
Abstract
Mycobacterium gordonae is an occasional human pathogen associated with cutaneous infections and nodular granulomatous skin lesions. A case of cutaneous nodular infection caused by M. gordonae in a colony of African clawed frogs (Xenopus tropicalis) is described and confirms this organism to be an opportunistic frog pathogen.
Amphibians of the genus Xenopus are increasingly used in biomedical research as they provide an invaluable model for studying fundamental cellular and developmental problems. 1 At MRC National Institute for Medical Research (MRC–NIMR), we house a large colony of the amphibian Xenopus tropicalis, a close relative of Xenopus laevis, which shares virtually all advantages of an embryological system 2 plus a much shorter generation time and a true diploid genome. 3
Mycobacteria are widely present in aquarium environments. 4 Amphibian mycobacteriosis has been described as a disease of the integument, and as a systemic disease with multiple gray nodules of varying sizes in the skin, liver, spleen, respiratory tract and intestinal tract. 5 Several species of Mycobacterium have been isolated in the past from X. laevis: M. marinum, 6,7 M. chelonae, 8–10 M. xenopi 8 and M. szulgai. 11 Recently, a species of Mycobacterium genetically related to M. ulcerans and M. marinum has been isolated from a colony of X. tropicalis 12,13 and X. laevis 14 and has provisionally been named M. liflandii. Infection with these bacteria can affect the experimental results by inducing inflammation, stress and causing death.
This report describes a cutaneous nodular infection in a colony of X. tropicalis caused by M. gordonae and the associated microbiological and pathological features.
Materials and methods
Animals
X. tropicalis were bred in-house and kept according to the UK Animals (Scientific Procedures) Act 1986. 15 Briefly, aquarium environmental conditions consisted of a 12:12 h light cycle, 25–26°C water temperature and 26°C ambient room temperature, pH range 6.5–7.5, electrical conductivity between 500 μS and 1200 µS, un-ionized ammonia <0.5 parts per million (ppm), nitrite <0.5 ppm and nitrates <50 ppm. Chlorine and chloramine were removed from the supply water using carbon filtration. Water was recirculated in a standalone system with manual top-up as required. Hardness of the water was kept at around 130–170 ppm. Water system and animals were monitored for microorganisms including Mycobacterium spp. at least four times a year. Frogs were housed in Z-Mod and XR4 systems (Marine Biotech, Inc, Beverly, MA, USA) and density varied between the different rooms and modules according to the European Union recommendations. 16 All tanks were fully cleaned every six weeks. Frogs were removed from the tanks during this cleaning process. Food and debris were removed from the tanks on a daily basis. UV filters were changed every 6 months or as needed.
Postmortem
Animals were euthanized by immersing in a buffered solution of tricaine methane sulphonate (MS222) (Pharmaq Ltd, Fordingbridge, Hampshire, UK). Tissues for histological examination were taken in 10% neutral buffered formaldehyde.
Microbiology
Specimens of skin lesions and swabs taken during postmortem were plated on Horse Blood agar (Oxoid Ltd, Hampshire, UK), McConkey agar (Oxoid Ltd) and Lowenstein-Jensen medium (BioMérieux UK Ltd, Basingstoke, Hampshire, UK) and incubated at 30°C in air. Biochemical identification methods used in our laboratory were in accordance with those described in Collins and Lyne's Microbiological Methods. 17
DNA extraction
DNA was extracted from bacteria using Qiagen QIAamp kit (Qiagen, West Sussex, UK) according to the manufacturer's instructions. The DNA was dissolved in 200 μL of elution buffer (AE buffer, Qiagen) and stored at −70°C. Polymerase chain reaction (PCR) products for cloning and sequencing were purified using QIAquick kit (Qiagen) according to the manufacturer's instructions.
