
Abstract
Objective
The increased blood stasis and venous volume pressure causing tissue hypoxia are observed in both varicocele and varicose veins. Metallothionein (MT), a metal-binding protein, protects against cell apoptosis under hypoxic stress. It also plays an important role in collateral flow recovery and angiogenesis. We studied the distribution of hypoxia-inducible factor-1α (HIF-1α) and MT in varicocele and varicose veins.
Methods
The study specimens consisted of 1 cm venous segments that were obtained from 12 male patients during vascular stripping surgery for varicose veins and 1 cm of internal spermatic vein (ISV) obtained from 12 patients during left varicocele repair. The control samples of 1 cm ISV were obtained from 10 male patients who underwent left inguinal herniorrhaphy. All vascular specimens were analysed for HIF-1α and MT expression by immunoblotting, immunohistochemical (IHC) staining and confocal microscopy. Data were analysed using one-way analysis of variance with Tukey's comparison test.
Results
In both venous diseases, the increased expression of HIF-1α and MT compared with the control group (P < 0.05) and most of the proteins distributed over smooth muscle layers were detected by IHC staining; HIF-1α and MT in the muscle layer with co-localization, and MT overexpression especially located in the endothelium of both venous diseases under confocal microscopy.
Conclusions
Our results revealed the higher expression of HIF-1α and MT in varicocele and varicose veins than in the control group; MT overexpression in the muscle layer of both diseased vessels and especially located in the endothelium under confocal microscopy. MT has the function to protect vascular cells from apoptosis under hypoxia. Thus, this MT function may cause a decreased vascular cell apoptosis and then contribute to the dilated and thickened walls of varicocele and varicose veins.
Keywords
Introduction
Varicocele is characterized by dilated and thickened walls of the internal spermatic veins (ISVs). Although this condition is found in 15–20% of men1–3 and the recurrence rate (about 2–14% 4 ) after varicocele repair is still an issue, the investigation of these diseased vessels is very rare. Similarly, the prevalence of varicose veins is definitely underestimated, and they affect up to 40% of men and up to 51% of women. 5 The increased blood stasis and venous volume pressure causing tissue hypoxia are observed in both above venous diseases.6,7 However, hypoxia did not increase vascular cell death or vascular wall atrophy. 5 Conversely, the decreased vascular cell apoptosis causing the dilated and thickened walls were observed in both diseased vessels.5,8
Metallothionein (MT) is a heavy metal-binding protein with a low molecular weight. It has physiological functions, including the abilities to detoxify metals, to combat oxidative stress, to regulate essential biometals such as zinc and copper,9–11 and to protect cells against apoptosis under hypoxic conditions. 12 It also plays an important role in collateral flow recovery and angiogenesis. 13 This is the first report to study the relationship between MT and hypoxia-inducible factor-1α (HIF-1α) distribution in both diseased vessels (varicocele and varicose veins) of humans.
Materials and Methods
Patients and venous samples
This study included 34 young non-smoking patients who were recruited between May 2007 and October 2008. One study group consisted of 12 patients aged from 20 to 25 years old with left varicocele, grade 3, who underwent varicocele repair due to scrotal dragging pain. Varicocele was graded according to Dubin and Amelar. 14 To prevent inter-observer bias, all physical examinations were performed by one physician. The other study group consisted of 12 patients (6 men, 6 women) with a mean age of 56 ± 2.3 years who underwent vascular stripping surgery for varicose veins of the lower leg after colour Doppler ultrasound survey. The saphenous vein insufficiency was diagnosed in the supine position and a reverse flow wave-form lasting longer than two seconds at the Valsalva manoeuvre. The operative indications were leg pain, oedema or venous stasis disease. All patients were characterized as having primary varicosities, and their symptoms persisted for more than six months. According to the CEAP (clinical, aetiological, anatomical and pathological elements) classification for chronic lower-extremity venous disease, 15 most of the patients were in class 3 (n = 8) and others in class 4 (n = 4). A 1-cm venous segment was obtained to compare with the control group.
