
Abstract
More than 10,000 Caspian seals (Phoca caspica) were reported dead in the Caspian Sea during spring and summer 2000. We performed necropsies and extensive laboratory analyses on 18 seals, as well as examination of the pattern of strandings and variation in weather in recent years, to identify the cause of mortality and potential contributory factors. The monthly stranding rate in 2000 was up to 2.8 times the historic mean. It was preceded by an unusually mild winter, as observed before in mass mortality events of pinnipeds. The primary diagnosis in 11 of 13 seals was canine distemper, characterized by broncho-interstitial pneumonia, lymphocytic necrosis and depletion in lymphoid organs, and the presence of typical intracytoplasmic inclusion bodies in multiple epithelia. Canine distemper virus infection was confirmed by phylogenetic analysis of reverse transcriptase-polymerase chain reaction products. Organochlorine and zinc concentrations in tissues of seals with canine distemper were comparable to those of Caspian seals in previous years. Concurrent bacterial infections that may have contributed to the mortality of the seals included Bordetella bronchiseptica (4/8 seals), Streptococcus phocae (3/8), Salmonella dublin (1/8), and S. choleraesuis (1/8). A newly identified bacterium, Corynebacterium caspium, was associated with balanoposthitis in one seal. Several infectious and parasitic organisms, including poxvirus, Atopobacter phocae, Eimeria- and Sarcocystis-like organisms, and Halarachne sp. were identified in Caspian seals for the first time.
Keywords
In the spring and summer of 2000, there was apparent increased mortality of Caspian seals (Phoca caspica), a species listed as vulnerable by the World Conservation Union. 5 Although canine distemper was identified as the primary cause in a preliminary report, 29 a comprehensive analysis of this mortality event has not yet been performed. The Caspian seal is a small phocid seal that is limited to the Caspian Sea, where it inhabits all parts (Fig. 1). 32 The Caspian Sea is bordered by Russia, Kazakhstan, Azerbaijan, Turkmenistan, and Iran and measures approximately 1,200 km from north to south, with an average width of about 320 km. From late January to the end of April, most seals congregate in large groups on ice in the northern Caspian Sea. 32 The young are born on stable ice from the end of January to the beginning of February and are weaned after 3 to 4 weeks. 33 From May to September, seals migrate south and form groups in feeding areas in the central and southern Caspian Sea. In October and November, they migrate back north and congregate on islands in the eastern part of the northern Caspian Sea until the ice forms. 32 Limited aerial surveys of breeding areas in the northern Caspian Sea indicate that numbers of breeding females decreased from 90,500 in 1973 to 46,800 in 1989. Based on the latter figure, the total size of the Caspian seal population in 1989 was roughly estimated at 360,000 to 400,000. 32 Since then, the population size has not been determined.

Map of the Caspian Sea, showing the position of places mentioned in the text.
Unusually high mortality of Caspian seals in spring and summer of 2000, with more than 10,000 seals estimated dead, was reported in various media in that year. The first report was on 25 April, when 50 first-year seals were found dead on Zuidwest Island, 5 km from the mouth of the Ural River, northwest Caspian Sea (Fig. 1). It was reported that 1,300 seal carcasses were collected and destroyed on the Mangyshlak Peninsula, Kazakhstan, northeast Caspian Sea on 19 May and that 3,654 seal carcasses from the coast of Kazakhstan were destroyed on 1 June. By 23 June, this number had increased to 11,000. On 10 July, more than 300 seals were reported dead on the coast of Turkmenistan, near the Kazakhstan border, with the first deaths having been recorded in early April.
There has been a recent worldwide increase in mass mortalities of marine species, from corals to marine mammals. Both climate change and human activities have been implicated in these events. 21 Documented causes of mass mortality in pinnipeds include influenza in harbor seals (Phoca vitulina), 18 canine distemper in Baikal seals (Phoca sibirica), 19, 42 phocine distemper in harbor and gray seals (Halichoerus grypus), 25, 31, 41 and domoic acid poisoning in California sea lions (Zalophus californianus). 48 Furthermore, there is experimental evidence that the concentrations of anthropogenic pollutants in tissues of harbor seals from the North Sea were sufficiently high to cause suppression of their immune systems, and so may have increased the mortality rate during the phocine distemper epidemic in this species in 1988. 12, 47
To identify the cause of mortality, necropsies were performed on 18 Caspian seals found dead or dying during the spring and summer of 2000. In this article, the results of pathologic, virologic, bacteriologic, parasitologic, and toxicologic analyses are presented. The pattern of strandings in recent years and the possible effects of weather variation on the observed mortality are also examined.
Materials and Methods
Air temperature and disappearance of ice cover
The mean daily air temperatures in the northern Caspian Sea from November to March from 1990 to 2001 were obtained from the Kazakh Hydrometeorological Service Center, Almaty, Kazakhstan. The average of the mean daily air temperatures was used as the mean monthly air temperature. From the same source and for the same period, the dates of disappearance of ice cover were obtained for the northern Caspian Sea at Peshnoi (46°52′N, 51°42′E) and at Fort Shevchenko (44°31′N, 50°16′E), both in Kazakhstan (Fig. 1).
Strandings of Caspian seals in Azerbaijan
Starting in 1997, weekly surveys of a 10-km stretch of beach at Azizbekov (40°30′N, 50°10′E), on the north shore of the Apsheron Peninsula, Azerbaijan, were performed by car (Fig. 1). The age category and sex were determined of each stranded seal (i.e., a seal found moribund or dead on the shoreline). During the mortality event in 2000, extra surveys of beaches elsewhere on the Apsheron Peninsula were performed between 2 and 11 June 2000 at Pirshaga (40°35′N, 49°54′E), Kalaga (40°36′N, 49°58′E), and Adsyz (40°35′N, 49°47′E). The standard length, 2 sex, and stage of decomposition of each seal were determined. Seals with a standard length of ≤100 cm were considered 0 to 1 year of age, and seals with a standard length of >100 cm were considered >1 year of age. 39 Seals estimated to have been dead for zero to a few days were categorized as freshly dead to mildly decomposed; seals estimated to have been dead for a few days to 1 month were categorized as moderately to markedly decomposed; and seals estimated to have been dead for more than 1 month were categorized as skeletal.
Gross pathology
Eighteen Caspian seals found dead or dying between 3 May and 12 August 2000 were examined (Table 1). Gross necropsies on seal Nos. 1, 2, 8–15, 17, and 18 were performed by veterinarians; those on seal Nos. 8–15 according to a standard protocol, 34 those on seal Nos. 1, 2, 17, and 18 according to a reduced protocol. Gross necropsies on seal Nos. 3–7, and 16 were performed by nonveterinarians according to a reduced protocol.
Stranding data, signalment, and primary diagnoses of 18 Caspian seals found stranded during the canine distemper epidemic.





Strandings = a seal found dead or moribund on the shoreline; KZ = Kazakhstan; AZ = Azerbaijan; TU = Turkmenistan.
Age determined from number of growth layer groups in canine teeth27 or estimated based on standard length.39
At midsternal level.
Histopathology
Samples of adrenal gland (6 seals), aorta (2), brain stem (4), bronchus (3), cecum (3), cerebellum (9), cerebrum (5), colon (3), common bile duct (1), duodenum (1), eye (1), epididymis (1), eyelid (2), fallopian tube (2), gall bladder (2), heart (6), ileum (3), inguinal lymph node (1), jejunum (5), kidney (12), larynx (1), liver (12), lung (13), medulla oblongata (1), mesenteric lymph node (3), esophagus (3), ovary (3), pancreas (10), penis (1), prepuce (2), prostate (1), skeletal muscle (2), skin (4), spleen (13), stomach (5), testis (2), thymus (3), thyroid (3), tongue (1), tonsil (1), trachea (4), tracheo-bronchial lymph node (2), unspecified lymph node (4), urinary bladder (5), and uterus (4) were fixed in 10% neutral-buffered formalin and embedded in paraffin. Sections were cut at 5 µm, stained with hematoxylin and eosin, and examined by light microscopy. Selected sections also were stained with the Gram, Ziehl-Neelsen, and Grocott stains, and periodic acid Schiff method for identification of bacteria, protozoa, and fungi.
Transmission electron microscopy
Samples of skin from seal No. 15 were fixed in 4% formaldehyde and 1% glutar(di)aldehyde after brief fixation in 10% neutral-buffered formalin, and afterwards fixed in 1% osmium tetroxide. After embedding in epoxy resin, thin sections were prepared, stained with 6% saturated uranyl acetate and lead citrate, and examined using a Philips Morgagni 268D electron microscope (Eindhoven, The Netherlands).
Immunohistochemistry for morbillivirus
Paraffin-embedded sections of the same tissues prepared for histopathology for each seal also were examined for morbillivirus antigen by an immunohistochemical technique. 29
Reverse transcriptase–polymerase chain reaction for influenza A and B virus and morbillivirus
Samples of brain, lung, kidney, and urinary bladder were preserved at room temperature in a commercial lysis buffer from the High Pure RNA Isolation Kit (Roche Diagnostics, Indianapolis, IN) until use. All four tissues from 14 seals were examined for morbillivirus nucleic acid by reverse transcriptase–polymerase chain reaction (RT-PCR), and resulting PCR products were sequenced for phylogenetic comparison. 29 Samples of lung from all 14 seals except Nos. 3–7, 17, and 18 were examined for the presence of influenza A or B virus nucleic acid by RT-PCR. 43
Serology
Blood samples were collected postmortem from 13 seals, stored at −20°C until use, and tested for canine distemper virus (CDV)-specific immunoglobulin (Ig)M and IgG by an antibody-capture enzyme-linked immunosorbent assay (ELISA) and an indirect ELISA, respectively. 29 Sera of seal Nos. 8, 9, 11, and 13 were also tested for Brucella-specific antibodies by competitive ELISA and indirect ELISA. 26
Bacteriology
Samples were collected from 10 seals for bacteriological culture. All tissues were cultured as previously reported 17 on Columbia sheep blood agar and Farrells medium (Oxoid, Basingstoke, UK), incubated at 37°C in a capnophilic atmosphere, and examined daily for up to 7 days, and on MacConkey agar (Oxoid), incubated aerobically at 37°C, and examined after 24 and 48 hours. Intestinal samples were also enriched in selenite broth, incubated at 37°C for 18–24 hours, after which they were subcultured on brilliant green agar (modified) and further incubated at 37°C for 18–24 hours. Isolates were selected for further identification using a combination of standard bacteriological methods and identification kits (BioMerieux, Basingstoke, UK). Salmonella isolates were serotyped at the Scottish Salmonella Reference Laboratory (Stobhill, Glasgow, UK) according to standard procedures. 46 Bordetella bronchiseptica isolates were ribotyped as previously described at the Avian and Swine Respiratory Diseases Research Unit (Ames, Iowa). 45
Parasitology
Parasites found at gross necropsy were fixed in 70% ethanol and stored at room temperature until identification. 11, 32, 33, 44
Algal toxins
Samples of liver or kidney from eight seals were stored at −20°C until analysis by a colorimetric protein phosphatase 1 inhibition assay for microcystins and nodularins. 52 Samples analyzed by ELISA from three of these seals were diluted with Milli-RO® water to a final methanol concentration of 10% (vol/vol) methanol. The antimicrocystin ELISA kit (Envirologix, Portland, ME), which cross-reacts with a number of microcystins and nodularins, was used according to the manufacturer's instructions.
