
Abstract
The aim of the present study was to evaluate the behavioural and physiological responses to environmental disturbances (live and recorded dynamite explosions) in laboratory non-human primates in preparation for a future tunnel construction underneath our animal facility. In a pilot study (A) on 20 female Macaca fascicularis, a day of test blasts resulted in an increase in faecal cortisol and immunoreactive cortisol metabolites (CICM), and the animals reacted behaviourally with vertical flight and vocalizations. In a follow-up study (B), we assessed the impact of 10 days of exposure to recorded detonations on the behaviour and CICM in 16 M. fascicularis. In the latter study we introduced a predictive signal, serving as a conditional stimulus, to half of the animals. We found no significant effects of the noise in the Signal group; while the Control groups’ CICM values were affected. The behaviour was largely unaffected in the two groups. It was decided not to introduce a research moratorium on biomedical research planned to be conducted during the future tunnel construction, and that a conditional stimulus (‘warning signal’) will be used.
Non-human primates in captivity may sometimes be exposed to noise and vibrations in connection with construction work in or around the animal facility. Little is known about the extent to which captive animals experience stress and anxiety due to dynamite detonations that may occur during this kind of work (giant pandas, 1 snow leopards 2 ). Stress is known to have many physiological effects in different organ systems in the body, such as the immune system, the hormonal system and the gastrointestinal system. 3,4 A biomedical study involving assessment of variables related to these or other affected organ systems may obviously be compromised if the stress caused by the construction work gives rise to major changes in hormone levels, immune functions, etc. The present study was undertaken in an attempt to evaluate the potential stress effect caused by the construction of a tunnel underneath our primate facility due to take place within the next few years. The aim of the study was to assess the magnitude of the response in order to plan for a potential research moratorium or relocation of animals during the construction work, should the animals’ responses be too strong, both from a welfare perspective and to avoid potential confounding variables in future research endeavours. We conducted a pilot study (study A) during a test blast occasion. When that blast proved to affect the animals, we conducted a simulated blast study – for practical reasons involving noise only (study B) to further assess the effects of the environmental disturbance.
Quantitative measurements of faecal glucocorticoid metabolites have been used as a non-invasive biomarker of stress in several different mammal and avian species. 3,5–11 The aim of the present study was to evaluate the behavioural and physiological responses to environmental disturbances (live and recorded dynamite explosions) in cynomolgus monkeys (Macaca fascicularis) by measurement of concentrations of cortisol and immunoreactive cortisol metabolites (CICM) in single faecal droppings, and by monitoring the animals’ behaviour.
Materials and methods
Animals
The animals were housed at the Primate Research Centre of the Swedish Institute for Infectious Disease Control (SMI) in Solna, Sweden. All animals (both in study A and B) were born and raised in a breeding facility in China, quarantined in Holland for three months and had arrived together at SMI six months prior to a test blast carried out by the construction company to determine rock quality underneath the facility, which we documented in study A. Study A used 20 pair-housed female cynomolgus macaques (M. fascicularis) aged 4–5 years.
In study B, 16 pair-housed female cynomolgus macaques (13 of which had participated in study A), 6–7 years of age, were studied. As the animals were kept in two separate rooms, without visual and audible contact, we allocated each room to a different treatment, each consisting of eight individuals (4 pairs; subsequent analysis was done on each pair). This design was probably not ideal to yield truly independent samples since one single animal presumably could influence all other animals in the room by its behaviour, but it was the only practical solution.
In both studies, the animals were housed in pairs in stainless-steel barred cages (1 m×2 m×2 m, D×W×H) with a solid floor without bedding (a prerequisite of the Biosafety Level 3 housing), but otherwise enriched to give the animals the possibility of expressing their physiological and behavioural needs (daily refilling or replacing 5–8 enrichment options such as grain or rice spread in a foraging mat; treats in challenge balls or kong toys; shampoo bottles, old telephone books, tennis balls, aspen wood sticks, pine cones or dog bones to demolish; bath tubs with water for bathing; small amounts of straw, wood shavings or wood wool for foraging; PVC bottles stuffed with substrate and small treats, etc.). The room temperature was 22 ± 0.5 °C, relative humidity was kept above 50% and the ventilation rate was 15 air changes per hour. The lighting was provided at a 12/12 h schedule, with 30 min dusk and dawn, but natural light was also supplied through large windows. As a part of the daily routines, the animals were subjected to positive reinforcement training (PRT) in order to reduce the stress that may be involved in procedures and normal housing and care routines. The rooms were cleaned and the environmental enrichment items were changed or refilled every day between 07:00 h and 08:00 h. Pellets (Standard OWM (E); product code 808000, SDS Diets, Essex, UK) were given in conjunction with the morning cleaning, and different kinds of fruit and vegetables – three or four varieties – were given between 12:00 h and 13:00 h. Fresh drinking water was supplied daily. PRT took place before 09:00 h, between 12:00 h and 13:00 h, or after 16:00 h in study B.
