV, M. pulmonis, CARB, TMEV, P. pneumotropica and PVM. Since eliminating the infected colonies, JHMI has had outbreaks in barrier rodent colonies (from most to least common): pinworms (A. tetraptera and S. muris), mesostigmatid mites (L. echidninus and O. bacoti), parvoviruses (MPV and RPV) and mouse hepatitis virus (Watson J, unpublished data).
Pinworms were not found in wild rats in Baltimore; therefore, pinworm outbreaks at JHMI and other research institutions are most likely being perpetuated by environmental contamination or laboratory rodents. L. echidninus had not been found in JHMI rodent colonies or reported in other laboratory colonies before 2004; coincidentally, the source of the first JHMI outbreak proved to be a colony of research mice housed in a laboratory infested with commensal mice (Chu & Couro 2005, Coons 2005). Thus, L. echidninus and other ectoparasites (i.e. O. bacoti) may well have spread from wild rats via commensal mice to the laboratory mice. Recent mesostigmatid mite outbreaks may indicate increased infestation of wild rat populations or increased migration into the laboratory because of construction and/or displacement of wild rats from their natural habitat. Antibodies to various parvoviruses (i.e. KRV, H-1 and RPV/RMV) were detected in almost half of the wild Norway rats; thus, wild rats could contribute to parvovirus outbreaks in JHMI rodent colonies.
Antibodies against M. pulmonis, CARB and RCV/SDA were detected in over half of the sampled wild Norway rats. Although control of these laboratory infections has recently improved (Gaertner 2004), wild Norway rats may be involved in laboratory rodent colony outbreaks of M. pulmonis, CARB and RCV/SDA. Antibodies against TMEV and Sendai virus, however, were only detected in a few wild rats and antibodies against PVM and reovirus were not detected in the sampled wild rats. Laboratory rodent contamination with TMEV (or the rat TMEV-like virus), Sendai virus, reovirus and PVM are unlikely to have been caused by wild Norway rats. Although detected in wild rats, N. braziliensis, T. muris and R. nana or H. diminuta have not been recently reported in laboratory rodent populations (Livingston & Riley 2003). Although parvoviruses and helminth ova are both transmitted by the faecal–oral route, parvoviruses also are shed in urine, saliva and nasal secretions, so transmission often occurs by direct contact or fomites; thus, transmission of parvoviruses may more readily occur from wild rats as compared with transmission of helminths. Several pathogens that are common in laboratory rat colonies worldwide, including rat respiratory virus (RRV), Clostridium pilforme, Spironucleus sp. and Giardia sp., were not included in this study, but may be present in wild rat populations.
The effect of age on the serological analyses may be an artefact because only one juvenile rat was included. Juveniles were not selected for the serological screening because maternal antibodies are likely present in young animals and may overestimate seropositivity in juveniles and young rats have less time to have been exposed to a pathogen, thus underestimating the overall prevalence. Increasing age-related prevalence of N. braziliensis may reflect an increased probability of encountering the pathogen with lifespan, but this does not explain why other parasites examined were not affected by age. Seasonal patterns were observed for the prevalences of R. nana or H. diminuta (i.e. lowest during the spring) and N. braziliensis (i.e. highest during the fall); explanations for these observations are not known.
Ectoparasites were collected more frequently during the summer and fall than spring and winter. Whether temperature contributes to the prevalence of ectoparasites in wild-caught rats requires additional consideration.
Presence of antibodies to CARB and M. pulmonis, both of which are transmitted by aerosol and produce chronic infections, were significantly correlated and co-infection has been reported previously in laboratory rodents (Cundiff & Besch-Williford 1992). Dual infection with CARB and M. pulmonis produces more severe pulmonary lesions as compared with a single infection (Baker 1998). RCV/SDA also is transmitted by aerosol, but was not correlated with M. pulmonis or CARB, possibly because the distribution was skewed in which a majority of the rats had detectable antibodies against RCV/SDA.
Introduction of zoonotic pathogens into laboratory rodent colonies may not only compromise research, but also the health of the rodents and laboratory personnel. Infectious agents currently found in laboratory rodent colonies do not produce overt pathology in immunocompetent rodents, but still may alter results obtained for scientific research (Baker 1998). Increasing numbers of genetically altered rodents are utilized in research and may be more susceptible to morbidity and mortality caused by pathogens commonly identified in laboratory rodent colonies (Rozengurt & Sanchez 1993, Baker 1998). Laboratory animal handlers also are at risk to zoonotic pathogens carried by laboratory rodents. Antibodies against zoonotic agents, including Seoul virus (the hantavirus carried by Norway rats; 57.7%), hepatitis E virus (HEV; 73.5%), Leptospira interrogans (65.3%), Bartonella elizabethae (34.1%) and Rickettsia typhi (7.0%) have recently been identified in wild Norway rats (Easterbrook et al. 2007). Documented outbreaks among laboratory personnel caused by infected laboratory rodents have included tropical rat mites (i.e. O. bacoti), hantaviruses and Leptospira sp. (Sarasin et al. 1963, Fox 1982, Kawamata et al. 1987); therefore, animal handlers should be aware of the potential for zoonotic transmission of infectious agents.
Wild populations of Norway rats harbour many rodent-specific and zoonotic pathogens (Webster & Macdonald 1995, Easterbrook et al. 2007). Although most outbreaks in laboratory colonies are likely caused by introduction of an infected laboratory animal, wild Norway rats should be considered as an alternative potential source of contamination.
Footnotes
Acknowledgements
We would like to thank Darren Kaw for help with trapping the rats during summer 2005 and Andrew Glenn and Bruce Baldwin for help in identification of helminth adults and ova. Financial support was provided by NIH grant R01 A1054995 (SLK), Research Animal Resources (Johns Hopkins University) and NSF grant EF-0525751 (GEG).