Toxicology
We reinterpreted the toxicological data on the Caspian seals found stranded in 2000, by using the results of pathologic and virologic analyses to categorize the data according to the canine distemper status of the seals. Most toxicologic data were obtained from published sources. 3, 27 For seal Nos. 3 to 5, unpublished toxicological data, which had been determined according to the same methods, were kindly provided by Robin Law and Colin Allchin (Centre for Environment, Fisheries and Aquaculture Science, Burnham-on-Crouch, UK). In our reinterpretation, we concentrated on organochlorines and zinc, because preliminary analyses had indicated that their concentrations in tissues of Caspian seals stranded in 2000 were higher than in previous years. 3, 27
Results
Air temperature and disappearance of ice cover
The mean monthly air temperatures in the winter 1999/2000 were higher than the 95% confidence interval over the period 1990/1991 to 2000/2001 (Fig. 2). Ice cover disappeared earlier in the winter of 1999/2000 than in any other winter in the period 1990/1991 to 2000/2001 (Fig. 3).
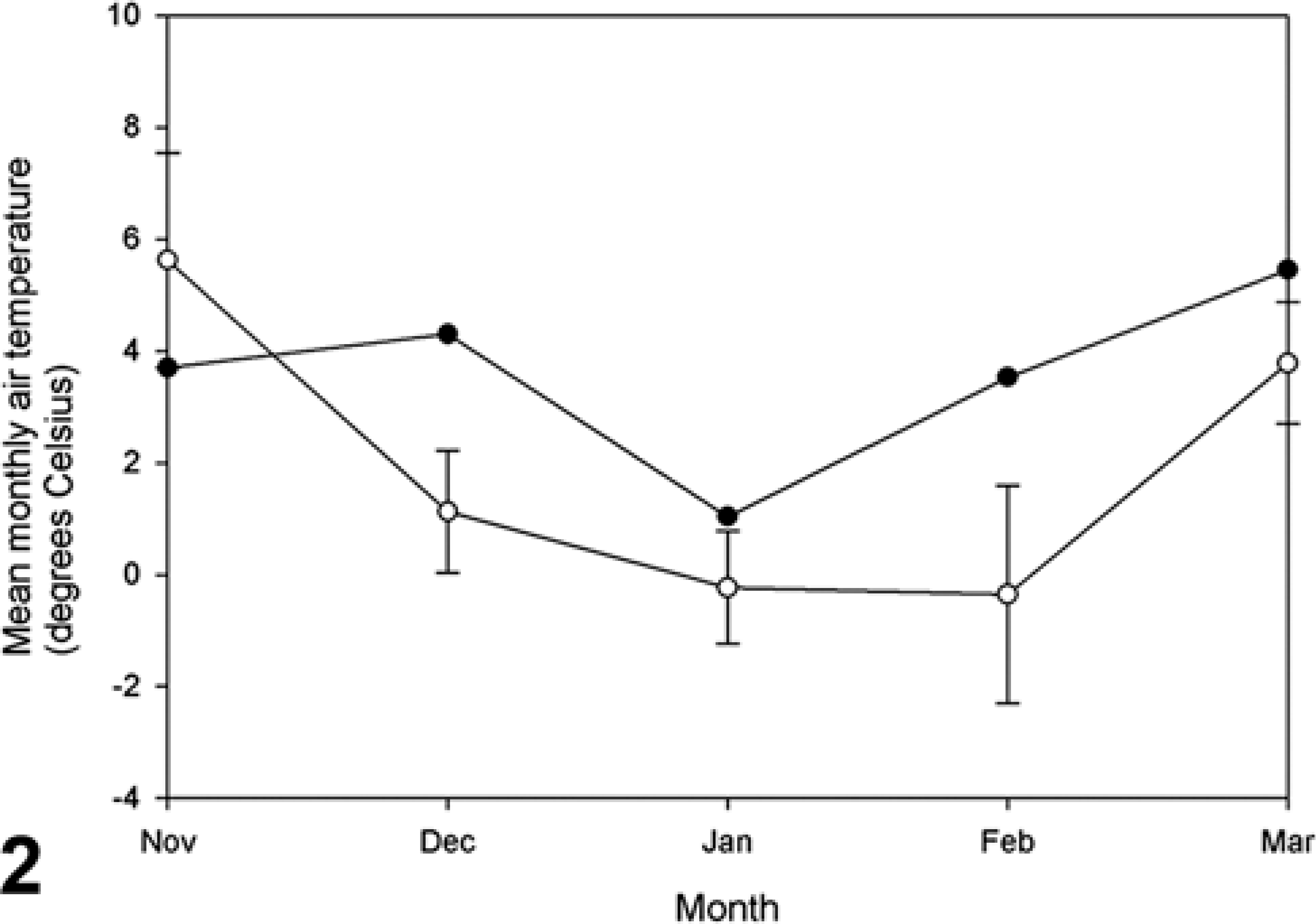
Mean monthly air temperature in the northern Caspian Sea during winter 1999/2000 (•) compared with the overall average (±95% confidence limit, ○) in the period 1990/1991 to 2000/2001.
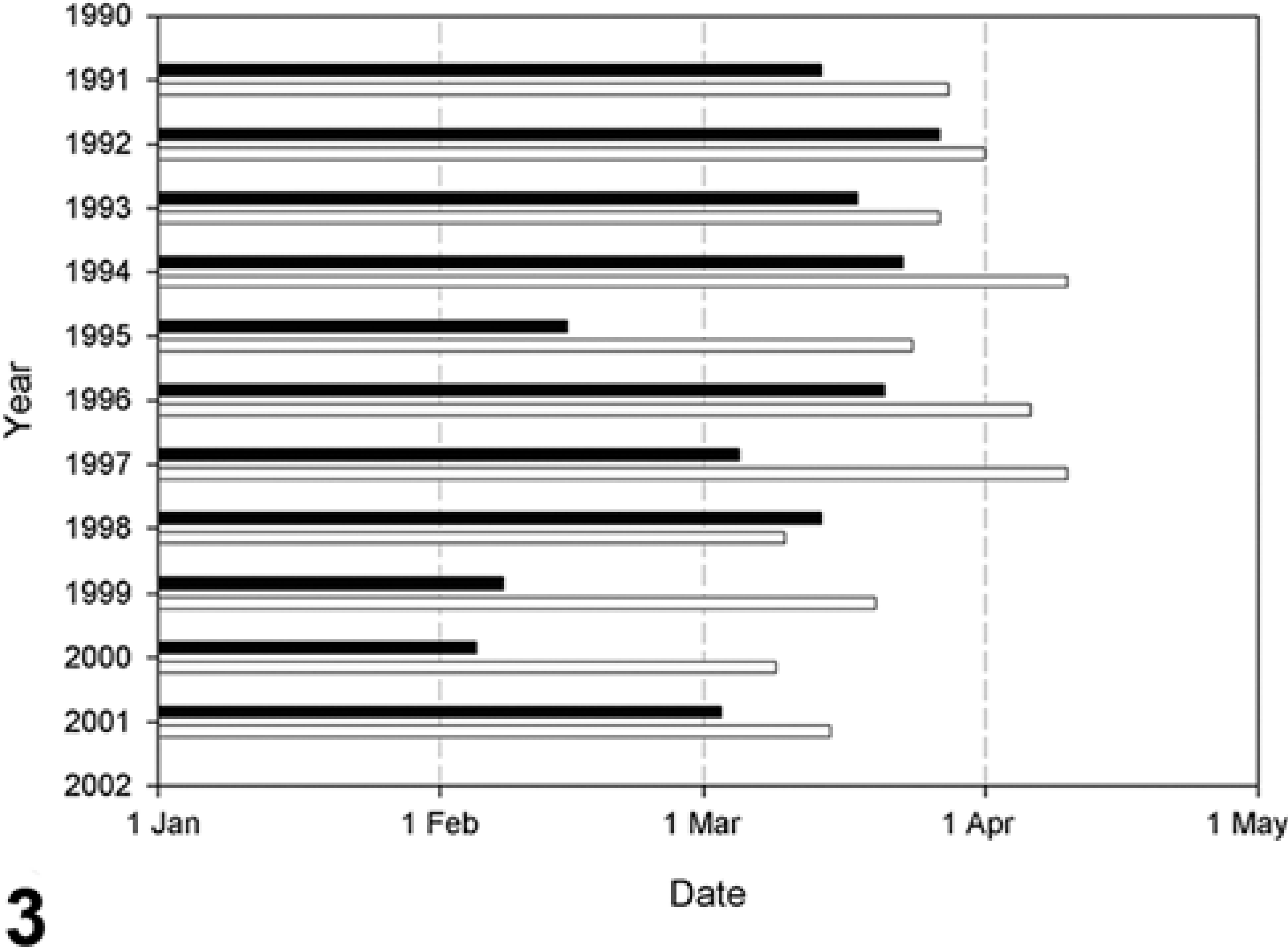
Date of disappearance of ice cover at Fort Shevchenko, Mangistau Region (▪), and at Peshnoi Island, Atyrau Region (□), northern Caspian Sea in the period 1990/1991 to 2000/2001. (Figure 1 shows these ← locations on the Caspian Sea coast.)