PCR assay
PCR amplification of the 16S rRNA gene was carried out as previously described. 18 Amplification and enzyme digestion of the heat-shock protein 65 (hsp65) gene was carried out as previously described for identification of up to 34 Mycobacterium spp. 19,20 The internal transcribed spacer region (ITS) between 16S and 23S rRNA genes was amplified following the method described by Roth et al. 21 Briefly, PCRs were carried out as follows: 50 ng of template was added to a PCR mix containing 2 mmol/L dNTPs, 10 pmol of each primer and 2.5 U of HotStartTaq DNA polymerase (Qiagen) in PCR buffer containing 15 mmol/L MgCl2. PCR was performed using the Robocycler Gradient 96 (Stratagene, London, UK). Amplified DNA was resolved using gel electrophoresis in ethidium bromide prestained 1.5% agarose gels.
DNA cloning
DNA PCR products were cloned using pGEM-T Vector (Promega, Southampton, UK), according to the manufacturer's instructions. Briefly, PCR products were ligated using T4 DNA ligase (Promega) and the ligation transformed by heat shock at 42°C for 45 s into JM109 Escherichia coli competent cells (Promega). Cells were incubated in 1 mL of super optimal broth with catabolite repression (SOC) medium (2.0% tryptone, 0.5% yeast extract, 10 mmol/L NaCl, 2.5 mmol/L KCl, 10 mmol/L MgCl2, 20 mmol/L MgSO4 and 20 mmol/L glucose) and plated on Luria Bertani medium (1.0% tryptone, 0.5% yeast extract and 171 mmol/L NaCl) plates containing ampicillin (100 μg/mL), isopropyl-β-D-thiogalactopyranoside (IPTG) (0.5 mmol/L) and Xgal (80 μg/mL). Positive clones from each PCR product were sent for sequencing.
DNA sequencing and sequence analysis
Sequencing was carried out using the Big Dye Version 1.0 Sequence Reaction Kit (Applied Biosystems, Foster City, CA, USA). Samples were sequenced using an ABI Prism 3700 Capillary Sequencer (Applied Biosystems).
Sequences were edited and analysed using EditSeq, Megalign and SeqMan of Lasergene programs (DNAStar, Konstanz, Germany). Sequences were identified using BLAST from the NCBI (National Center for Biotechnology Information [
GeneBank accession numbers – 16S rRNA gene: EF428556, EU486079, EU486080; hsp65 gene: EF546780, EU486081; ITS: EU497913.
Results
Pathology study
The first case appeared in room 1 and subsequent cases appeared 6, 8 and 9 weeks later in rooms 1 and 3. More cases appeared 10 and 11 months later in rooms 1 and 3 again.
Postmortem examinations of affected frogs did not reveal macroscopic lesions in any organ other than the skin. Animals presented multiple raised cutaneous nodules of various sizes on different body locations (Figure 1). Samples from all organs were taken for histopathology. Microscopically, acid-fast positive bacilli were demonstrated within the cutaneous inflammation of frogs with clinical records 1, 20 and 577 (Figure 2). Sections of amphibian skin were characterized by subcutaneous nodular inflammation predominately composed of macrophages. Numerous lymphocytes were also present with the macrophage infiltration. Multifocally similar granulomatous inflammation overlaid small surface epidermal erosions. Numerous acid-fast positive bacilli were present within inflamed regions both within the macrophages and extracellular regions. In particular many bacilli were present in the granulomatous inflammation on the skin surface (Figure 2).