The control group consisted of 10 volunteers aged from 20 to 25 years old with indirect left inguinal hernia, for whom the possibility of varicocele was ruled out by physical examination and colour flow Doppler sonography (ISV diameter <2 mm), 16 and there was no venous disease by duplex ultrasound study. The same left inguinal surgical incision was performed in patients with hernia and varicocele; the 1-cm ISV was resected during the operation. Then, the vascular specimens were stored at –80°C for immunoblotting or were fixed in 10% formalin and then embedded in paraffin blocks for immunohistochemical (IHC) staining and double immunofluorescence (IF) staining for confocal laser scanning microscopic study. All specimens were removed only after written informed consent was obtained from the patients. This study was approved by the Institutional Review Board of Taichung Armed Forces General Hospital.
Antibodies
Five types of primary antibody were used in the present study: (1,2) HIF-1α: two rabbit polyclonal antibodies for immunoblotting (#3716; Cell Signaling, Beverly, MA, USA; dilution 1:200) and immunostaining (sc-10790; Santa Cruz, Santa Cruz, CA, USA; dilution 1:50); (3,4) MT: two mouse monoclonal antibodies for immunoblotting (M3009-10; United States Biological, Swampscott, MA, USA; dilution 1:4000) and immunostaining (MS-1175; Thermo Fisher Scientific, Fremont, CA, USA; dilution 1:150 and 1:50 for IHC and IF stains, respectively); and (5) β-actin: a mouse monoclonal antibody for immunoblotting (#8226, Abcam, Cambridge, MA, USA; dilution 1:4000).
The secondary antibodies for immunoblotting were horseradish peroxidase-conjugated goat anti-mouse IgG (#0031430; Pierce, Hercules, CA, USA) or goat anti-rabbit IgG (#0031460; Pierce). The secondary antibodies for immunostaining were provided in the commercial kit (PicTureTM; Zymed, South San Francisco, CA, USA) used for IHC staining and the Alexa-Fluor 488 conjugated goat anti-rabbit (Molecular Probes, Eugene, OR, USA; dilution 1:50) or the Alexa-Fluor 546 conjugated goat anti-mouse antibodies (Molecular Probes; dilution 1:200) for IF staining. Preliminary experiments of negative controls (sections stained with only primary or secondary antibodies) demonstrated that non-specific staining and over-staining of the background were not found (data not shown) in IF staining.
Immunoblotting
The method used was modified from that of our previous studies.8,17 Each sample stored at –80°C was homogenized on ice and then centrifuged at 13,000g at 4°C for 20 minutes. The supernatants were used for determination of protein concentrations and for immunoblotting. Protein concentrations were measured using reagents from the BCA Protein Assay Kit (#23225, Pierce), using bovine serum albumin as the standard (#23209, Pierce).
For immunoblotting, the antibodies for HIF-1α, MT and β-actin detected bands with molecular weights of about 120, 54 and 42 kDa, respectively. The blots were cut into upper and lower portions at feasible sites for incubation, incubated at 4°C overnight with the diluted primary antibodies, and then incubated with diluted secondary antibody for one hour. Finally, the immunoreactive bands were analysed using MCID software version 7.0 (Imaging Research, St Catharines, Ontario, Canada). The results were converted to numerical values in order to compare the relative protein abundance of the immunoreactive bands.
Immunohistochemical staining
This method was modified from a protocol used in our previous studies.6,17 The deparaffinized ISV sections (4 μm) were rinsed with phosphate-buffered saline (PBS). Endogenous peroxidase was inactivated by incubating the sections with 3% hydrogen peroxide. The sections were stained with primary antibody (HIF-1α or MT) before being analysed using a commercial kit. Negative control experiments, in which PBS was used instead of the primary antibody, were conducted to confirm the positive results for HIF-1α or MT. Finally, the sections were counterstained with haematoxylin (catalog no. 1.05175.0500; Merck, Darmstadt, Germany) and rinsed with tap water. Sections were observed by using a light microscope (BX50; Olympus, Tokyo, Japan).
Double immunofluorescent staining and confocal laser scanning microscopy
The method used for IF staining and microscopy was a modified version of that used in our previous study. 17 The sections were incubated at 4°C overnight with the diluted primary antibody and were then exposed to the respective secondary antibodies for one hour. Finally, the sections were covered by a slip with mounting solution (Zymed) before being viewed by confocal laser scanning microscopy.