The study was approved by the Stockholm North Ethical Committee on Animal Experiments (no. N30/06).
Project outline
The present investigation was divided into two different studies. In the pilot study (A), faecal CICM were determined and direct visual observation of the animals was performed during a series of test detonations on 7 July 2005. Faecal sampling commenced six days before the blasts and continued for 11 days in total. The second study (B) involved both ethological observations and faecal CICM determinations, and proceeded in four phases. In both studies, faecal sampling occurred every morning; 09:00–10:00 h in study A and 08:00–09:00 h in study B, by closing off half the cage to allow the technician access to faeces without animal contact. In study B, a first phase of six days was introduced, where the animals were habituated to this daily routine. In the second phase (5 days long), baseline faecal collection was carried out. During the third phase of 10 days, the animals were exposed to the noise recordings (see below) and continued faecal collection, and in the fourth phase, faecal collection was continued for 18 days after cessation of noise exposure (second baseline). Faecal CICM measurements were performed for all but the first phase (n = 33). Due to illness and technical failure, four days (days 6, 7, 17 and 32) collection of behavioural data was not made.
Exposure to detonations
In study A, the animals were subjected to six brief (<1 s) dynamite explosions occurring in a drilled hole approximately 20 m under the primate facility. The six detonations were performed during one day in one-hour intervals as a test of vibrations and noise in preparation for the future train tunnel construction. The amount of dynamite used in each detonation gradually increased, and ranged from 0.4 to 4.4 kg. At the time we were told that the actual detonations during the upcoming construction work would both last longer (7 s) and contain more dynamite; the explosions in study A were in this respect not representative of the future disturbance level. The noise level indoors in our facility measured up to 43 dB(A), and the vibration recordings showed a peak particle velocity (ppv) of up to 5.9 mm/s and a maximal acceleration of 2.9 m/s2.
The noise recordings presented to the animals during study B had been sampled during actual dynamite explosions performed during tunnel construction work, to the best of our knowledge captured the full range of sound and were thus better suited to mimic the future disturbance at our facility. The detonations were approximately 7 s long and were presented to the animals through high-quality speakers at a sound intensity of 92 dB(A), which was estimated to be the approximate noise level that would be generated from the real detonations during the future tunnel construction. One of the two groups (n = 8) was subjected to a stimulus acting as a Signal, consisting of a 30 s long piano melody, which ended 2 s before the dynamite explosions began. This classical conditioning served to give the animals some predictability over the environment. 12 The explosion recordings (and preceding conditional Signal for 8 of the animals) were played twice a day, at 10:00 h and 14:00 h, for 10 consecutive days.
Ethological observations
In study A, the animals’ behaviours were noted only in conjunction with the explosions, and some of the animals were also filmed during this event. No systematic ethological observations were performed during the days before and after the detonations.
The behavioural repertoire of the animals in study B was monitored daily during the investigation. The behavioural observations were performed using instantaneous sampling by studying films taken with permanent surveillance cameras present in the animal rooms. Each pair of animals was observed for a total of one hour per day, with selected behaviours (see Table 1) being sampled every 10th minute (6×2 data points) at exactly the same time every day. Observations were evenly distributed over the day (2 pair observations started with a few minutes’ interval at 09:04, 11:06, 13:01 and 15:07 h). The objective of the study was to measure any long-term effects of the noise; observations were not made earlier than one hour after the noise exposures so behavioural data collected were not the immediate surprise responses to the disturbance. Due to camera limitations, animal identification was not possible so each pair was allotted two data points per registration.