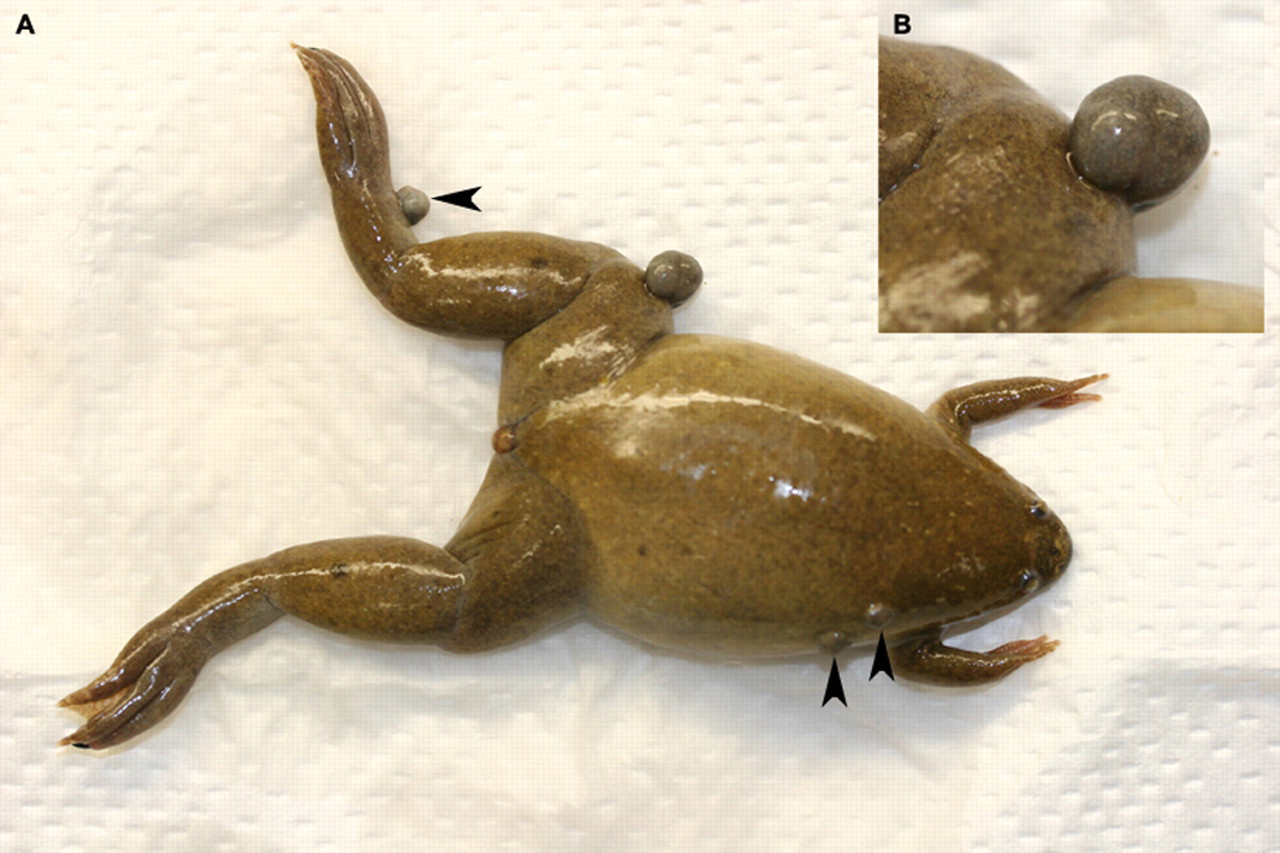
(A) Nodular granulomatous skin lesions (arrow heads). (B) Detail of the knee lesion
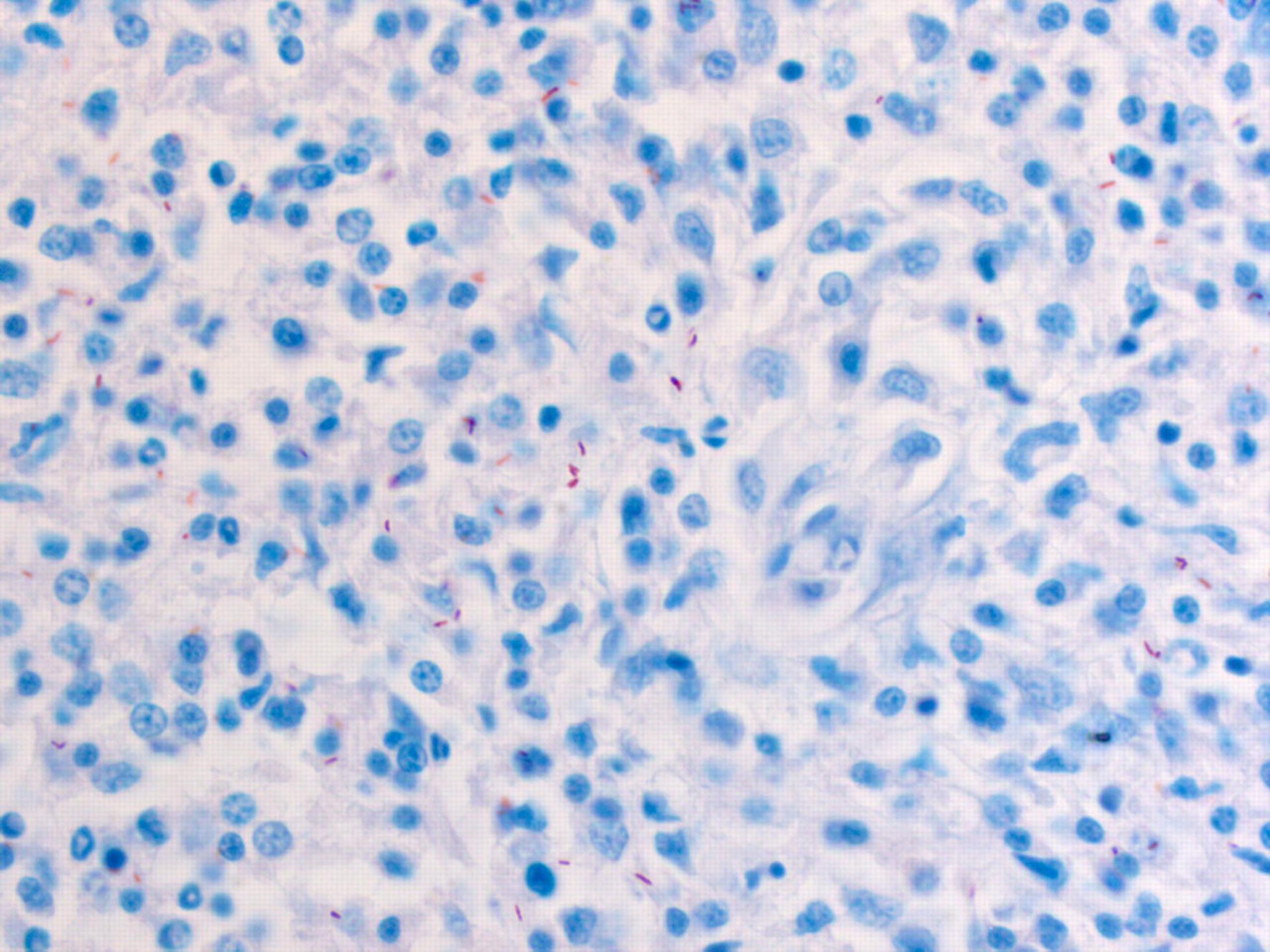
Microscopical details of the nodular granulomatous lesions seen in Figure 1
Microbiology
Specimens of skin tissue and swabs taken from the lesions were cultured from two clinical cases, clinical records 20 and 577, and a pure culture of Mycobacterium spp. recovered from skin tissue. After 10 days of incubation, yellow growth was evident. On Gram stain, the isolate was found to be a rod-shaped Gram-positive bacillus. Ziehl-Neelsen stain demonstrated that the isolate was an acid-fast positive bacillus. Results of biochemical characterization are shown in Table 1. Biochemical tests included the following: thiacetazone susceptibility, nitrate reduction, Tween 80 hydrolysis, tellurite reduction and growth in N-medium. The organism fulfilled the biochemical criteria for M. gordonae, being scotochromogenic, nitrate reductase (nitrase test) negative and Tween hydrolysis test positive. 22 Interestingly, the isolate grew on N-medium and presented growth in 10 days and grew better at 30°C than at 37°C. Samples from the culture were sent to the Health Protection Agency Mycobacterium Reference Unit (Bart's and the London Queen Mary School of Medicine and Dentistry, London, UK) and identification confirmed this to be M. gordonae.