To determine and compare the localization of specific proteins, IF-stained sections were examined using a Zeiss LSM 510 inverted laser scanning microscope equipped (Hamburg, Germany) with an argon laser (488 and 543 nm) for excitation. The IF images of HIF-1α and MT were obtained with the Alexa-Fluor 488/546 filter set (BP505-530 for 488 and LP 560 for 546) controlled by the software (Zeiss LSM Image Browser version 3.5.0.223; Hamburg, Germany). The filter set was used to separate and transmit the emission wavelengths of Alexa-488- and Alexa-546-conjugated antibodies to different photomultipliers. The micrographs taken from each photomultiplier were subsequently merged so that the different-coloured labels could be simultaneously visualized.
Statistical analysis
Data were analysed using the one-way analysis of variance with Tukey's comparison. P < 0.05 was considered statistically significant.
Results
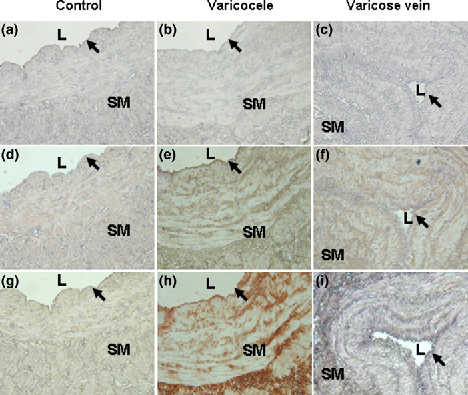
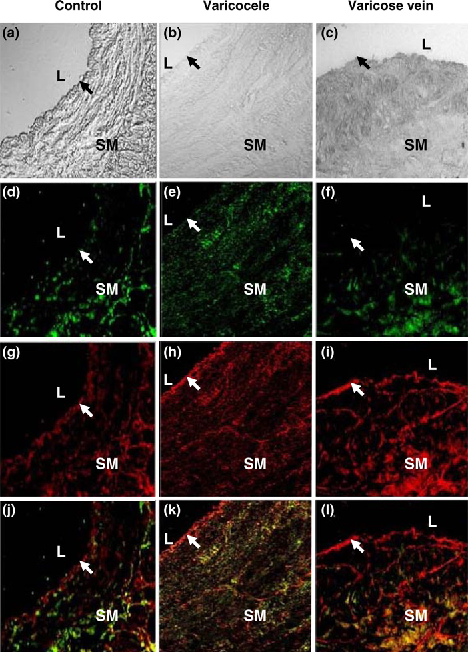
In the study groups, there was a higher expression of HIF-1α in varicocele and varicose veins compared with the control group (87.98 ± 3.53, 92.61 ± 5.65 and 26.54 ± 3.82, respectively, P < 0.05; Table 1 and Figure 1). Similar results were obtained for higher MT expression in the study groups than the control group (55.18 ± 2.73, 48.01 ± 3.64 and 26.27 ± 9.26, respectively, P < 0.05; Table 1 and Figure 2). The most HIF-1α and MT were distributed over the smooth muscle layer, as demonstrated by IHC staining (Figure 3). Confocal microscopy showed the HIF-1α (green colour) and MT (red colour) distributions in the muscle layer with co-localization, and MT overexpression especially located in the endothelium of both diseased vessels (Figure 4).