Studied and quantified behaviours in the ethological observations
In the analysis, object manipulation and eating/drinking/foraging were combined into the category ‘exploration’
Faecal CICM determinations
For estimation of total CICM in faeces, it has been deemed necessary to collect and weigh all faeces produced in the relevant time frame. 13,14
For practical applications, it is obviously an advantage to take samples from single droppings, without having to collect all faeces produced. There is evidence that the CICM concentrations in faeces may vary considerably over the day. 14 However, by taking the sample at the same time of day during a study, much of this variation is avoided. 15 Fresh faecal samples were taken at the same time every morning as soon as the animals defecated after cleaning, ensuring that no other droppings were present in the cage at the sampling occasion. In order to identify the individual faecal droppings, the dominant animal in each pair was given a piece of banana stained with non-toxic brilliant blue. 16 From each cage one brown and one blue entire fresh dropping was taken and mixed. Each joint sample was immediately frozen to −20°. The frozen samples were carefully stored in dry ice ensuring that they did not thaw during transport, and express-shipped to Copenhagen, Denmark for analysis.
The samples were thawed out and homogenized. From each sample 3 g of faeces was mixed with 3 mL of distilled water and vortexed for 30 min, after which 25 mL of 96% ethanol was added. After homogenizing and reduction to 10 mL, the mixture was then centrifuged, the supernatant transferred to glass tubes, and all liquid was evaporated. Dichloromethane (3 mL per tube) was added, and the tubes were vortexed and incubated for 30 min. After being transferred to smaller glass tubes, the organic solvent was left to evaporate. Enzyme-linked immunosorbent assay (ELISA) buffer (Cortisol EIA 1887, DRG Diagnostics, Marburg, Germany) was added, and the sample was ready for the ELISA procedure, which was carried out according to the manufacturer's manual.
Statistical methods
CICM values
Study A. A Wilcoxon matched-pairs signed rank test was performed on individual median values before (6 days) and after (5 days) the construction blasts, using Analyse-it.
Study B. Four samples were considered outliers (concentrations of over 100 ng/mL; probably an error in the analysis), and one was missing. These were replaced with median values of the day preceding and following the three faulty samples during noise exposure versus baseline 2. Two outliers of day 5 were recalculated as the median of days 4 and 3. 17
We tested whether exposure to construction noise affects the animals by using Runs tests for randomness. 18 An overall median value was calculated for the whole period for each group (Signal/No Signal). Then the median values of the cages per day within each respective group was calculated, and grouped into runs, defined as a series of observations (daily medians) in a sequence on the upper or lower side of the overall median value. The test statistic is computed by subtracting the expected runs from the observed runs divided by the square root of the variance of runs. The expected number of runs and the variance are functions of the numbers of daily medians below and over the overall median. These tests were performed in the two groups ‘Signal’ and ‘No Signal’ for the entire test period (33 days).
In order to test differences between groups (Signal/No Signal), exact permutation tests were used. This approach was chosen because assumptions about data belonging to a normal distribution was not fulfilled, and due to the small sample size of each group. Baseline values were summarized as median values from days 1 to 5 (before construction noise exposure) within cages. The following observations (days 7–33) were standardized according to these values to account for differences in baseline between the groups. After standardization, daily observations from the eight cages were ranked regardless of group membership. The test statistic from the sample was the sum of all rank values during the test period in the Signal group. We then calculated the probability of the sample results being random by permutating blocks (allocating the blocks in a random order without replacement to the two different groups) and for each permutation the test statistic was recalculated. We considered each cage during the test period as a block, which keeps the sample dependence within the cage over time. For each permutation run, k, the test statistic from the actual sample is compared with the test statistic from permutation k. Finally, the P value was derived by dividing the number of times that the permutation test statistic was equal or more extreme than the sample test statistic with the number of permutations.
Behavioural studies
Behavioural observations were expressed as the proportion of the total number of behaviours per cage and day. In order to test whether exposure to construction noise affected the animals, Runs tests on each behavioural category were used, using the overall median and daily medians for each group, as described above. Since behavioural data were missing from four days, these were simply excluded from all analyses regarding the behavioural observations.
We did the same types of permutation tests on the behavioural categories locomotion, exploration, pacing, grooming, inactivity and other as described for the cortisol values above in order to compare the Signal group with the No Signal group.
STATA version 10 and R version 2.9.2 were used for programming the statistic tests. In study B, one-tailed tests were used for cortisol calculations, and two-tailed tests in behavioural calculations. P values below or equal to 0.05 were considered significant and P ≤ 0.001 noted as (***) in the results.
Results
Study A
The primates’ behavioural reactions to the six detonations showed an approximate proportionality to the noise and vibration levels – or a sensitization effect. At the first detonation (0.4 kg of explosive), the animals showed almost no reaction. When later detonations occurred, the animals reacted with signs of fear, such as vocalizations and vertical flight reactions to higher cage levels. It is uncertain as to whether these behaviours were due to the effects of sensitization due to the repeated exposure to blasts with short rest intervals, or whether there was a threshold effect. Since no quantitative ethological observations were made, it is not known whether any overt signs of stress or anxiety remained.