Biochemical characterization of the Mycobacterium gordonae isolate
Molecular identification
PCR products were cloned into pGEM-T and 24 clones for 16S ribosomal RNA selected and sequenced for each isolate. The identity of the isolates was confirmed by 16S ribosomal RNA gene sequencing as M. gordonae (GeneBank accession numbers EF428556, EU486079 and EU486080). Amplification, enzyme digestion and sequence of the hsp65 gene was also carried out. 19,20 Enzyme digestion with BstEII gave one clear band of approximately 245 nucleotides plus a smear around the 120–100 nucleotide mark (Figure 3). Enzyme digestion with HaeIII gave us two bands of approximately 130/115 nucleotides plus a set of lower weight bands in the region of 30–40 nucleotides (Figure 3). This pattern obtained did not match with the published algorithm for M. gordonae. PCR products were cloned into pGEM-T and 12 clones for the hsp65 selected and sequenced for each isolate. The sequence identified was related to M. gordonae (GeneBank accession numbers EF546780 and EU486081) and confirmed a different pattern of enzyme digestion (BstEII, 347/94 both; HaeIII, 167/112/2 × 36/34/33/23 and 167/129/58/54/33, respectively). Finally, we amplified and sequenced the ITS region between 16S and 23S rRNA genes following the method described by Roth et al. 21 PCR products were cloned into pGEM-T and 12 clones for the ITS selected and sequenced for each isolate. The isolates were confirmed as M. gordonae and the sequence obtained was in one case identical to M. gordonae (strain 91-637) DNA (GeneBank accession numbers L42260 and EU497913).
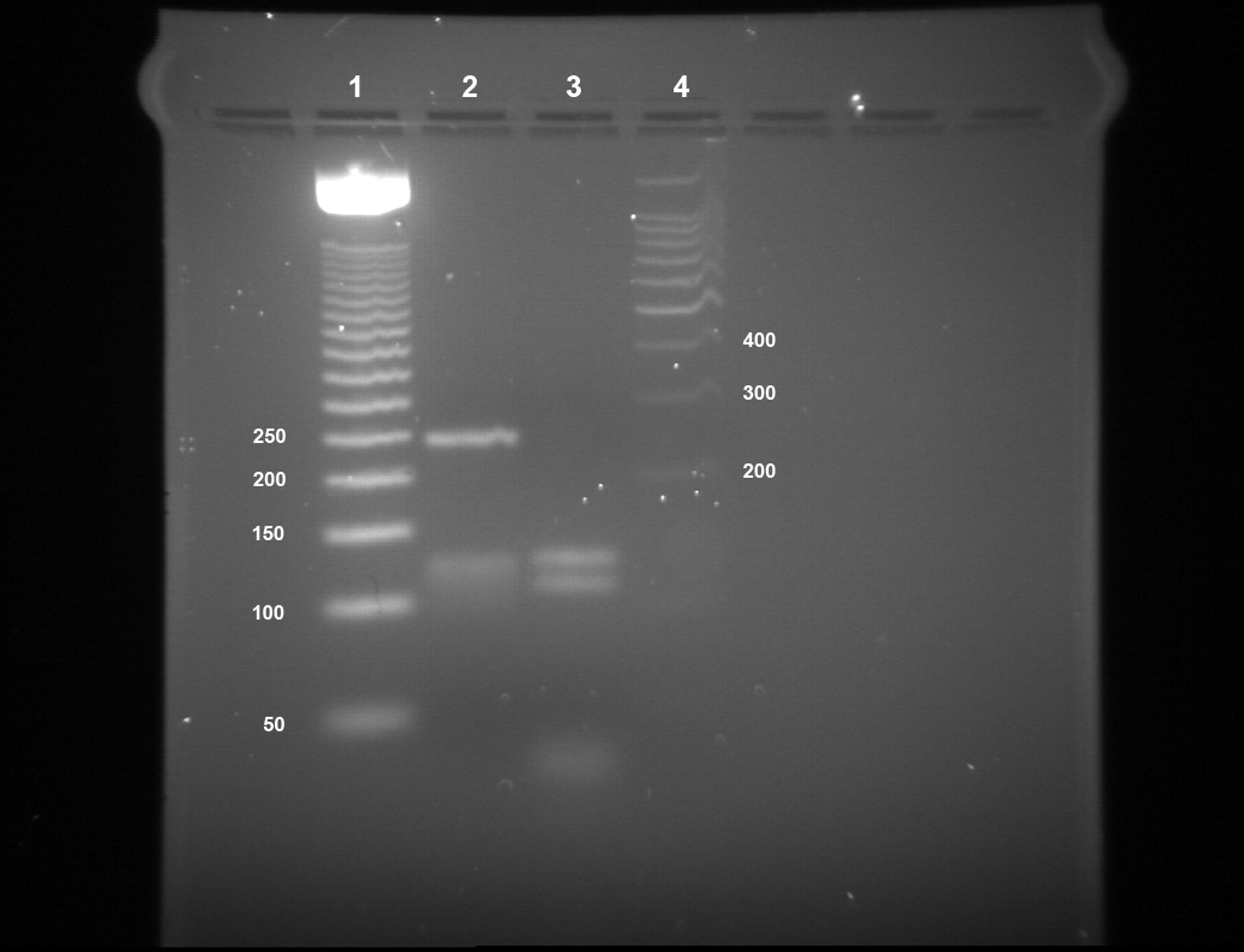
Polymerase chain reaction restriction fragment length polymorphism analysis of the Mycobacterium gordonae isolate hsp65 gene GeneBank EF546780. Lanes: 1, molecular weight marker (50 bp DNA ladder, Promega, Southampton, UK); 2, BstEII digestion pattern; 3, HaeIII digestion pattern; 4, molecular weight marker (100 bp DNA ladder, Promega)
Discussion
Here we have described a cutaneous nodular infection in a colony of X. tropicalis caused by M. gordonae and the associated microbiological and pathological features. Clinical presentation and pathology of M. gordonae infection in our colony is consistent with the presentation previously observed in human patients. 23–25 Our laboratory bred colony of X. tropicalis had been free of Mycobacterium spp. since the establishment of the colony in 2000 till this incidence. We have not isolated this organism from any other healthy or diseased animal species in our facility.