Strandings of Caspian seals in Azerbaijan
Weekly beach surveys on the northern shore of the Apsheron Peninsula from 1997 to 2000 showed two peaks of strandings per year, one in the spring and the other in the fall (Fig. 4). From January to May 2000, the stranding rate exceeded the historic mean plus SD for the corresponding months, except for February 2000 (Fig. 4).

Monthly stranding rate of Caspian seals at Azizbekov Beach, Apsheron Peninsula, Azerbaijan, in 2000 (▪) compared with the historical mean (±SD, □) for the period 1997 to 1999.
A total of 92 Caspian seal carcasses were found during single beach surveys at three different locations along the northern shore of the Apsheron Peninsula in June 2000. The density of carcasses was 15 ± 4.2 (mean ± SD; range, 11 to 20) per kilometer of beach. Of these carcasses, 2.2% were freshly dead to mildly decomposed, 78% were moderately to markedly decomposed, and 20% consisted only of skeletal remains with partly dried skin. Of those in which the sex was determined (n = 35), 74% were male and 26% were female. However, it is likely that the percentage of males was overestimated because it was easier to identify males than females in autolyzed carcasses. Based on those in which the standard length was determined (n = 70), 40% were 0 to 1 year of age and 60% were >1 year of age.
Clinical signs
Two seals, Nos. 1 and 18, were examined alive before death. They were weak, had opaque, colorless (No. 1) or green-white (No. 18) ocular and nasal exudate, and muscle tremors and spasms; the body of one seal (No. 18) was bent.
Gross lesions
In this study, a seal was considered to have canine distemper if morbilliviral antigen was detected by immunohistochemistry, or—in those seals where immunohistochemistry was not performed—characteristic inclusion bodies were seen histologically, and morbilliviral nucleic acid was detected by RT-PCR. Using this case definition, the diagnosis of canine distemper was made in 11 of 13 seals (Table 2). Advanced decomposition of 3 of 18 examined seals (Nos. 7, 8, and 16) precluded meaningful pathologic examination and collection of adequate samples for diagnosis of canine distemper. No samples for histology or immunohistochemistry were collected from two other seals (Nos. 1 and 2).
Summary of laboratory results for diagnosis of canine distemper virus infection in Caspian seals.∗
RT-PCR = reverse transcriptase–polymerase chain reaction; Ig = immunoglobulin.
Based mainly on the presence of intracytoplasmic inclusion bodies in epithelial tissues.
Seals with canine distemper were in poor to good nutritional condition and had no food in their stomach (5/5). The most frequent gross lesion was multifocal pulmonary consolidation throughout both lungs (4/7), which consisted of multiple, irregular, dark-red, firm areas, samples of which sank when placed in water. Other respiratory lesions included scant to abundant white to red-tinged foamy fluid in trachea and bronchi (7/7), subpleural emphysema in the caudodorsal region of both lungs (1/7), and diffuse interlobular edema (1/7). Lesions in other organs included generalized moderate thickening of the bile ducts (1/7), multiple to coalescing erosions in the oral mucosa (2/7), multifocal ulceration of the stomach (1/5), and suppurative balanoposthitis, consisting of a 2-mm-thick layer of pus covering the penis and prepuce (1/4).
The liver of one seal (No. 15) was pale tan, had an accentuated acinar pattern, and exuded yellow-tinged bloody fluid on section. The gall bladder (about 5 × 3 × 3 cm) was distended with viscous bile. The common bile duct was firm and thickened (1 cm in diameter) at the level of the duodenal wall but was still patent. The blubber, aorta, and kidneys of this seal also were tinged yellow. Seal No. 15 also had multiple epidermal plaques on the ventral surface of the left and right metatarsus suggestive of poxvirus infection. These plaques (7- to 10-mm diameter, 2-mm high) were alopecic, round, and raised, and on section the lesional epidermis was about twice the normal thickness.
Two of 13 seals (Nos. 10 and 12) were diagnosed negative for canine distemper (Table 2). The cause of death for these seals was not determined; although Salmonella dublin was isolated from multiple tissues of seal No. 10 (see Bacteriology section). Both seals were in poor nutritional condition and had no food in their stomach. A liver lobe of one seal (No. 10) was very firm, mottled gray and black, and contained numerous trematode parasites. Another liver lobe contained a poorly encapsulated abscess containing green-beige pus. The rest of the liver contained small, white fibrous cysts containing trematode parasites. The hepatic lymph node was enlarged, firm, and gray-brown. Otherwise, neither of these two seals had significant gross lesions.
Histologic lesions
The most severe lesion associated with CDV infection was a broncho-interstitial pneumonia (6/11). Multifocally throughout the lungs, there was degeneration and necrosis of bronchiolar epithelium, characterized by sloughing, intra- and intercellular vacuolization, and infiltration with occasional lymphocytes and neutrophils. Many bronchiolar epithelial cells contained round to oval, eosinophilic to amphophilic intracytoplasmic inclusion bodies (ICIB), ranging from about 2 to 10 µm in diameter and often surrounded by a clear space (halo) (Fig. 5). The bronchiolar lumina contained necrotic debris, sloughed epithelial cells, and macrophages, some also containing ICIB. Similar lesions were occasionally present in the epithelium of bronchi and trachea, and in the bronchial mucous glands. Multifocally with a peribronchiolar concentration, the alveolar walls were thickened by congestion and infiltration by mononuclear cells and occasional neutrophils. There was loss of alveolar epithelium and, rarely, type II pneumocyte hyperplasia; and the alveolar lumina contained erythrocytes, serous fluid, fibrin, cellular debris, macrophages, and occasional syncytia. ICIB were present in some of these macrophages and syncytia.
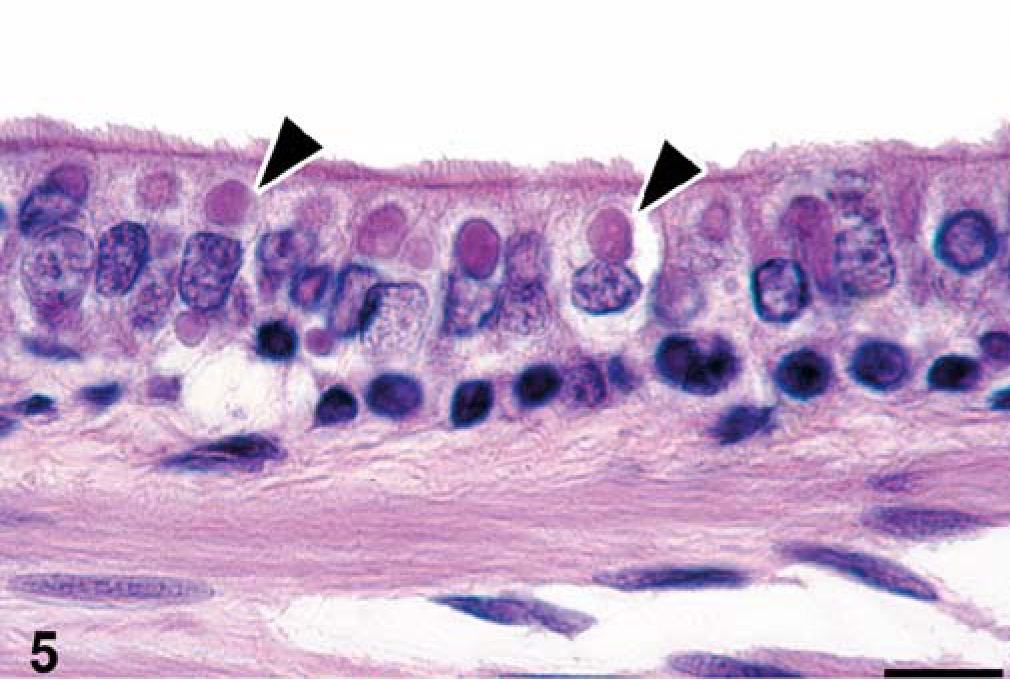
Bronchiole, seal No. 9. Single, large, eosinophilic intracytoplasmic inclusion bodies (arrowheads), often surrounded by an empty space (halo) are present in ciliated epithelial cells. HE stain. Bar = 12 µm.
In addition to the respiratory epithelium, ICIB were found in epithelial cells of the bile ducts (3/7) (Fig. 6), pancreatic collecting ducts (2/6), renal pelvis (5/8) (Fig. 7), penile urethra and outer mucosa (1/1), urinary bladder (3/5) (Fig. 8), preputium (1/1), acinar glands and collecting ducts of the prostate (1/1), esophagus (1/2), jejunum (1/2), colon (1/2), and eyelid conjunctiva (1/1), often associated with epithelial degeneration and necrosis and infiltration with scant to moderate numbers of neutrophils, lymphocytes, and plasma cells. In addition to ICIB, rare epithelial cells in the collecting ducts of the prostate also contained eosinophilic intranuclear inclusion bodies (INIB), 2 to 3 µm in diameter, surrounded by a halo. Intracytoplasmic inclusion bodies were found in neurons and ependymal cells of the cerebrum (2/7) and in the fibroblasts associated with chronic cholangitis (1/5).

Liver, seal No. 17. Multiple, variably sized, eosinophilic, intracytoplasmic inclusion bodies (arrowheads) are present in epithelial cells of the bile duct. HE stain. Bar = 12 µm.