The results of the faecal CICM determinations are shown in Figure 1. All individuals except two showed an increase in median CICM levels from the six days preceding the detonation exposure (median level 14.3 ng/mL) to the four days following the day of the explosions (21.0 ng/mL). The difference was significant: Wilcoxon P < 0.0001, Wilcoxon statistic: 5.
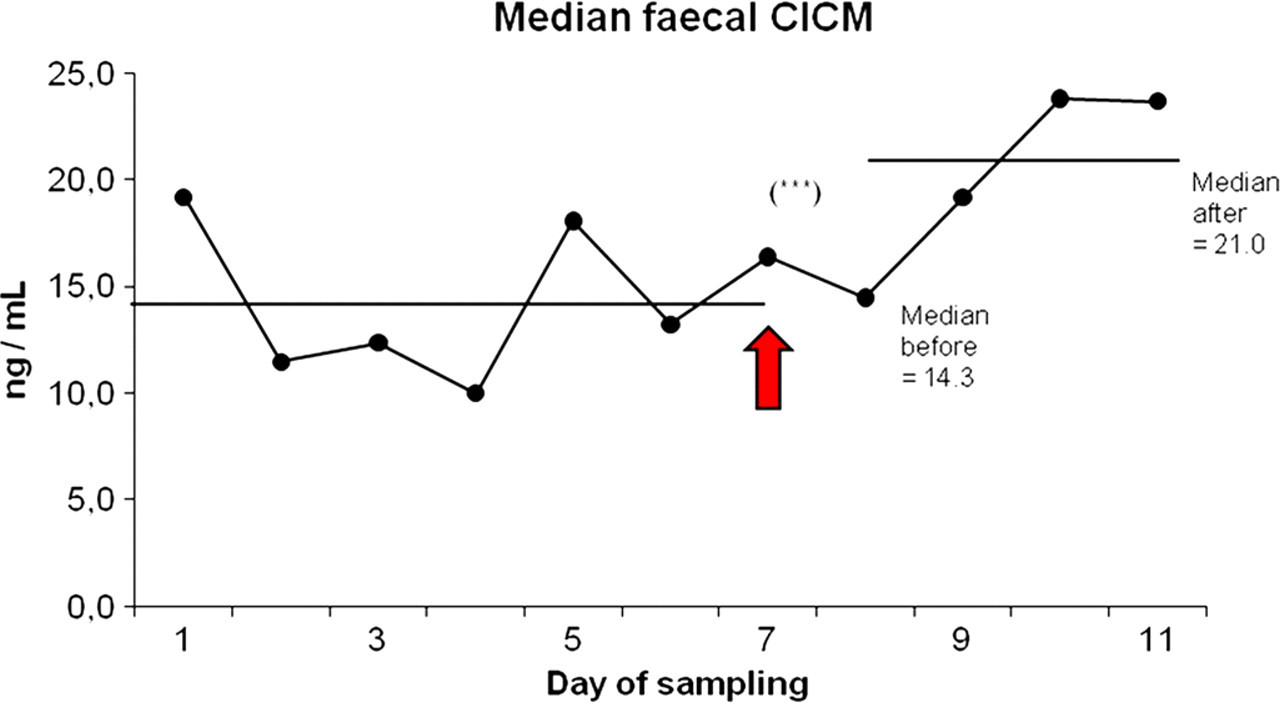
Daily median faecal cortisol and immunoreactive cortisol metabolite (CICM) concentrations in the 20 monkeys during study A. Overall median levels over the periods before and after the day of dynamite explosions (arrow) are indicated with solid horizontal lines. Values calculated after the explosions start 24 h after the first explosion (days 8–11). 18 ***=P < 0.001. See discussion for interpretation of results
Study B
Within the group which received no warning signal, we saw no statistically significant effects of the noise on behaviour over time, with one exception. Runs tests identified the category ‘other’ as non-randomly distributed over time. See Figure 2a and Table 2 for details.