M. gordonae, usually considered a non-pathogenic commensal, may be found in soil and water 22 and is considered as an occasional human pathogen, especially in immunocompromised patients, 26–29 which has been associated with cutaneous infections and nodular granulomatous skin lesions. 23–25,30 Acid-fast bacilli have not been identified in all skin granulomas submitted for examination in this outbreak, as has been previously reported, 9 emphasizing the importance of including numerous tissue samples.
Because of known identification problems when using only 16S rRNA gene sequences, 13 we have used 16S rRNA sequence 31 hsp65-restriction fragment length polymorphism 20 and sequence, and ITS region sequence 21 plus biochemical properties to identify the isolated Mycobacterium. We would like to note that our M. gordonae isolate has a rapid growth (10 days) and grows on N-medium and thus does not completely fit with the phenotypic characteristics of the species.
The source of the infection in this report has not been identified, but M. gordonae is likely present in the animal facility environment. Multiple factors are likely to predispose X. tropicalis to clinical mycobacteriosis due to M. gordonae. However, the presentation pattern, few individual animals, indicates that stress (experimental manipulations and physical handling) and subsequent immunosuppresion may have been possible factors contributing to infection and disease in this M. gordonae infection. 9 X. tropicalis with signs of infection are now immediately separated from the rest of the animals and culled. There has been no transmission of M. gordonae from infected frogs to any human handlers in our facility.
This is the first report of M. gordonae causing disease in amphibians. The potential for human transmission warrants the use of preventive measures such as wearing protective clothing, gloves in particular, when working with amphibians.
Footnotes
ACKNOWLEDGEMENTS
We would like to thank Dr Kathleen E Mathers for critical reading of the manuscript. We would also like to acknowledge MRC–National Institute for Medical Research for their financial support to carry out this work.