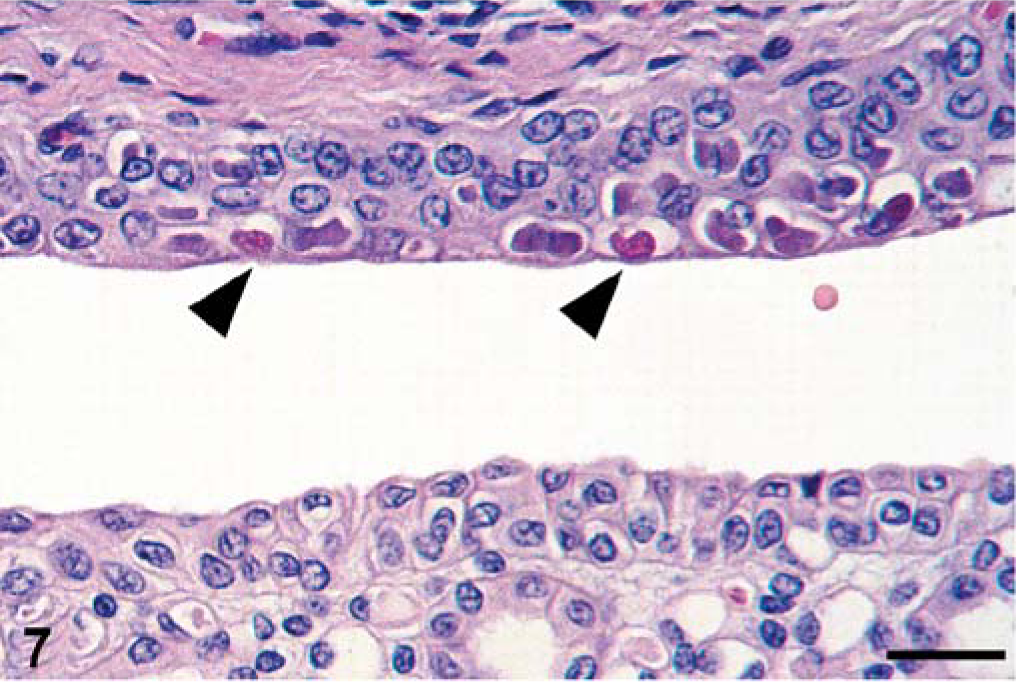
Kidney, seal No. 18. Single, large, eosinophilic intracytoplasmic inclusion bodies (arrowheads), often surrounded by a halo are present in transitional epithelial cells of the renal pelvis. HE stain. Bar = 30 µm.

Urinary bladder, seal No. 18. Single, large, eosinophilic intracytoplasmic inclusion bodies (arrowheads), often surrounded by a halo are present in sloughed transitional epithelial cells that are loose in the bladder lumen. HE stain. Bar = 12 µm.
The spleens of seals with canine distemper had marked lymphocytic depletion (8/9). The splenic capsule was corrugated, suggesting splenic atrophy, and the white pulp was partly to completely devoid of lymphocytes. The remaining dendritic cells and large mononuclear cells (histiocytes) often contained one to multiple ICIB, similar to those found in epithelial cells. In 3/11 seals, ICIB were present in histiocytes in the red pulp. The mesenteric (1/1) and inguinal (1/1) lymph nodes had similar changes, with marked lymphocytic depletion and ICIB in many histiocytes in cortex and medulla.
Lesions not associated with CDV infection included those of viral, bacterial, protozoal, metazoan, and noninfectious etiology. The only other viral lesion identified was poxvirus-associated dermatitis in seal No. 15. It consisted of multiple foci of epidermal hyperplasia, epidermal necrosis, dermatitis, and characteristic, irregular, eosinophilic to amphophilic ICIB, 2 to 10 µm in diameter, in keratinocytes (Fig. 9). By transmission electron microscopy, poxvirus particles with a typical internal dumbbell structure were detected in affected keratinocytes (Fig. 9, inset).
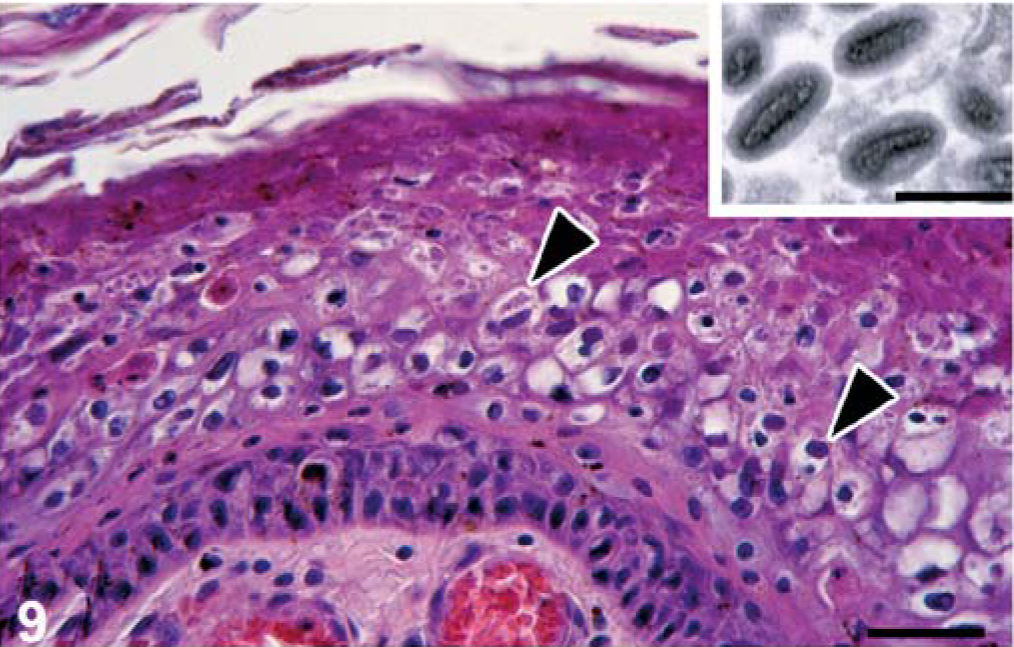
Skin, seal No. 15. The epidermis is hyperplastic, and irregular eosinophilic to amphophilic intracytoplasmic inclusion bodies (arrowheads), often surrounded by a halo, are present in swollen keratinocytes. HE stain. Bar = 30 µm. Inset: By transmission electron microscopy, poxvirus particles with typical internal dumbbell structures are present in a serial section of the same specimen. Uranyl acetate and lead citrate. Bar = 400 nm.
Bacteria associated with histologic lesions included Salmonella choleraesuis and Corynebacterium caspium. Salmonella choleraesuis infection in seal No. 15 was associated with septicemia, where clusters of Gram-negative bacilli (S. choleraesuis) were found associated with hemorrhage and serofibrinous exudation into pulmonary alveoli and microthrombi in the cerebellum and cerebrum (Fig. 10). These clusters of bacteria also were found without associated lesions in thymus, skeletal muscle, renal glomeruli, and lumen of the common bile duct. This seal also had chronic epithelial hyperplasia of the common bile duct, resulting in its partial obstruction. The bile duct mucosa had multiple highly convoluted papillary structures lined by simple columnar epithelium. Corynebacterium caspium infection was associated with suppurative balanoposthitis (Fig. 11) in seal No. 9. The mucosa of the penis and prepuce was largely necrotic and covered by a thick layer of cellular debris admixed with many colonies of Gram-positive coccobacilli (C. caspium). The underlying connective tissue was infiltrated by many neutrophils and a few macrophages and erythrocytes. In a remaining segment of penile epithelium, many epithelial cells contained ICIB characteristic for CDV infection.
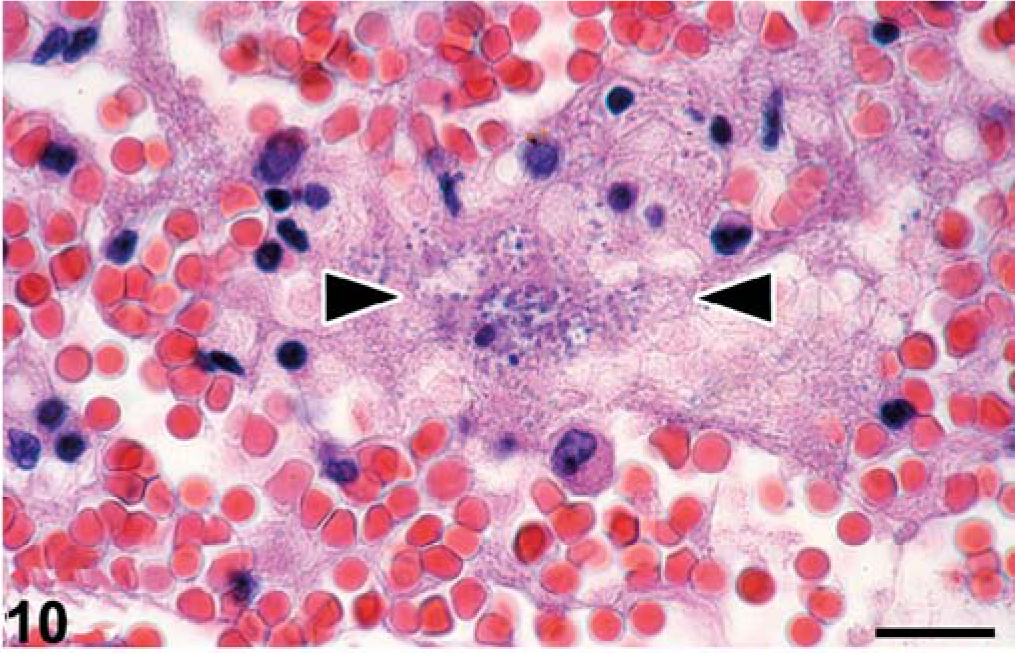
Lung, seal No. 22. A cluster of coccobacilli (Salmonella choleraesuis) (between arrowheads) are present in the lumen of a pulmonary alveole, mixed with serous fluid, fibrin, erythrocytes, and degenerate inflammatory cells. HE stain. Bar = 12 µm.