Representative immunoblot and relative intensities of hypoxia-inducible factor-1α (HIF-1α) protein expression in three venous groups. β-actin was used as the loading control. Different letters indicated significant differences (P < 0.05) Representative immunoblot and relative intensities of metallothionein (MT) protein in three venous groups. β-actin was used as the loading control. Different letters indicated significant differences (P < 0.05) Micrographs of representative sections of control (a, d, g), varicocele (b, e, h) and varicose vein (c, f, i) after haematoxylin staining (negative control, a–c), hypoxia-inducible factor-1α (HIF-1α) immunostaining (d–f) or metallothionein (MT) immunostaining (g–i). L: lumen; SM: smooth muscle layer; black arrow: endothelium. Magnification: ×100 (c, f, i) and ×200 (others) Representative sections of patients with control (a, d, g, j), varicocele (b, e, h, k) and varicose vein (c, f, i, l) after double immunofluorescence staining and confocal laser scanning microscopy for visible-light image (a–c), hypoxia-inducible factor-1α (HIF-1α) (green colour; d–f), metallothionein (MT) (red colour; g–i) and a merged image (j–l). L: lumen; SM: smooth muscle layer; arrow: endothelium. Magnification: ×200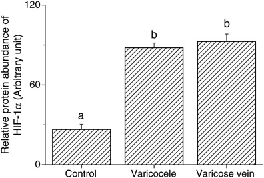
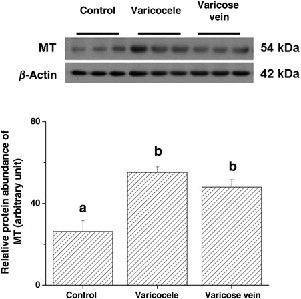
Expression of hypoxia-inducible factor-1α (HIF-1α) and metallothionein (MT) in three venous groups

Dissimilar letters indicated significant differences among three groups (P < 0.05)
Discussion
Both varicocele and varicose veins are venous diseases characterized by tortuous, dilated and thickened vein walls as a compensatory result of increased blood stasis and venous volume pressure.5,18–20 Both vascular diseases also affect millions of people, but the investigation of their mechanism is still very poor. We had reported that HIF-1α expression in the ISV of varicocele patients was significantly higher than in the control group. 6 Similarly, the varicose veins occurrence because of poor venous return and blood stagnation causing tissue hypoxia is associated with re-modelling of vessel wall. Blood stasis will provoke an ischaemia, thus decreasing oxygen availability to tissues. Under hypoxia, the vascular endothelium is the starting point of a cascade of events leading to the disorganization of the vessel wall typical of these pathologies.18,21
By reviewing previous papers, various functions of MT are roughly classified as (1) to regulate the activity of transcription factors, involved in cell proliferation, differentiation and apoptosis; (2) to control the redox status by reducing oxyradicals and nitrogenoxides; and (3) to bind to and protect from toxic heavy metals such as cadmium and mercury.22–25 MT is also involved in collateral vessel development: the endothelial cells, vascular smooth muscle cells and the macrophage. 13 Especially, endothelial cells play a major role in angiogenesis 13 and this may be the reason why MT overexpression is significantly located in the endothelium as in our study findings.
Recently, more studies have shown that MT is upregulated under hypoxic conditions to protect cells from apoptosis.26–28 Similarly, the hypoxic factor is a co-activator of MT-I gene expression to reduce the production of hypoxia-induced reactive oxygen species.27–29 Lim and Davies 30 have reviewed the pathogenesis of varicose veins and these reveal that areas of intimal hyperplasia and smooth muscle cell proliferation are often noted in varicose veins compatible with our findings, although regions of atrophy are also present.
In the present study, there is higher expression of MT and HIF-1α in the muscle layer with co-localization of both diseased vessels and MT overexpression especially located in the endothelium under confocal microscopy. MT has the function to protect cells from apoptosis under hypoxic stress. This function may cause a decreased vascular cell apoptosis and then contribute to the dilated and thickened walls of patients with varicocele or varicose veins. Finally, the limitation of this study is the small case number and lack of normal saphenous veins as a control group. Of course, we have not studied the mechanism of how the MT protect cells against apoptosis under hypoxic stress just by reviewing the previous studies.
Conclusions
Our results revealed the higher expression of HIF-1α and MT in varicocele and varicose veins than in the control group. MT overexpression in the muscle layer of both diseased vessels and especially located in the endothelium under confocal microscopy. MT has the function to protect vascular cells from apoptosis under hypoxia. Thus, this MT function may cause a decreased vascular cell apoptosis and then contribute to the dilated and thickened walls of varicocele and varicose veins.
Conflict of interest
The authors have no commercial, proprietary or financial interests in any companies or products described in this paper.
Footnotes
Acknowledgements
This study was supported by a grant from Taichung Armed Forces General Hospital, Taiwan, Republic of China.