Medians, test statistic and P value from Runs tests from the study period (days 1–33) for the Signal group and the No Signal group
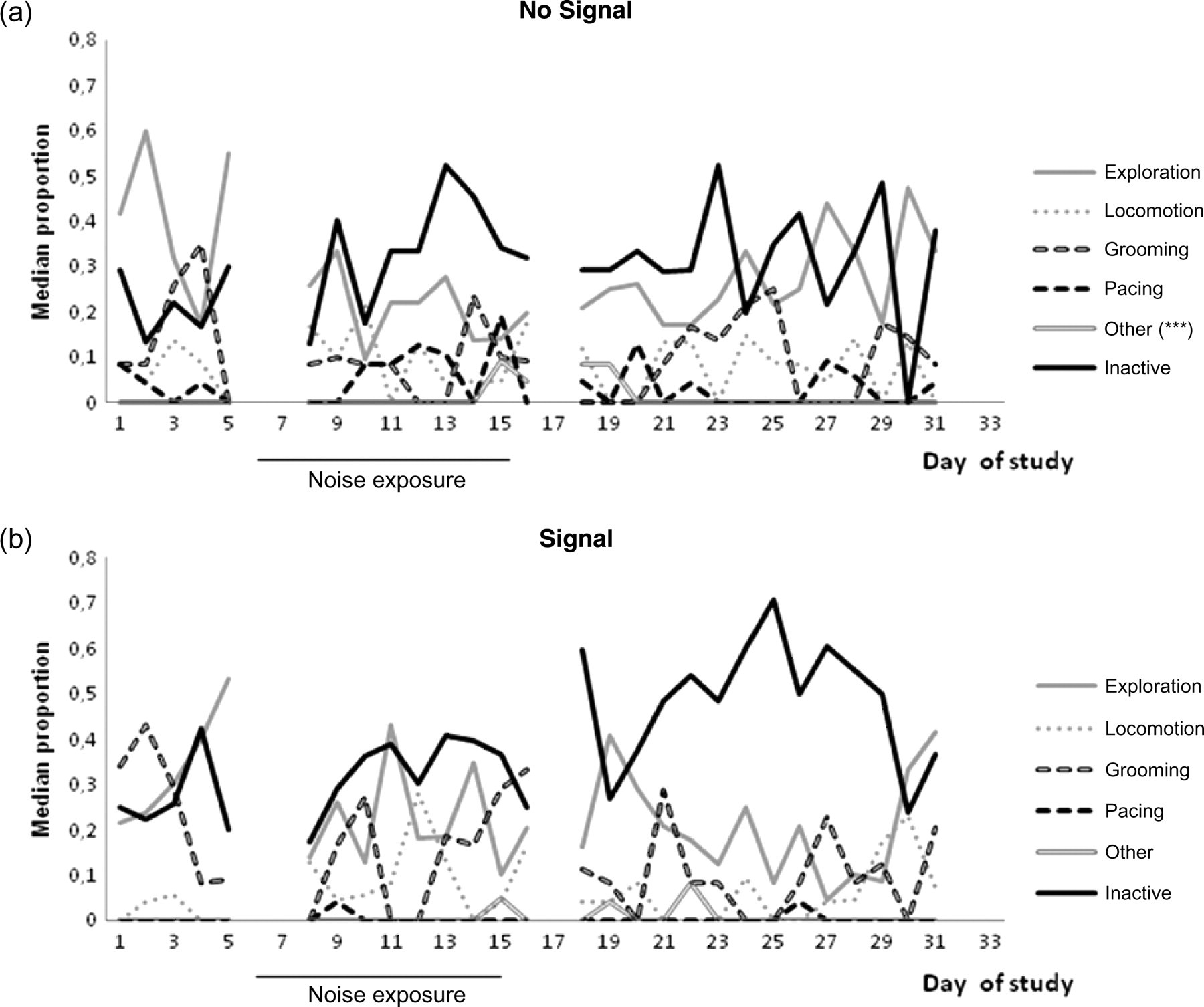
Study B: The daily median proportion of the different behavioural categories throughout the study (days 1–33) for two groups, each of eight pair-housed animals, subjected to explosion noise: (a) No Signal group which received no warning signal predicting the impending noise, and (b) Signal group which received a warning signal. Data are missing from days 6, 7, 17 and 32. Within-group analyses show the only significant difference is in ‘other’ behaviour (No Signal group ***=P = 0.001.)
Within the group which received a warning signal, no statistically significant effects of the noise on behaviour over time were identified, although there was a trend that inactivity increased after noise exposure. See Figure 2b and Table 2 for details.
When comparing the two treatment groups there was no statistically significant difference between the two groups (Signal/No Signal) with respect to behaviours shown (see Table 3).