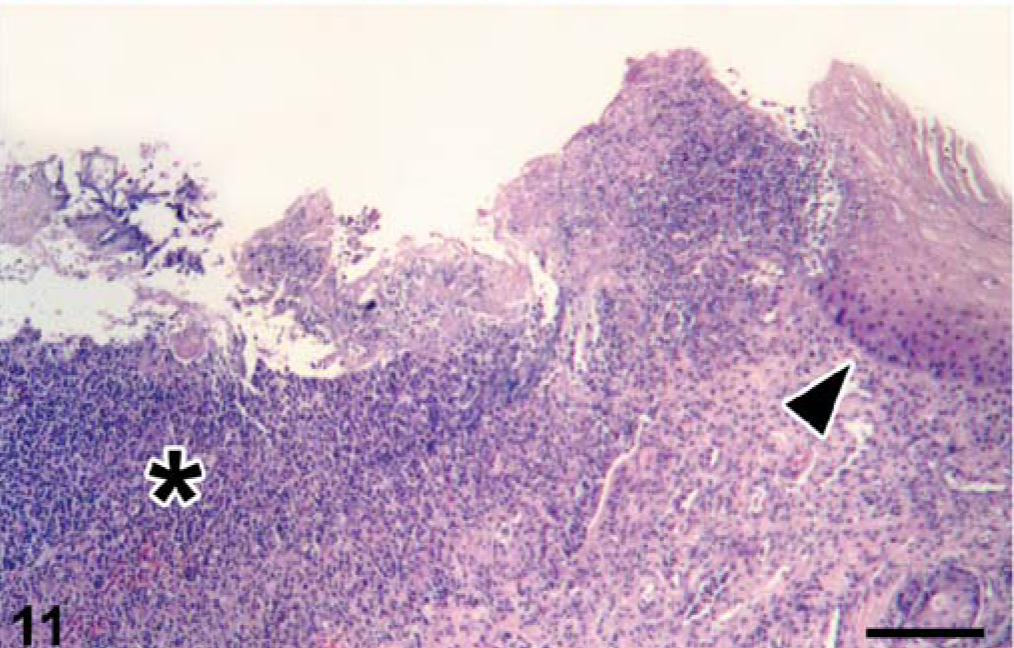
Penis, seal No. 9. Balanoposthitis. The penis is ulcerated and infiltrated with many neutrophils (asterisk). A remnant of epithelium (arrowhead) is present on one side. The surface of the penis is covered with debris mixed with aggregates of bacteria (Corynebacterium caspium). HE stain. Bar = 120 µm.
Protozoa detected histologically were Sarcocystis-like organisms and Eimeria-like organisms. Sarcocystis-like cysts containing many banana-shaped, light-purple bradyzoites (about 6 × 2 µm) with a dark-purple nucleus were detected in myocytes of the esophageal muscularis and skeletal muscle of seal No. 11. The cysts distended the infected myocyte but did not induce an inflammatory response in adjacent tissue (Fig. 12). Eimeria-like oocysts were detected in the cytoplasm of jejunal enterocytes of seal No. 9. The oocysts consisted of 4 to 8 banana-shaped zoites (2 × 8 µm) with a central blue nucleus and surrounded by a narrow, birefringent wall. Some of the infected cells were sloughed.
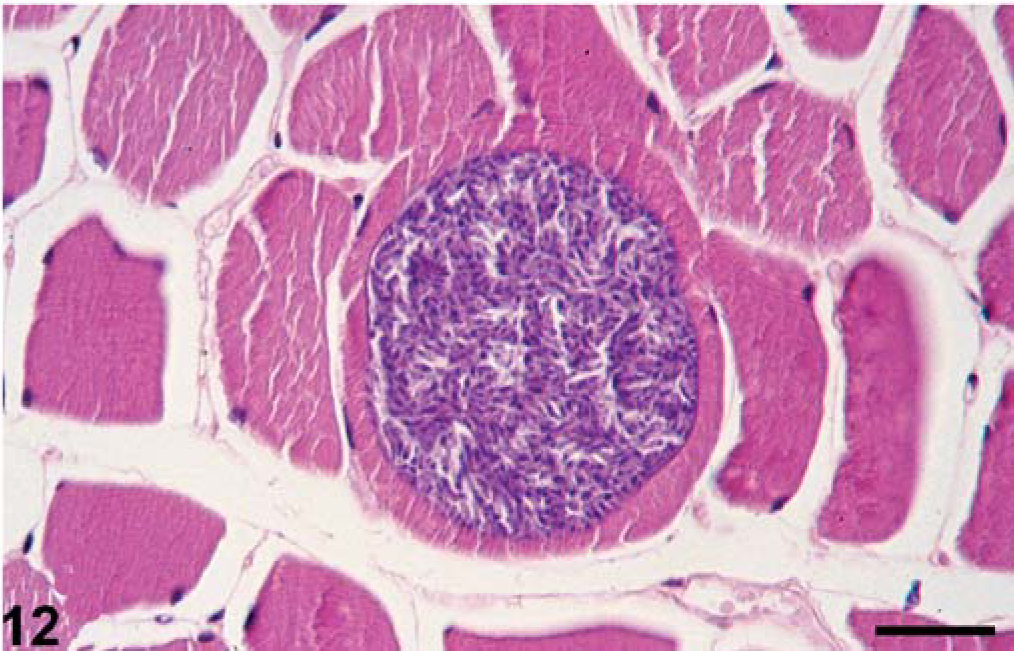
Skeletal muscle, seal No. 11. A myocyte is distended by a round Sarcocystis-like cyst, filled with many bradyzoites. HE stain. Bar = 30 µm.
Metazoa associated with lesions included the trematode Pseudamphistomum truncatum, a lung nematode, probably Filaroides caspicus, and the acanthocephalan Corynosoma strumosum. The most pathogenic was Pseudamphistomum truncatum infection, detected histologically in seal Nos. 3, 4, 5, 9, and 10. The most severely affected seal was No. 10. In this seal, the presence of trematodes in bile duct lumina was associated with bile duct hyperplasia and fibrosis, severe hepatic necrosis, and abscess formation. Trematodes also were present in the lumen of large pancreatic ducts in association with pancreatic duct hyperplasia, and in the sinusoids of the hepatic lymph node in association with sinus histiocytosis, microabscess formation, and fibroplasia. In the other seals, the presence of P. truncatum was associated with hepatic granulomas (Nos. 3, 4, and 5), bile duct hyperplasia (No. 9), pancreatic granulomas, and pancreatic duct hyperplasia (Nos. 4 and 5). Infection of the lungs with a nematode, probably Filaroides caspicus, was found in seal Nos. 6, 11, 13, and 17. These were characterized by multiple cross-sections of nematode parasites of about 10 to 50 µm diameter (the larger ones containing larvae) in the lumen of bronchi and alveoli. There was no or minimal associated inflammatory response, except in seal No. 11, where eosinophilic granulomas had formed around parasite remnants in the alveolar tissue. Corynosoma strumosum infection of the jejunum and colon was found in seal Nos. 9, 11, and 15. These parasites were characterized by a spike-covered head and were embedded multifocally in the mucosa up to the level of the submucosa. There was minimal infiltration of neutrophils and macrophages in the adjacent tissue.
A lesion of noninfectious etiology was arteriosclerosis in the uterus of seal No. 11. This lesion was characterized by distension of the media by homogeneous pink (fibrinoid) and granular blue (calcified) material, intimal thickening, and endothelial hyperplasia, resulting in narrowing or obliteration of the arterial lumina.
Immunohistochemistry for morbillivirus
Morbillivirus antigen expression was detected in 9 of 11 seals tested (Table 3) in the epithelial cells of the trachea (Fig. 13), bile duct (Fig. 14), renal pelvis (Fig. 15), gastric mucosa (Fig. 16), fallopian tube, renal tubule, pulmonary alveole, bronchus, esophagus, esophageal submucosal gland, pancreatic duct, gastric mucosa, and urinary bladder. Besides in epithelial cells, morbillivirus expression also was detected in ependymal cells and neurons in the brain; alveolar macrophages in the lung; lymphocytes, macrophages, dendritic cells, and Schwann cells in lymph nodes; submucosal fibroblasts and submucosal glandular epithelial cells in the esophagus; fibroblasts in hepatic and pancreatic P. truncatum-associated granulomas; lymphocytes and macrophages in the spleen; and mononuclear cells, fibroblasts, smooth muscle cells, and myenteric plexus cells in the stomach wall.
Distribution of morbillivirus antigen in Caspian seals diagnosed with canine distemper.
Number of cells expressing morbillivirus antigen, based on immunohistochemistry: - = none; + = small numbers; ++ = moderate numbers; +++ = large numbers. No morbillivirus antigen expression was seen in the following tissues from these seals: adrenal gland (3 seals tested), heart (4), intestine (2), ovary (3), skin (3), testis (1), thyroid gland (1), and uterus (3).
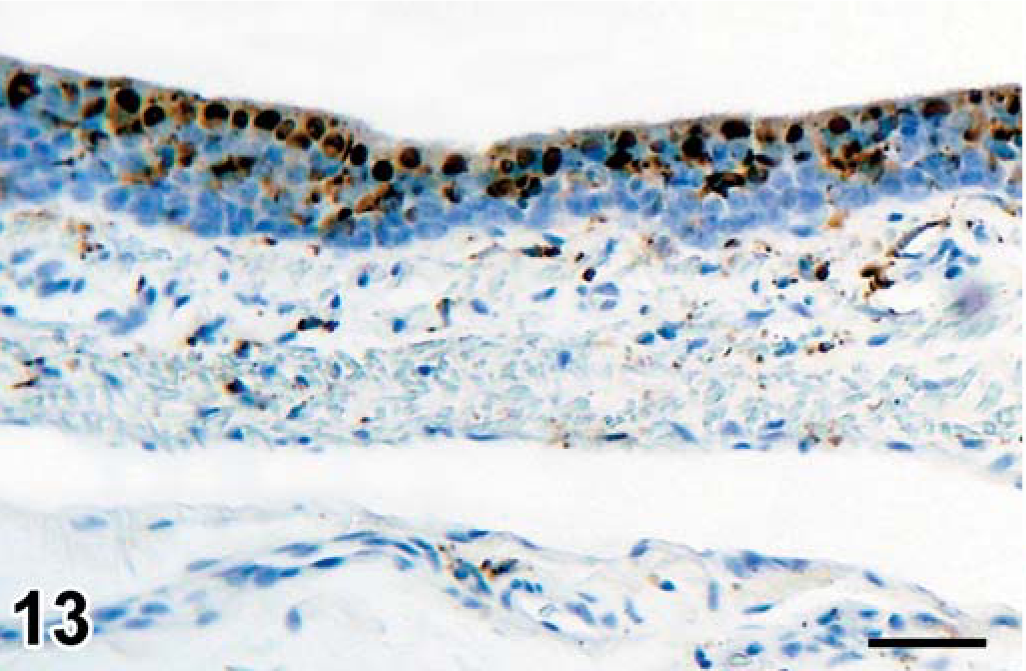
Trachea, seal No. 9. Morbillivirus antigen in cytoplasm of ciliated epithelial cells. Immunoperoxidase histochemistry and hematoxylin. Bar = 60 µm.
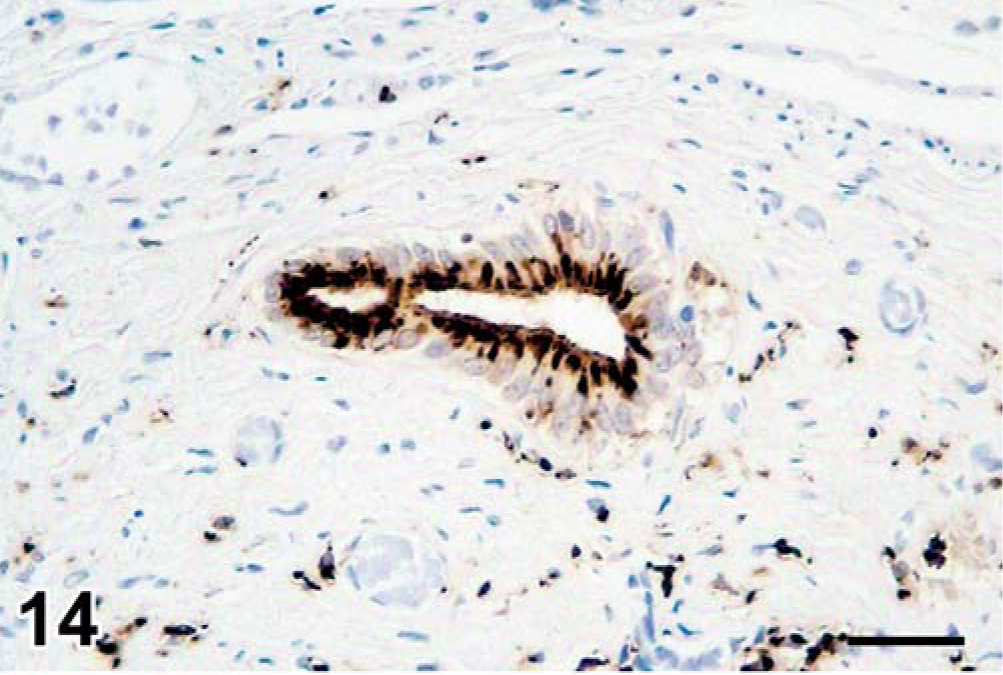
Liver, seal No. 9. Morbillivirus antigen in cytoplasm of bile duct epithelial cells. Immunoperoxidase histochemistry and hematoxylin. Bar = 60 µm.
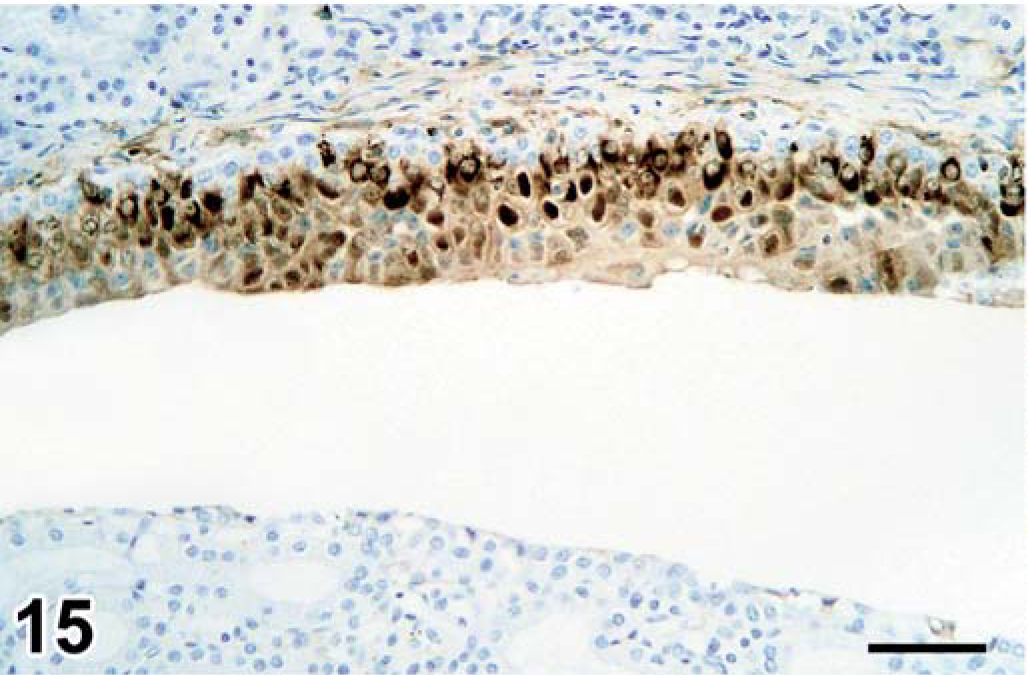
Kidney, seal No. 9. Morbillivirus antigen in cytoplasm of renal pelvis epithelial cells. Immunoperoxidase histochemistry and hematoxylin. Bar = 60 µm.
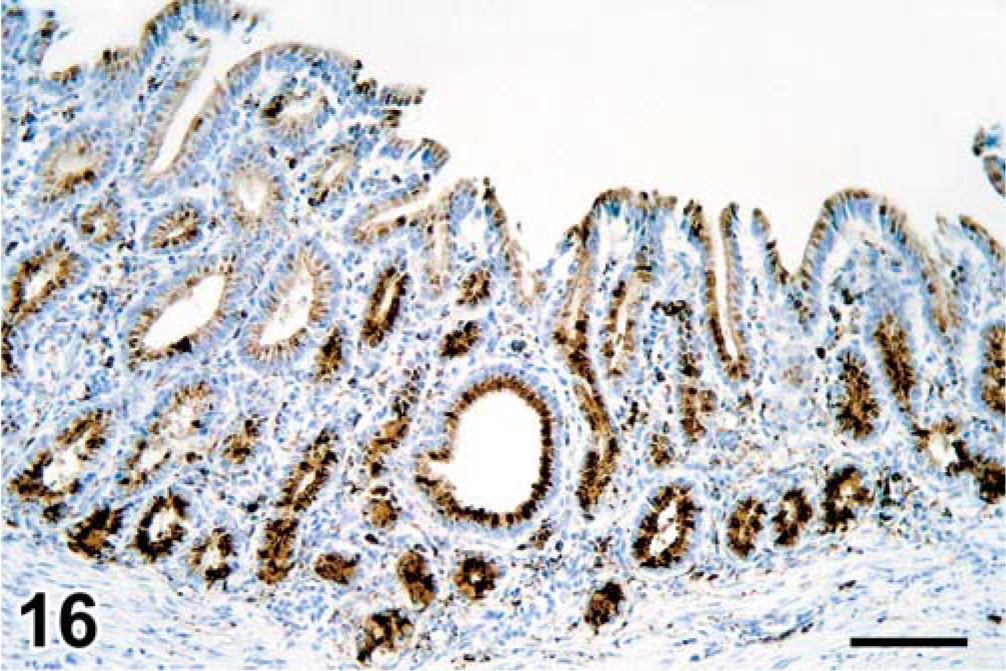
Stomach, seal No. 9. Morbillivirus antigen in cytoplasm of gastric epithelial cells. Immunoperoxidase histochemistry and hematoxylin. Bar = 120 µm.
No morbillivirus antigen expression was found in the following tissues of two seals (Nos. 10 and 12): adrenal gland (2 seals), brain (2), heart (1), intestine (2), kidney (2), liver (2), lung (2), lymph node (2), pancreas (2), skin (1), spleen (2), stomach (2), testis (1), thymus (1), thyroid gland (1), trachea (1), urinary bladder (1), and uterus (1).
RT-PCR for influenza A and B virus and morbillivirus
Nine of 14 seals tested positive for morbillivirus by RT-PCR (Table 2). All samples tested negative in the RT-PCR for influenza A and influenza B virus.
Serology
Eight of 13 seals had CDV-specific IgM antibody, and 12 of 13 had CDV-specific IgG antibody (Table 2). No antibodies to Brucella were found in the sera tested.
Bacteriology
Bordetella bronchiseptica was isolated from lung samples of four seals (Nos. 3 to 5, 14) (Table 4). In addition, it was also isolated from samples of trachea/bronchi and liver of one of these seals (No. 3). By ribotyping, the four isolates belonged to RT8. A hitherto unrecognized Corynebacterium sp. nov. was isolated from samples of lung and penis of one seal (No. 9). It was named Corynebacterium caspium, and its detailed description is presented elsewhere. 10 Salmonella choleraesuis was isolated from liver, kidney, spleen, and mesenteric lymph node of one seal (No. 15). Salmonella dublin was isolated from samples of lung, spleen, liver, pancreas, kidney, heart, and blood of one seal (No. 4), and from samples of lung, liver, kidney, spleen, cerebrum, mesenteric lymph node, and jejunum of another seal (No. 10). Streptococcus phocae was isolated from samples of lung (3/4), liver (2/4), spleen (1/4), cerebrum (1/3), mesenteric lymph node (1/2), and heart (1/1) of four seals (Nos. 4, 10, 14, and 15). Streptococcus sp., group G, was isolated from samples of liver (3/4), kidney (3/4), spleen (3/4), lung (2/4), cerebrum (2/3), mesenteric lymph node (2/2), heart (1/1), left thoracic cavity (1/1), right uterine horn (1/1), and penis (1/1) of four seals (Nos. 3, 9, 12, and 14).
Bacteria isolated from Caspian seals.
Atopobacter phocae, Corynebacterium caspium, Vibrio anguillarum
Bacillus sp., Plesiomonas shigelloides
Acinetobacter johnsonii, Actinobacillus sp., Pseudomonas fluorescens, Salmonella choleraesuis, Vibrio harveyii
Parasitology
Numerous nasal mites, identified as Halarachne sp., were found in the nasal cavity (1/13). Nematode parasites, identified as Anisakis schupakovi, were found free in the stomach lumen (2/9). One seal had about 10, and the other about 50 of these stomach worms. Acanthocephalan parasites, identified as Corynosoma strumosum, were found lightly attached to the intestinal mucosa (7/7). They were generally more frequent in the second half of the small intestine than in the first half of the small intestine and colon and had a density of up to 50 parasites per 5-cm length of intestine. Not observed macroscopically but detected incidentally during laboratory investigation of ethanol-fixed intestinal samples, were trematode parasites (Cryptocotyle lingua, 1/3; and Mesorchis advena, 2/3). The presence of the above parasites was not associated with grossly visible lesions. Trematode parasites, identified as Pseudamphistomum truncatum, were found in the liver of one of three seals with gross lesions of the biliary tree and/or liver.