Rank sums and P values from the exact permutation tests comparing the Signal group and the No Signal group after the first baseline (days 7–33)
The No Signal group showed a non-random distribution of daily median CICM values during the study period (Runs test, median = 30.03, z = −1.74, P = 0.041, one-tailed). No such effect was evident in the Signal group (Runs test, median = 20.96, z = −1.04, P = 0.145, one-tailed). The rank sums calculated in the exact permutation tests were 337 for the Signal group and 563 for the No Signal group. This difference was found to be significant (P = 0.014; one-tailed) (see Figure 3).
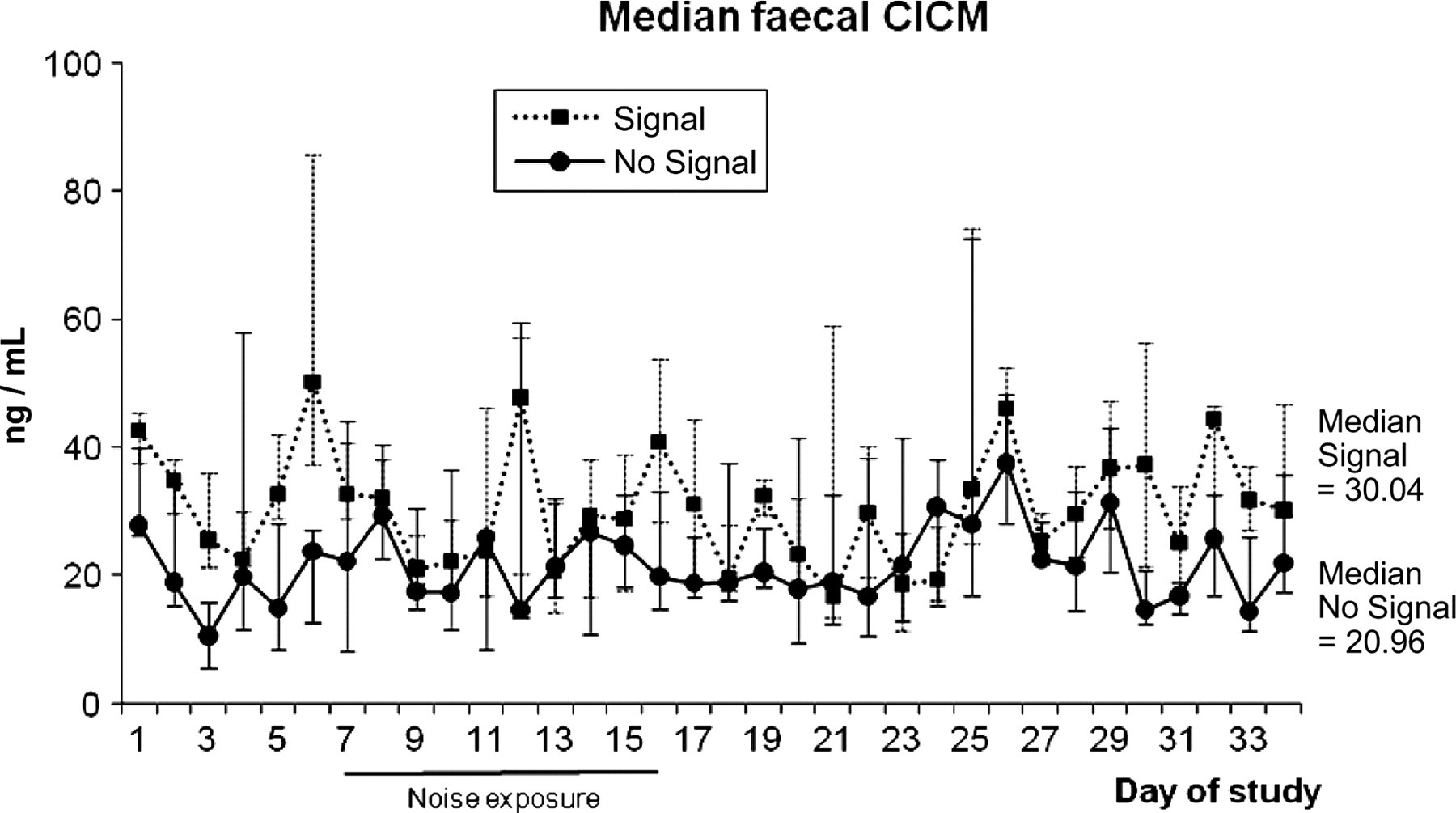
Daily median faecal cortisol and immunoreactive cortisol metabolite (CICM) levels during the different phases of study B (Signal [n = 4 cages, 8 animals]/No Signal [n = 4 cages, 8 animals]) before, during and after noise exposure. Vertical bars depict ranges (max/min). Differences were significant, see text
Discussion
The increase in faecal CICM concentration in the days following the real dynamite explosions in study A indicates a stress response in the monkeys. However, we were slightly puzzled by the timing of the onset of the elevated CICM reaction. The last, and most intense, blast took place at 14:50 h, and faecal samples were taken between 09:00 h and 10:00 h in the mornings. As seen in Figure 1, the CICM increase is apparent from day 9 of sampling (42 h after the last blast) and is even higher on the following sampling occasion (66 h after the last blast). Note that we have no measurements between 42 and 66 h post-blasting; it is possible that the peak occurred during this interval. In a previous study, the peak cortisol metabolite excretion occurred 26 h after intravenous cortisol administration in long-tailed macaques. 19 The timing of steroid passage in M. fascicularis from blood via bile through the gut until their metabolites are expelled in faeces depends on gut motility and seems to be highly variable, as evidenced by previous studies; up to 2.3 days in one study. 20 In chimpanzees, the peak of faecal cortisol excretion occurred after 48 h in one study, 6 whereas in another study 19 it peaked after only 22 h. Another study 21 showed that faecal steroid concentrations could actually increase during storage conditions, while the samples awaited analysis, and others have emphasized the importance of validation of new assays and cautioned about the potential pitfalls of using faecal samples to predict stress-sensitive markers. 13,14,22 It is therefore prudent to treat all samples within a specific study identically to avoid these types of errors, and use validated analysis kits, as was done in the present studies. Indeed, we could detect no other extraneous variable that could explain our results. No particular cases of aggression or social disturbance were noted, and there was no capture for biomedical procedures or other potentially stressful distractions. There are seasonal changes in vertebrate glucocorticoid production. 