Algal toxins
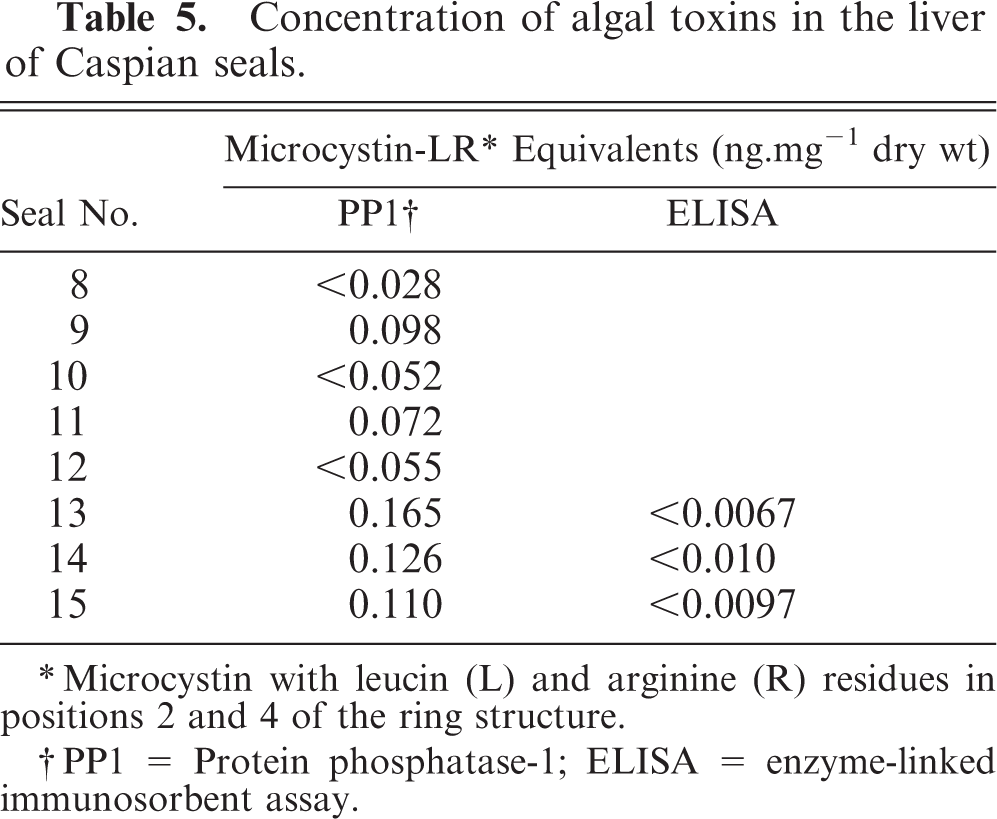
The liver samples from five seals tested weakly positive for microcystin (i.e., less than 0.2 ng mg−1 dry weight) in the protein phosphatase 1 inhibition assay (Table 5). Three of these also were tested in an ELISA specific for microcystins and nodularin and were negative. These results indicate that neither of these toxins was present in the liver samples at detectable concentrations (i.e., less than 0.03 ng mg−1 dry weight).
Concentration of algal toxins in the liver of Caspian seals.
Microcystin with leucin (L) and arginine (R) residues in positions 2 and 4 of the ring structure.
PP1 = Protein phosphatase-1; ELISA = enzyme-linked immunosorbent assay.
Toxicology
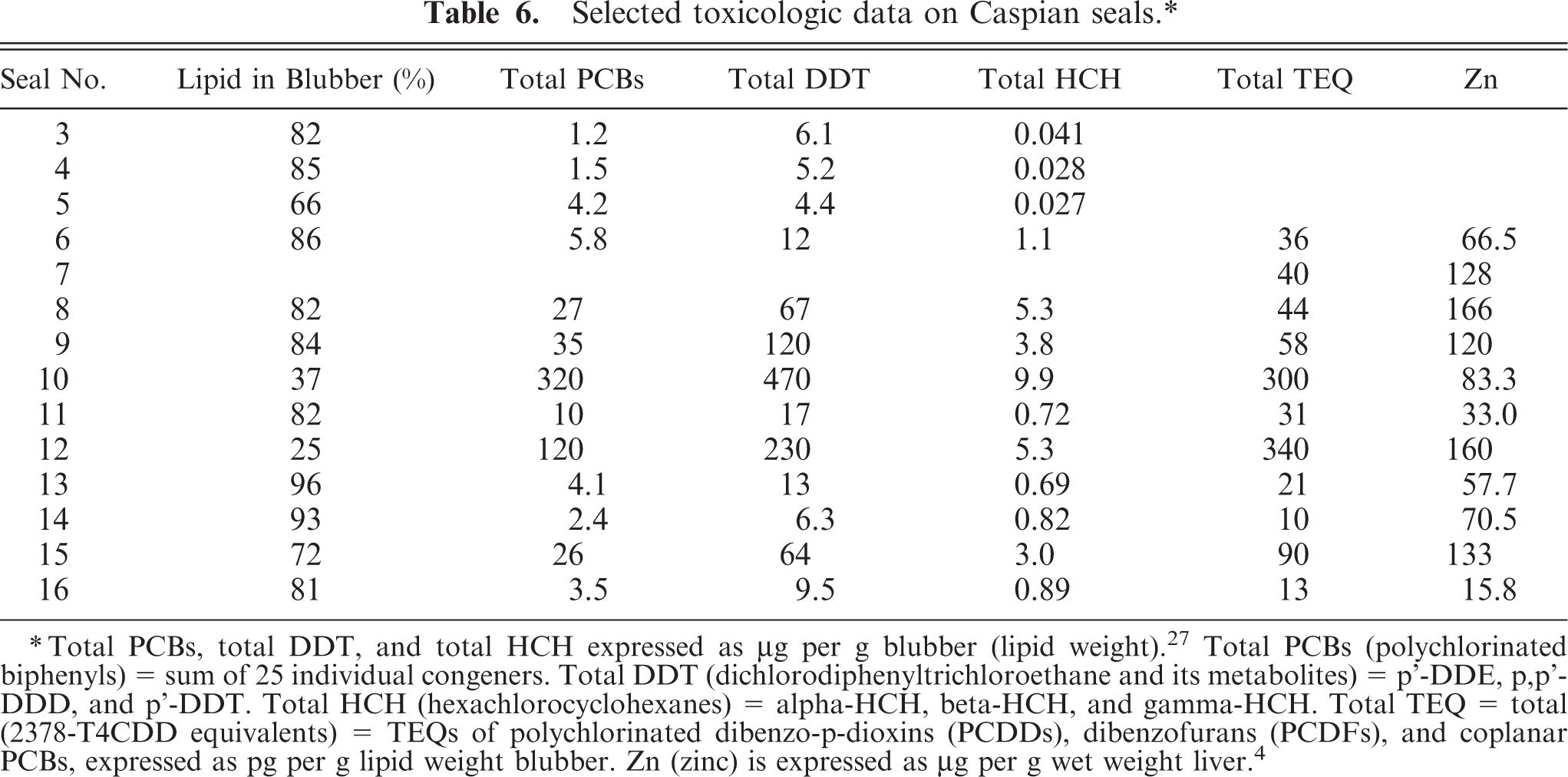
The concentrations of organochlorines in the blubber of seals with distemper were approximately 5 to 10 times lower than in seals without distemper and were comparable to the concentrations found in seals in previous years (Tables 6 and 7). Similarly, the concentrations of Zn in the liver of seals with distemper were approximately 1.3 times lower than in seals without distemper (Table 7). The concentrations were comparable to those found in Caspian seals in previous years (Table 7). The lower organochlorine and zinc concentrations were associated with the lower proportion of adults (33% vs. 80%) and the greater blubber thickness (16 mm vs. 8 mm) in the canine distemper–positive group compared with the canine distemper–negative group (Table 7).
Selected toxicologic data on Caspian seals.∗
Total PCBs, total DDT, and total HCH expressed as μg per g blubber (lipid weight).27 Total PCBs (polychlorinated biphenyls) = sum of 25 individual congeners. Total DDT (dichlorodiphenyltrichloroethane and its metabolites) = p'-DDE, p,p'-DDD, and p'-DDT. Total HCH (hexachlorocyclohexanes) = alpha-HCH, beta-HCH, and gamma-HCH. Total TEQ = total (2378-T4CDD equivalents) = TEQs of polychlorinated dibenzo-p-dioxins (PCDDs), dibenzofurans (PCDFs), and coplanar PCBs, expressed as pg per g lipid weight blubber. Zn (zinc) is expressed as μg per g wet weight liver.4
Organochlorine concentrations (mean and range, μg per g blubber [lipid weight]) and zinc concentrations (mean and range, μg per g liver [wet weight]) in Caspian seals from 2000 with and without canine distemper, and in those from previous years.
Seals of 4 years of age or younger were considered juveniles (J); seals greater than 4 years of age were considered adults (A).
Total PCBs (polychlorinated biphenyls) = sum of 25 congeners.
Total DDT (dichlorodiphenyltrichloroethane and its metabolites) = p′-DDE, p,p′-DDD, and p′-DDT.
The age category of one seal was not determined.
Not determined for three seals.
C. Allchin and R. Law (unpublished data).
The sex of three seals was not determined.
Discussion
The high mortality of Caspian seals in the spring and summer of 2000 exceeded 10,000 animals and extended from the breeding area in the northern Caspian Sea, via the spring migration route, to the feeding grounds in the central Caspian Sea (Fig. 1). The number of strandings in 2000 was increased compared with that in previous years, with a monthly stranding rate of up to 2.8 times the historic mean (Fig. 4). Canine distemper was the primary diagnosis in 11/13 Caspian seal carcasses obtained from the north to the southwest of the Caspian Sea during this period (Table 1), based on the combined results of histopathology, immunohistochemistry, and RT-PCR (Table 2). Phylogenetic analysis of samples from Azerbaijan and Kazakhstan showed that seals from widely separated regions of the Caspian Sea were infected by the same virus. 29
The clinical signs in Caspian seals with canine distemper were lethargy, serous or mucopurulent discharge from eyes and nose, muscle tremors, and abnormal posture. The most consistent gross lesion was multifocal pulmonary consolidation, and the major histologic lesions were broncho-interstitial pneumonia, lymphocytic necrosis and depletion in lymphoid organs, and the presence of eosinophilic intracytoplasmic and, occasionally, intranuclear inclusion bodies in epithelia of multiple tissues. These findings are similar to those in harbor seals with phocine distemper. 28 The presence of morbillivirus antigen in the penile and preputial epithelium in one Caspian seal also was found in a harbor porpoise (Phocoena phocoena) with morbillivirus infection, 30 which suggests the possibility of venereal transmission of CDV. Morbillivirus antigen in the prostate, found in one Caspian seal, to our knowledge has not been recorded previously in marine mammals with morbillivirus infection. The occurrence of intracytoplasmic inclusion bodies in fibroblasts of one seal (No. 9) with chronic cholangitis was surprising because they are normally not found in this cell type. However, intracytoplasmic inclusion bodies in fibroblasts have been detected in the lung of a harbor porpoise with morbillivirus and concurrent lungworm infection 30 and in a mesenchymal epididymal tumor of a common dolphin with dolphin morbillivirus infection. 8 The relatively thick blubber layer compared with animals that died from undetermined cause (Table 7), the absence of fibroplasia in alveolar walls, and the rare occurrence of type 2 pneumocyte hyperplasia in the Caspian seals with canine distemper indicates that these animals died acutely from this disease.
The reservoir for CDV in the Caspian Sea area is not known but could be either the Caspian seal population itself, or sympatric terrestrial carnivore populations. It is not known what size Caspian seal population is minimally required to maintain CDV. A human population size of at least 300,000 is considered necessary to maintain measles virus, another morbillivirus. 9 Based on a longitudinal serological survey, Duignan and co-authors hypothesized that a phocine distemper virus–like morbillivirus infection was endemic in the eastern Canadian gray seal population, and presumed that the population size of more than 120,000 was large enough to maintain infection. 14 Therefore, based on the most recently published population estimate of 360,000 to 400,000 for Caspian seals, 32 it is conceivable that the Caspian seal population itself could act as a reservoir for CDV. An alternative is that CDV is introduced into Caspian seals by adjacent populations of terrestrial carnivores. Domestic or feral dogs and wolves have been suggested as possible sources of CDV infection for Caspian seals. 16 However, until now there has been no confirmed diagnosis of canine distemper or isolation of CDV in these two species or other terrestrial carnivores from the Caspian Sea area.
Bordetella bronchiseptica was isolated preferentially from lung tissue of Caspian seals with canine distemper (Table 4), probably because canine distemper causes immunosuppression and facilitates infection with opportunistic pathogens. 15 Bordetella bronchiseptica, which is part of the normal flora of the upper respiratory tract in dogs, 15 is also a common secondary pathogen in the lungs of both harbor seals with phocine distemper 6, 22, 40 and dogs with canine distemper. 4 The ribotype (RT8) of the Caspian seal isolates has only been found previously in two horses (K. Register, personal communication). All previously ribotyped B. bronchiseptica strains recovered from harbor seals (34 from Scotland and one from Denmark) belonged to ribotype RT19. 45 Similar to B. bronchiseptica, Streptococcus phocae was recovered preferentially from Caspian seals with canine distemper (Table 4). This organism also was a common secondary pathogen in harbor seals with phocine distemper. 49
Salmonella spp. have been found commonly in different species of pinnipeds 23 and have been associated occasionally with disease, including meningoencephalomyelitis. 50 One Caspian seal (No. 15) in our study had Salmonella choleraesuis septicemia, characterized by hemorrhage and serofibrinous exudation into pulmonary alveoli and microthrombi in brain vessels in association with Gram-negative bacilli. This serovar also causes septicemia in domestic pigs. 7 The concurrent severe bile duct lesions and resulting obstructive jaundice in this individual may have predisposed it to salmonellosis, as has been found in humans and rats. The proposed mechanisms for this phenomenon are that decreased concentration of bile acids in the intestinal lumen enhance epithelial internalization of enteric bacteria, 13, 54 while the increased serum bile acid level reduces uptake of endotoxins by Kupffer's cells in the liver. 51 Two other Caspian seals (Nos. 4 and 10) had generalized Salmonella dublin infection, but associated lesions were not identified by gross necropsy or histologic examination. This serovar primarily affects cattle. 7
Two other bacterial isolates that are particularly noteworthy are Corynebacterium caspium, which is the first and only known isolation of this organism and has been described in more detail elsewhere, 10 and Atopobacter phocae. The latter organism was described following its isolation from two harbor seals in Scotland 38 and has since been recovered from a further three harbor seals, seven gray seals, and a European otter (Lutra lutra) in the United Kingdom (G. Foster, unpublished). However, this is the first report of A. phocae from Caspian seals and also the first from outside the United Kingdom. Several other species of bacteria were isolated from these Caspian seals, including known facultative pathogens such as Aeromonas sp., Escherichia coli, Plesiomonas shigelloides, and Vibrio spp. (Table 4). Although these isolates were not obviously associated with septicemia or histologic lesions, it cannot be ruled out that they played a role in the mortality of these seals.
The trematodes Pseudoamphistomum truncatum, Cryptocotyle lingua, and Mesorchis advena, the nematode Anisakis schupakovi, and the acanthocephalan Corynosoma strumosum have been described previously in Caspian seals. 44 They were not considered significant pathogens in the seals of this study, except for P. truncatum, which was associated with hepatic and pancreatic lesions of variable severity. Infection with P. truncatum was considered by Krylov to cause mortality in Caspian seals. 32, 33 No published records were found of the nasal mite Halarachne sp. in Caspian seals, although infection with mites of this genus have been found in many other phocid seals. 44 Although parasites were not found in the lungs at gross necropsy, cross-sections of nematode parasites were found in the lung of four seals (Nos. 6, 11, 13, and 17) by histologic examination. The species concerned is likely Filaroides caspicus, because this is the only nematode known to occur in the lung of Caspian seals. The coccidial oocysts observed in the intestine of seal No. 9 were not identified more specifically. They were similar to the oocysts of Eimeria phocae, which occurs in harbor seals. 24 However, no published records of protozoal infections in Caspian seals were found. The protozoal cysts found in the striated muscle of seal no. 11 were not identified further; such cysts have been found in harbor seals with a Sarcocystis neurona–like protozoal infection. 35
Based on reinterpretation of the toxicological data according to the CDV status of the Caspian seals, we found that the concentrations of organochlorines (particularly PCBs, DDTs, and HCHs) and Zn in tissues of seals with distemper in 2000 were comparable to the concentrations found in seals in previous years (Table 7). Furthermore, the contaminant burdens in these seals were seven times lower than the level (209 ng TEQ/kg lipid) at which the immune function of harbor seals was impaired in a semi–field experiment. 47 Finally, the Zn concentrations were within the range of 20–100 µg g−1 wet weight generally reported in marine mammals. 37 Based on these comparisons, albeit in a small number of animals, there is no evidence that these contaminants contributed to the mortality from canine distemper in Caspian seals in 2000. This conclusion differs from those of preliminary toxicological analyses, 3, 27 in which organochlorine and Zn concentrations in tissues of Caspian seals found stranded in 2000 were considered to be higher than in previous years. Based on these findings, the authors suggested that these contaminants may have made the seals more susceptible to CDV infection owing to immunosuppression and recommended further investigation. The reason for this discrepancy is that seal Nos. 10 and 12, which had the highest contaminant concentrations (Table 6), were included in the calculations of the earlier studies, while they were excluded from our study because they were diagnosed negative for canine distemper (Table 2). The unusually high organochlorine concentrations in these two seals is likely owing to their high age and thin blubber layer, both of which are positively correlated with organochlorine and Zn concentrations in seal tissues. 1, 37
The current trend toward a warming climate has been associated with a worldwide increase in reports of diseases affecting marine organisms. 21 In five mass mortality events of pinnipeds with available data, mean monthly temperatures preceding the mortalities were higher than the 10-year average. 36 We observed the same phenomenon in our study, where an unusually warm winter (Fig. 2) preceded the canine distemper outbreak of Caspian seals in 2000. The mechanism by which this warm winter might have increased mortality from canine distemper is unknown. Perhaps the unusually early disappearance of ice cover in the northern Caspian Sea in spring 2000 (5 February to 9 March; Fig. 3) decreased the available area for hauling out, thus concentrating seals at fewer haul-out sites, with a resultant increase in the spread of CDV. The early loss of ice cover also may have shortened the suckling period, which normally lasts until the second half of February, 33 resulting in reduced body weight of juveniles at weaning and decreased resistance to disease. These hypotheses would need to be tested by a combination of behavioral and meteorologic studies.
In conclusion, we identified canine distemper as the primary diagnosis in 11/13 Caspian seal carcasses obtained during a mortality event in the spring and summer of 2000, when the monthly stranding rate of seals was up to 2.8 times greater than the historic mean. The main lesions consisted of broncho-interstitial pneumonia, lymphocytic necrosis and depletion in lymphoid organs, and the presence of prominent intracytoplasmic inclusion bodies in multiple epithelia. Concurrent bacterial infections that may have contributed to the illness of the affected seals include Bordetella bronchiseptica, Streptococcus phocae, Salmonella dublin, and S. choleraesuis. A new bacterium, Corynebacterium caspium, was identified from the inflamed penis and prepuce of one Caspian seal, while several infectious and parasitic organisms, including poxvirus, Atopobacter phocae, Eimeria- and Sarcocystis-like organisms, and Halarachne sp. were identified in Caspian seals for the first time. Reinterpretation of the toxicological data according to the CDV status of the Caspian seals provided no evidence that anthropogenic contaminants contributed to the mortality from canine distemper in Caspian seals in 2000. However, the unusually mild winter preceding the mortality event in 2000 may have done so, through increased ambient air temperature and accelerated disappearance of ice cover at the breeding areas in the northern Caspian Sea.