23 However, the sampling periods were only 11 days (study A) and 31 days (study B), so seasonal variations are not expected to greatly influence the samples. Synchronized ovarian cycling could potentially explain the results, although previous studies disagree whether or not cortisol varies throughout the menstrual cycle, where one article suggests not 24 and another found the opposite. 25 Data were not collected on whether our animals exhibited synchronous cycling or not.
The consistent result of CICM increase in 90% of the animals in study A shows that it is possible to demonstrate changes in corticosteroid levels by sampling single faecal droppings (but see refs. 13 and 14). Sampling at a specific time of day controls for the influence of normal circadian rhythm on corticosteroid secretion. 15
In study B, where the monkeys were exposed to only noise recordings of detonations, without the accompanying vibrations, the two groups differed regarding CICM levels. The group receiving no information about the impending construction noise showed a non-random variation in CICM over time, as evidenced by the Runs test. No such effect was apparent in the group which was exposed to a predictable construction noise (the Signal group). Indeed, when comparing the two groups and controlling for baseline differences, the No Signal group exhibited higher CICM levels during and after the noise exposure.
The lack of effect on monkeys of explosive sounds preceded by a predictable noise (the Signal group) in study B might indicate that the animals reacted much more strongly to actual construction work, including vibrations, than to the simulation we attempted – the mere sounds of explosions. Indeed, previous studies of the impact of ambient noise on animal welfare have shown results ranging from the mild and transitory to more profound effects; see review in ref. 1. Since we found an effect for the No Signal group, it is evident that the Signal served its purpose as a predictive Stimulus, 12 giving the animals a cue about upcoming events and in this case alleviating the stressful impact of the disturbance.
Other studies focusing on animals’ behavioural reactions to construction work have observed a species difference in coping with demolition/construction disturbance. A reduction in locomotion could be seen in snow leopards 2 in contrast to an increase of ‘restlessness’ in giant pandas 1 during construction days. In ref. 2 they describe what might almost be termed learned helplessness, 26 whereby the animals, unable to ‘escape from the presumably aversive noise … become lethargic instead’ (p. 204). They also noted that the leopards spent less time in the outdoor enclosure (close to the construction site) during noisy days, which would reflect a preference for undisturbed areas. Both learned helplessness and tonic immobility are characterized by inactivity or utter stillness. In the current study, we did not see any significant effect in any of the groups on the variation of inactivity, foraging or locomotion over time.
In the study of the giant pandas exposed to construction noise, 1 the authors noted that one of the two individuals stereotyped during work days, in contrast to non-work days. In contrast, in the present study the expression of abnormal behaviour did not deviate from a random distribution over the course of the study. Indeed, the only statistically significant effect on behaviour was the occurrence of the category ‘other’ during a few days at the end of the noise exposure and beginning of the second baseline in the No Signal group. This category included disparate behaviours like mounting or swinging or the occasional aggression but was not broken down into constituent behaviour during analysis. However, aggressive behaviour was studied systematically (sampling all occurrences during 10 min daily intervals) but was not affected by the noise (unpublished observation). In study B the animals were not monitored at the time of the noise exposure (observations began 1 h after exposure); we might however expect a startle response similar to the one observed in study A, at least for animals in the No Signal group. The main focus of study B was not to investigate the startle response but rather to determine any long-term effects of the blasts that might potentially influence the outcome of research endeavours. In retrospect we might have prioritized differently.
Predictability of a stressor has repeatedly been found to have pronounced effects on behaviour and physiology, and when offered a choice, most animals studied to date prefer predictable aversive signals to unpredictable ones, even when they are unable to escape them (reviewed in ref. 12). In an attempt to generate predictability, we introduced a Signal to one of the groups in study B. Previous results were corroborated in the current study, indeed we did find a difference between CICM values obtained between the No Signal and Signal groups. Although it was the No Signal group that exhibited the lowest levels of CICM before the noise study, it was that group that was affected by the noise by a proportionately larger increase in CICM during the rest of the study. Note also that the baseline occurrence of pacing behaviour was more pronounced in this group (cf. Figures 2 and 3), although there was no group difference in the effect of noise exposure (Table 3). These two findings may indeed be correlated, as established stereotypic behaviour has been shown to reduce physiological stress responses such as corticosteroid secretion – a proactive coping behaviour, 27 but see ref. 28. In our data, this seems true on a group level, as described above, but the correlation is not clear: one of the two pairs who never showed any pacing behaviour had the lowest median CICM value. Indeed, it has been hypothesized that stress levels of corticosteroids may enhance the acquisition and formation of stereotypic habits, at least in rodents. 27 The three ‘new’ animals not included in study A were all in the same treatment group (No Signal). However, we do not think this a problem in the interpretation of our results since (i) they had also been exposed to the test blast in study A (although no samples were taken) and (ii) they had been housed in the same room and with the same compatible partner for more than 1.5 years before the start of study B and had thus had ample time to acclimatize to the surroundings, including habituation to the sampling procedure.
Due to the short follow-up study after the detonations in study A (only 5 days), we did not see a return to normal levels within that time frame. We thus do not know the exact duration of the animals’ response after the single day of detonation. Effects of a short-term moderate activation of corticosteroids may indeed be adaptive, and should not necessarily be regarded as intrinsically undesirable and negative (unless they confound the result of biomedical studies). Such a response may trigger physiological and behavioural responses that may help the animal overcome the impact of the stressor by facilitating foraging and exploratory behaviour, including enhanced memory performance, as well as suppressing reproductive and territorial behaviour – and may even be termed ‘anti-stress’. 29 In contrast, previous studies have shown that it is chronic stress (e.g. long-term activations) or severe elevations of corticosteroids that is potentially harmful, see review. 12
On the basis of our two studies, we decided not to apply a beforehand moratorium to ongoing research endeavours at our facility during the upcoming tunnel construction, scheduled to take place immediately under our facility, and involving weeks of daily blasts. However, we intend to monitor our animals’ behaviour and physiology very closely and are prepared to stop any research, should the animals show signs of chronic stress. Indeed, one important element of construction work involving blasting is vibrations, which may heavily influence the animals (indeed, this might explain the difference in outcome between study A and B, as the former included vibrations and the latter did not). Furthermore, we contend that these studies were important from a three R's perspective. We refine the situation by giving all animals a predictive signal alerting of an immediate blast in connection with the construction event, and we reduce the risk of carrying out biomedical studies generating impaired data.
Footnotes
Acknowledgements
We gratefully acknowledge all the help from our animal caretakers involved in the daunting task of delivering blue bananas and collecting faeces. Personnel at Copenhagen University were instrumental in helping with cortisol analyses, and Mia Kling helped with statistics. The project management of Citybanan at the Swedish Transport Administration is greatly acknowledged for its support and consideration of our animals during the performance of the study. We also thank an anonymous referee for constructive comments on an earlier version of the manuscript.